
Antifungal Activity and DNA Topoisomerase Inhibition of Hydrolysable Tannins from Punica granatum L.


 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Antifungal Activity of Pomegranate Polyphenols against Fungal Phytopathogens
2.2. Antifungal Activity of Pomegranate Polyphenols against Human Pathogens
2.2.1. Agar Disk Diffusion Assay
2.2.2. Minimum Inhibitory Concentration Assessment
2.2.3. Time Kill Curves
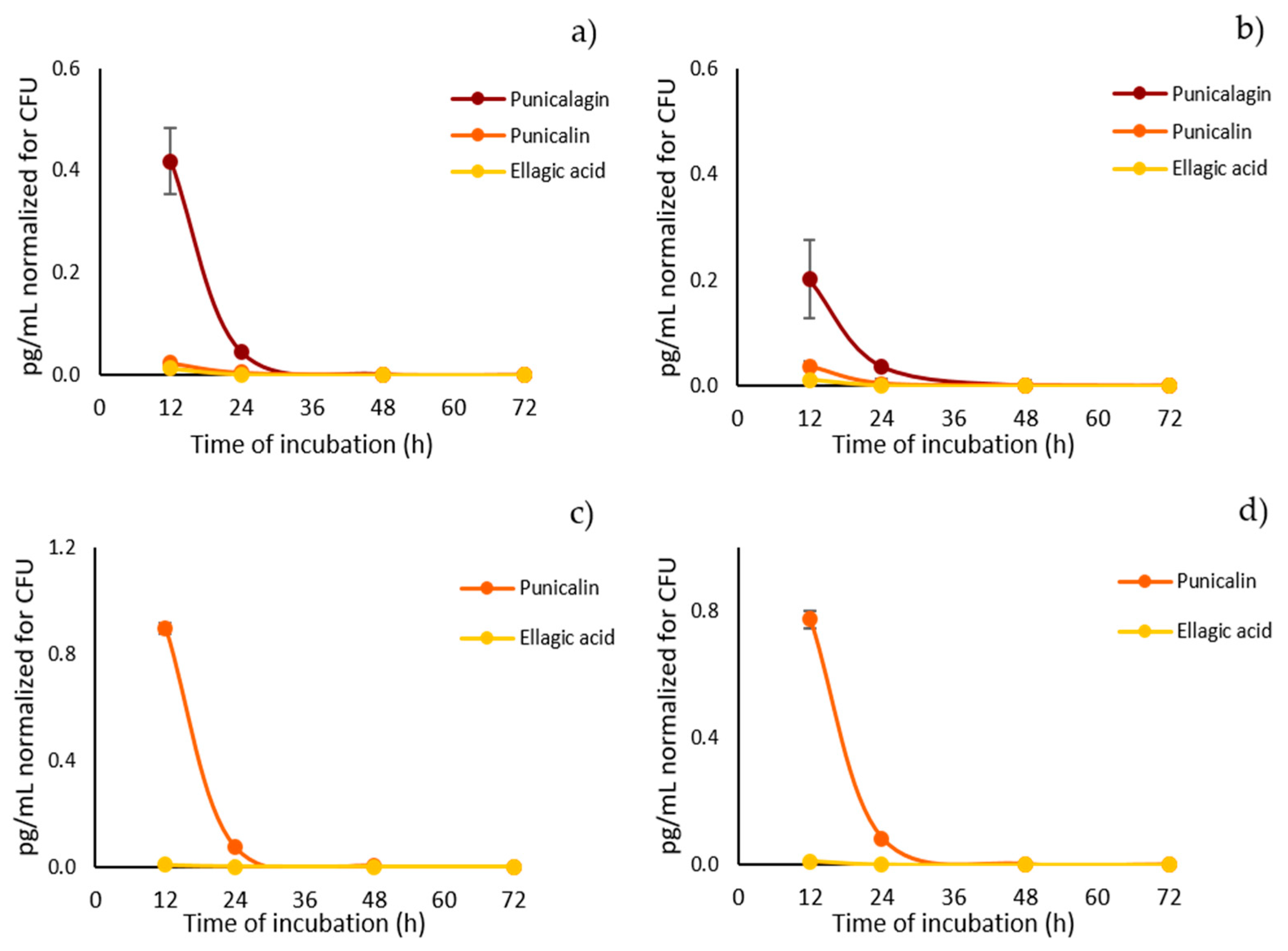
2.3. Quantitative Analysis of Pomegranate Polyphenols in Lysates from Candida albicans Cultures
2.4. Identification of Topoisomerase I and II as Potential Puncicalagin Targets from Chemoinformatic Analysis
2.5. In Vitro Interactions of Pomegranate Polyphenols with Candida Topoisomerases
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Pomegranate Fungal Pathogens Strains
3.3. Human Fungal Pathogens
3.4. Determination of the Antifungal Activity against Phytopathogens
3.5. Determination of Antifungal Activity against Human Pathogens
3.5.1. Agar Disk Diffusion Assay
3.5.2. MIC Determination
3.5.3. Time-Kill Curves
3.6. Incubation of Candida albicans Strains with Pomegranate Compounds and Cell Lysis
3.7. Quantitative HPLC-HRMS Analysis of Pomegranate Compounds in Lysates from Candida albicans Cultures
3.8. In Silico Ligand-Based Analysis
3.9. Evaluation of the In Vitro Interactions with Candida Topoisomerases
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rastelli, G.; Pellati, F.; Pinzi, L.; Gamberini, M.C. Repositioning Natural Products in Drug Discovery. Molecules 2020, 25, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldholmi, M.; Marchand, P.; Ourliac-Garnier, I.; Le Pape, P.; Ganesan, A. A Decade of Antifungal Leads from Natural Products: 2010–2019. Pharmaceuticals 2019, 12, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anibal, P.C.; Peixoto, I.T.A.; Foglio, M.A.; Höfling, J.F. Antifungal Activity of the Ethanolic Extracts of Punica granatum L. and Evaluation of the Morphological and Structural Modifications of Its Compounds upon the Cells of Candida spp. Braz. J. Microbiol. 2013, 44, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rongai, D.; Pulcini, P.; Di Lernia, G.; Nota, P.; Preka, P.; Milano, F. Punicalagin Content and Antifungal Activity of Different Pomegranate (Punica granatum L.) Genotypes. Horticulturae 2019, 5, 52. [Google Scholar] [CrossRef] [Green Version]
- Mincuzzi, A.; Ippolito, A.; Brighenti, V.; Marchetti, L.; Benvenuti, S.; Ligorio, A.; Pellati, F.; Sanzani, S.M. The Effect of Polyphenols on Pomegranate Fruit Susceptibility to Pilidiella granati Provides Insights into Disease Tolerance Mechanisms. Molecules 2020, 25, 515. [Google Scholar] [CrossRef] [Green Version]
- Ascacio-Valdés, J.A.; Aguilera-Carbó, A.F.; Buenrostro, J.J.; Prado-Barragán, A.; Rodríguez-Herrera, R.; Aguilar, C.N. The complete biodegradation pathway of ellagitannins by Aspergillus niger in solid-state fermentation. J. Basic Microbiol. 2016, 56, 329–336. [Google Scholar] [CrossRef] [PubMed]
- E Ribas, A.D.R.; Spolti, P.; Del Ponte, E.M.; Donato, K.Z.; Schrekker, H.; Fuentefria, A.M. Is the emergence of fungal resistance to medical triazoles related to their use in the agroecosystems? A mini review. Braz. J. Microbiol. 2016, 47, 793–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekaert, W.F.; Terras, F.R.; Cammue, B.P.; Vanderleyden, J. An automated quantitative assay for fungal growth inhibition. FEMS Microbiol. Lett. 1990, 69, 55–59. [Google Scholar] [CrossRef]
- Raudabaugh, D.B.; Iturriaga, T.; Carver, A.; Mondo, S.; Pangilinan, J.; Lipzen, A.; He, G.; Amirebrahimi, M.; Grigoriev, I.V.; Miller, A.N. Coniella lustricola, a new species from submerged detritus. Mycol. Prog. 2018, 17, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Lachhab, N.; Sanzani, S.M.; Bahouaoui, M.A.; Boselli, M.; Ippolito, A. Effect of some protein hydrolysates against gray mould of table and wine grapes. Eur. J. Plant. Pathol. 2016, 144, 821–830. [Google Scholar] [CrossRef]
- Fraternale, D.; Ricci, D.; Verardo, G.; Gorassini, A.; Stocchi, V.; Sestili, P. Activity of Vitis vinifera tendrils extract against phytopathogenic fungi. Nat. Prod. Commun. 2015, 10, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Viswanath, V.; Urooj, A.; Malleshi, N.G. Evaluation of antioxidant and antimicrobial properties of finger millet polyphenols (Eleusine coracana). Food Chem. 2009, 114, 340–346. [Google Scholar] [CrossRef]
- Bisignano, C.; Filocamo, A.; Faulks, R.M.; Mandalari, G. In vitro antimicrobial activity of pistachio (Pistacia vera L.) polyphenols. FEMS Microbiol. Lett. 2013, 341, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, T. Antimicrobial Activities of Tea Polyphenol on Phytopathogens: A Review. Molecules 2019, 24, 816. [Google Scholar] [CrossRef] [Green Version]
- Sanzani, S.M.; Schena, L.; De Girolamo, A.; Ippolito, A.; González-Candelas, L. Characterization of Genes Associated with Induced Resistance against Penicillium expansum in Apple Fruit Treated with Quercetin. Postharvest Biol. Technol. 2010, 56, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Munhuweyi, K.; Lennox, C.L.; Meitz-Hopkins, J.C.; Caleb, O.J.; Opara, U.L. Major Diseases of Pomegranate (Punica granatum L.), their Causes and Management—A Review. Sci. Hortic. 2016, 211, 126–139. [Google Scholar] [CrossRef]
- Endo, E.H.; Cortez, D.A.G.; Ueda-Nakamura, T.; Nakamura, C.V.; Dias Filho, B.P. Potent Antifungal Activity of Extracts and Pure Compound isolated from Pomegranate Peels and Synergism with Fluconazole against Candida albicans. Res. Microbiol. 2010, 161, 534–540. [Google Scholar] [CrossRef]
- Gaulton, A.; Hersey, A.; Nowotka, M.; Bento, A.P.; Chambers, J.; Mendez, D.; Mutowo, P.; Atkinson, F.; Bellis, L.J.; Cibrián-Uhalte, E.; et al. The ChEMBL Database in 2017. Nucleic Acids Res. 2017, 45, D945–D954. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Llorach, R.; Cerón, J.J.; Espín, J.C.; Tomás-Barberán, F.A. Evaluation of the bioavailability and metabolism in the rat of punicalagin, an antioxidant polyphenol from pomegranate juice. Eur. J. Nutr. 2003, 42, 18–28. [Google Scholar] [CrossRef]
- Hawkins, P.C.D.; Skillman, A.G.; Nicholls, A. Comparison of Shape-Matching and Docking as Virtual Screening Tools. J. Med. Chem. 2007, 50, 74–82. [Google Scholar] [CrossRef] [PubMed]
- ROCS 3.3.2.2; OpenEye Scientific Software: Santa Fe, NM, USA, 2019; Available online: http://www.eyesopen.com (accessed on 4 December 2019).
- Rahman, S.; Rahman, T. Unveiling Some FDA-Approved Drugs as Inhibitors of the Store-Operated Ca2+ Entry Pathway. Sci. Rep. 2017, 7, 12881. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, S.R.; Moore, J.B.; Schymura, Y.; Churchill, G.C. Shape-Based Reprofiling of FDA-Approved Drugs for the H1 Histamine Receptor. J. Med. Chem. 2012, 55, 7054–7060. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, L.; Benedetti, R.; Altucci, L.; Rastelli, G. Design of Dual Inhibitors of Histone Deacetylase 6 and Heat Shock Protein 90. ACS Omega 2020, 5, 11473–11480. [Google Scholar] [CrossRef]
- Pinzi, L.; Lherbet, C.; Baltas, M.; Pellati, F.; Rastelli, G. In Silico Repositioning of Cannabigerol as a Novel Inhibitor of the Enoyl Acyl Carrier Protein (ACP) Reductase (InhA). Molecules 2019, 24, 2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, S.M.; Berry, D.E.; MacKenzie, L.J.; Busby, R.W.; Nasuti, C.A. A Strategy for Identifying Novel, Mechanistically Unique Inhibitors of Topoisomerase I. J. Nat. Prod. 1992, 55, 401–413. [Google Scholar] [CrossRef]
- Lothstein, L.; Suttle, D.P.; Roaten, J.B.; Koseki, Y.; Israel, M.; Sweatman, T.W. Catalytic Inhibition of DNA Topoisomerase II by N-Benzyladriamycin (AD 288). Biochem. Pharmacol. 2000, 60, 1621–1628. [Google Scholar] [CrossRef]
- Bodley, A.; Liu, L.F.; Israel, M.; Seshadri, R.; Koseki, Y.; Giuliani, F.C.; Kirschenbaum, S.; Silber, R.; Potmesil, M. DNA Topoisomerase II-Mediated Interaction of Doxorubicin and Daunorubicin Congeners with DNA. Cancer Res. 1989, 49, 5969–5978. [Google Scholar] [PubMed]
- Tang, Z.-B.; Chen, Y.-Z.; Zhao, J.; Guan, X.-W.; Bo, Y.-X.; Chen, S.-W.; Hui, L. Conjugates of Podophyllotoxin and Norcantharidin as Dual Inhibitors of Topoisomerase II and Protein Phosphatase 2A. Eur. J. Med. Chem. 2016, 123, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.K.; Majumder, H.K.; Roychoudhury, S. Natural Compounds as Anticancer Agents Targeting DNA Topoisomerases. Curr. Genom. 2017, 18, 75–92. [Google Scholar] [CrossRef]
- Shen, L.L.; Baranowski, J.; Fostel, J.; Montgomery, D.A.; Lartey, P.A. DNA Topoisomerases from Pathogenic Fungi: Targets for the Discovery of Antifungal Drugs. Antimicrob. Agents Chemother. 1992, 36, 2778–2784. [Google Scholar] [CrossRef] [Green Version]
- Steverding, D.; Evans, P.; Msika, L.; Riley, B.; Wallington, J.; Schelenz, S. In Vitro Antifungal Activity of DNA Topoisomerase Inhibitors. Med. Mycol. 2012, 50, 333–336. [Google Scholar] [CrossRef] [Green Version]
- Abe, I.; Kashiwagi, Y.; Noguchi, H.; Tanaka, T.; Ikeshiro, Y.; Kashiwada, Y. Ellagitannins and Hexahydroxydiphenoyl Esters as Inhibitors of Vertebrate Squalene Epoxidase. J. Nat. Prod. 2001, 64, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.B.; Holloway, G.A. New Substructure Filters for Removal of Pan Assay Interference Compounds (PAINS) from Screening Libraries and for Their Exclusion in Bioassays. J. Med. Chem. 2010, 53, 2719–2740. [Google Scholar] [CrossRef] [Green Version]
- OMEGA 3.1.2.2; OpenEye Scientific Software: Santa Fe, NM, USA; Available online: http://www.eyesopen.com (accessed on 4 December 2019).
- Baell, J.B. Feeling Nature’s PAINS: Natural Products, Natural Product Drugs, and Pan Assay Interference Compounds (PAINS). J. Nat. Prod. 2016, 79, 616–628. [Google Scholar] [CrossRef]
- Bisson, J.; McAlpine, J.B.; Friesen, J.B.; Chen, S.-N.; Graham, J.; Pauli, G.F. Can Invalid Bioactives Undermine Natural Product-Based Drug Discovery? J. Med. Chem. 2016, 59, 1671–1690. [Google Scholar] [CrossRef] [PubMed]
- Bala, I.; Bhardwaj, V.; Hariharan, S.; Kumar, M.R. Analytical Methods for Assay of Ellagic Acid and its Solubility Studies. J. Pharm. Biomed. Anal. 2006, 40, 206–210. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Method for Antifungal Disk Diffusion Susceptibility Testing of Yeasts; Approved Guideline 2nd; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2009. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved standard—3rd ed. CLSI document M27-A3; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Lõoke, M.; Kristjuhan, K.; Kristjuhan, A. Extraction of Genomic DNA from Yeasts for PCR-based Applications. Biotechniques 2011, 50, 325–328. [Google Scholar] [CrossRef]
- Brighenti, V.; Groothuis, S.F.; Prencipe, F.P.; Amir, R.; Benvenuti, S.; Pellati, F. Metabolite Fingerprinting of Punica granatum L. (Pomegranate) Polyphenols by Means of High-Performance Liquid Chromatography with Diode Array and Electrospray Ionization-Mass Spectrometry Detection. J. Chromatogr. A 2017, 1480, 20–31. [Google Scholar] [CrossRef] [PubMed]
- ChEMBL Database. Available online: www.ebi.ac.uk/chembl (accessed on 5 March 2020).
- Schrödinger Release 2020-1: LigPrep; Schrödinger, LLC: New York, NY, USA, 2020.
- Hawkins, P.C.D.; Skillman, A.G.; Warren, G.L.; Ellingson, B.A.; Stahl, M.T. Conformer Generation with OMEGA: Algorithm and Validation Using High Quality Structures from the Protein Databank and Cambridge Structural Database. J. Chem. Inf. Model. 2010, 50, 572–584. [Google Scholar] [CrossRef]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Giambasu, G.; et al. AMBER 2018; University of California: San Francisco, CA, USA, 2018. [Google Scholar]
- Kraszni, M.; Marosi, A.; Larive, C.K. NMR Assignments and the Acid–Base Characterization of the Pomegranate Ellagitannin Punicalagin in the Acidic PH-Range. Anal. Bioanal. Chem. 2013, 405, 5807–5816. [Google Scholar] [CrossRef]
- Queimada, A.J.; Mota, F.L.; Pinho, S.P.; Macedo, E.A. Solubilities of Biologically Active Phenolic Compounds: Measurements and Modeling. J. Phys. Chem. B 2009, 113, 3469–3476. [Google Scholar] [CrossRef] [PubMed]
- Simić, A.Z.; Verbić, T.Ž.; Sentić, M.N.; Vojić, M.P.; Juranić, I.O.; Manojlović, D.D. Study of Ellagic Acid Electro-Oxidation Mechanism. Monatsh. Chem. 2013, 144, 121–128. [Google Scholar] [CrossRef]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef]
- Jiang, W.; Gerhold, D.; Kmiec, E.B.; Hauser, M.; Becker, J.M.; Koltin, Y. The Topoisomerase I Gene from Candida albicans. Microbiology 1997, 143, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Punicalagin a | Punicalin b | Ellagic Acid c | Gallic Acid d | Imazalil e |
|---|---|---|---|---|---|
| Alternaria alternata AL19 | 92.2 | 255.6 | 165.4 | 587.8 | 0.3 |
| Botrytis cinerea B2 | 92.2 | 255.6 | 165.4 | 587.8 | 0.3 |
| Colletotrichum acutatum s.s. M146-2 | 184.4 | 255.6 | 165.4 | 587.8 | 0.3 |
| Coniella granati M0_C2 | 184.4 | 255.6 | 165.4 | 587.8 | 0.3 |
| Strain | Punicalagin c | Punicalin d | Ellagic Acid e | Gallic Acid f | Fluconazole g | Amphotericin B h |
|---|---|---|---|---|---|---|
| Candida albicans ATCC 10321 a | 0.7 | 127.0 | 2.5 | 293.9 | 40.8 | 0.1 |
| Candida albicans SC 5314 a | 0.7 | 127.0 | 2.5 | 293.9 | 9.8 | 0.1 |
| Candida albicans 1 a,b | 1.4 | 3.8 | 5.0 | 293.9 | 81.6 | 2.2 |
| Candida albicans 2 a,b | 1.4 | 3.8 | 2.5 | 293.9 | 163.2 | 4.3 |
| Candida albicans H a,b | 92.2 | 1.9 | 331 | 587.8 | 81.6 | 4.3 |
| Candida albicans 40 a | 5.5 | 127.0 | 331 | 587.8 | 163.2 | 0.3 |
| Candida albicans 41 a | 92.2 | 127.0 | 331 | 293.9 | 81.6 | 0.3 |
| Candida albicans 44 a,b | 92.2 | 127.0 | 2.5 | 293.9 | 81.6 | 2.2 |
| Candida parapsilosis | 0.7 | 127.0 | 662 | 293.9 | 9.8 | 0.1 |
| Candida parapsilosis 7 a | 5.5 | 3.8 | 331 | 587.8 | 19.6 | 0.3 |
| Candida zeylanoides 33 a | 92.2 | 127.0 | 331 | 587.8 | 19.6 | 0.1 |
| Saccharomyces cerevisiae 42 b | 1.4 | 3.8 | 662 | 587.8 | 4.9 | 1.1 |
| Aspergillus brasiliensis ATCC 16404 | 2.8 | 1.9 | 662 | 8.8 | 4.9 | 0.1 |
| Aspergillus candidus 3 | 92.2 | 127.0 | 662 | 17.6 | 9.8 | 0.3 |
| Aspergillus candidus 25 | 92.2 | 127.0 | 662 | 17.6 | 9.8 | 0.3 |
| Cryptococcus neoformans B 3501 | 46.1 | 1.9 | 331 | 4.4 | 4.9 | 0.1 |
| Cryptococcus neoformans ATCC 11240 | 23.0 | 1.9 | 331 | 4.4 | 4.9 | 0.1 |
| Cryptococcus 67 | 92.2 | 127.0 | 662 | 8.8 | 9.8 | 0.1 |
| Cryptococcus var. grubii H99 serotype A | 92.2 | 127.0 | 662 | 4.4 | 4.9 | 0.1 |
| Reference | Compound Name | ChEMBL ID | Rank # |
|---|---|---|---|
| Punicalagin | Chebulagic acid | CHEMBL525240 | 8 |
| Punicalagin | Corilagin | CHEMBL449392 | 13 |
| Punicalagin | N, N-dibenzyl doxorubicin | CHEMBL3248005 | 14 |
| Punicalagin | Disaccharide derivative of daunorubicin | CHEMBL2367695 | 19 |
| Punicalagin | 4β-5-FU-substituted 4′-demethylepipodophyllotoxin | CHEMBL362359 | 41 |
| Punicalagin | 4-Acetic acid ester derivative of podophyllotoxin | CHEMBL4092572 | 48 |
| Punicalagin | N-benzyl doxorubicin | CHEMBL3303036 | 65 |
| Punicalagin | 4-Acetic acid ester derivative of podophyllotoxin | CHEMBL4060624 | 92 |
| Punicalagin | 4β-Norcantharidin-substituted -4′-demethylepipodophyllotoxin | CHEMBL3974286 | 99 |
| Punicalagin | Hydrazone derivative of N-morpholin-doxorubicin | CHEMBL66563 | 100 |
| Compounds | IC50 Value (µM) | |
|---|---|---|
| Topoisomerase I | Topoisomerase II | |
| Punicalagin | 9.0 ± 1.2 | 4.6 ± 0.4 |
| Punicalin | 14.7 ± 0.9 | 40.7 ± 0.8 |
| Ellagic acid | 56.6 ± 1.2 | 53.9 ± 0.2 |
| Camptothecin | 17.8 ± 0.7 | - |
| Etoposide | - | 15.9 ± 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brighenti, V.; Iseppi, R.; Pinzi, L.; Mincuzzi, A.; Ippolito, A.; Messi, P.; Sanzani, S.M.; Rastelli, G.; Pellati, F. Antifungal Activity and DNA Topoisomerase Inhibition of Hydrolysable Tannins from Punica granatum L. Int. J. Mol. Sci. 2021, 22, 4175. https://doi.org/10.3390/ijms22084175
Brighenti V, Iseppi R, Pinzi L, Mincuzzi A, Ippolito A, Messi P, Sanzani SM, Rastelli G, Pellati F. Antifungal Activity and DNA Topoisomerase Inhibition of Hydrolysable Tannins from Punica granatum L. International Journal of Molecular Sciences. 2021; 22(8):4175. https://doi.org/10.3390/ijms22084175
Chicago/Turabian StyleBrighenti, Virginia, Ramona Iseppi, Luca Pinzi, Annamaria Mincuzzi, Antonio Ippolito, Patrizia Messi, Simona Marianna Sanzani, Giulio Rastelli, and Federica Pellati. 2021. "Antifungal Activity and DNA Topoisomerase Inhibition of Hydrolysable Tannins from Punica granatum L." International Journal of Molecular Sciences 22, no. 8: 4175. https://doi.org/10.3390/ijms22084175
APA StyleBrighenti, V., Iseppi, R., Pinzi, L., Mincuzzi, A., Ippolito, A., Messi, P., Sanzani, S. M., Rastelli, G., & Pellati, F. (2021). Antifungal Activity and DNA Topoisomerase Inhibition of Hydrolysable Tannins from Punica granatum L. International Journal of Molecular Sciences, 22(8), 4175. https://doi.org/10.3390/ijms22084175