
Caffeine-Cyclodextrin Complexes as Solids: Synthesis, Biological and Physicochemical Characterization †


 , ,
, ,  , ,
, ,  and
and
Abstract
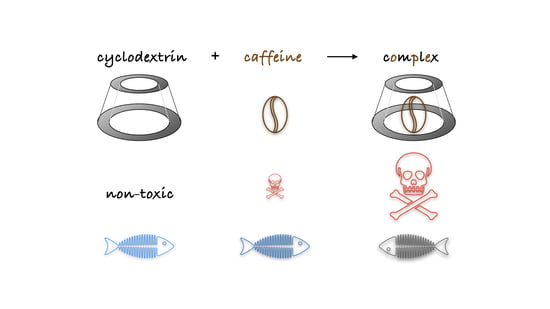
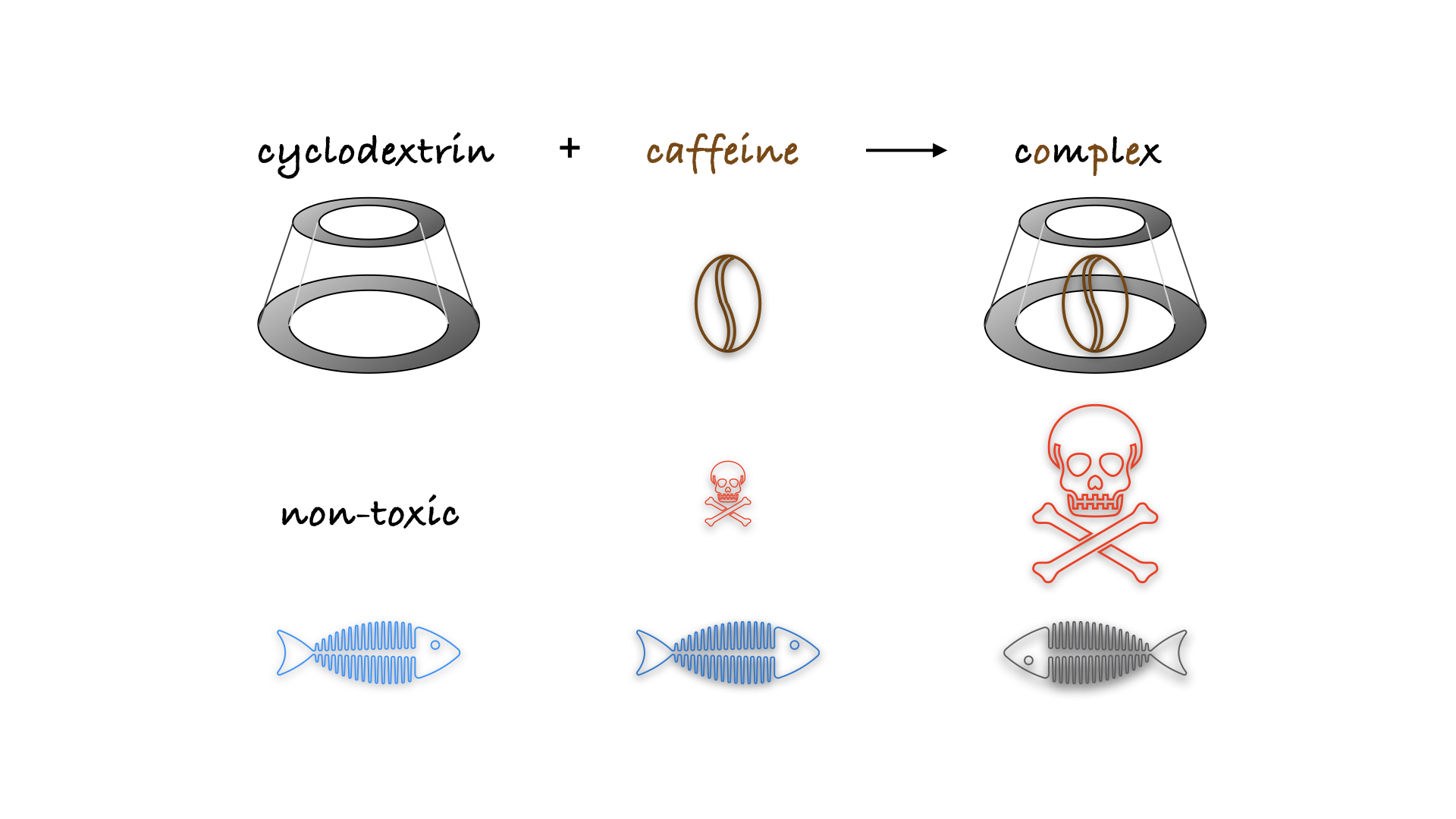
:
1. Introduction
2. Results and Discussion
2.1. Synthesis
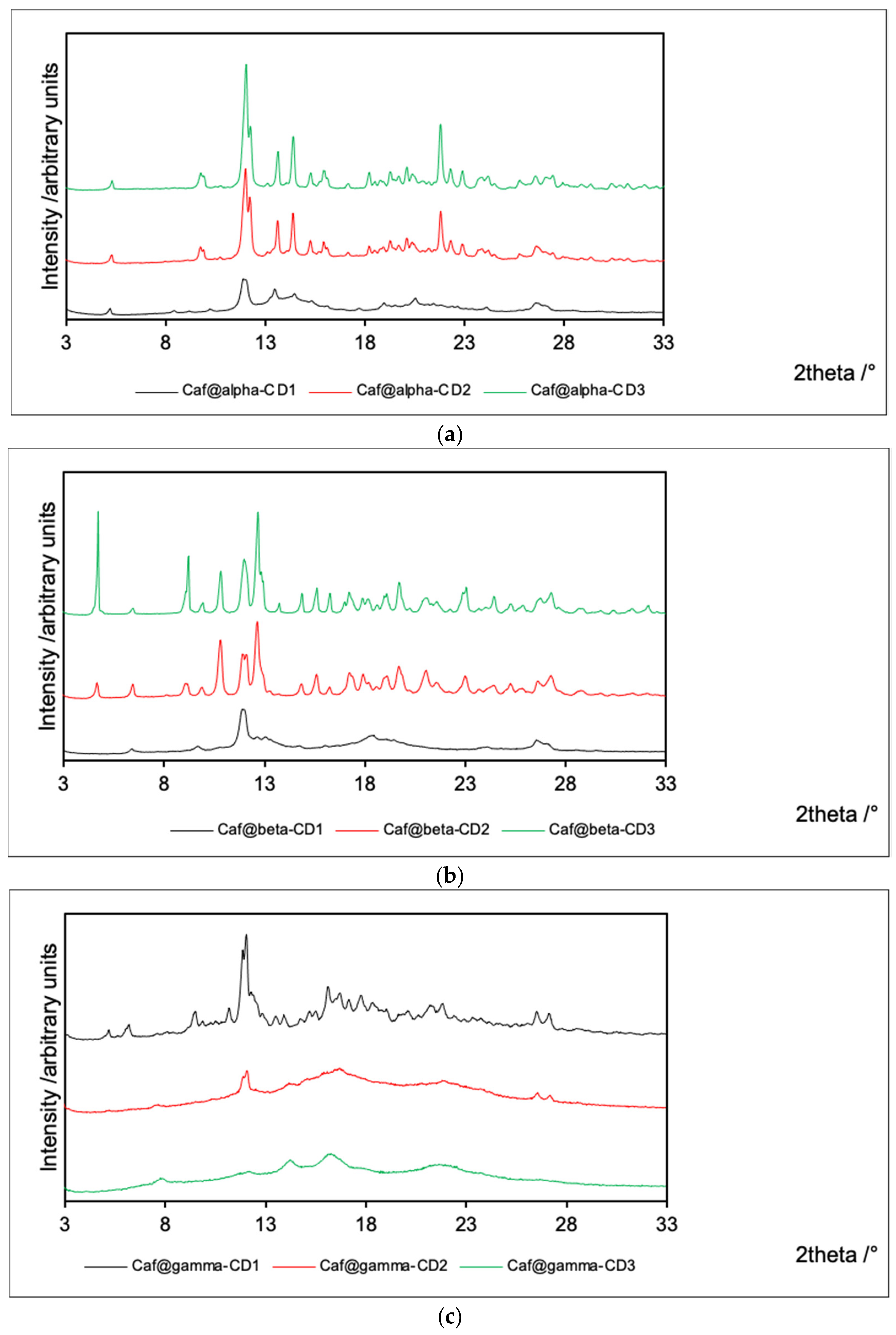
2.2. Powder X-ray Diffraction (PXRD)
2.3. Differential Scanning Calorimetry (DSC)
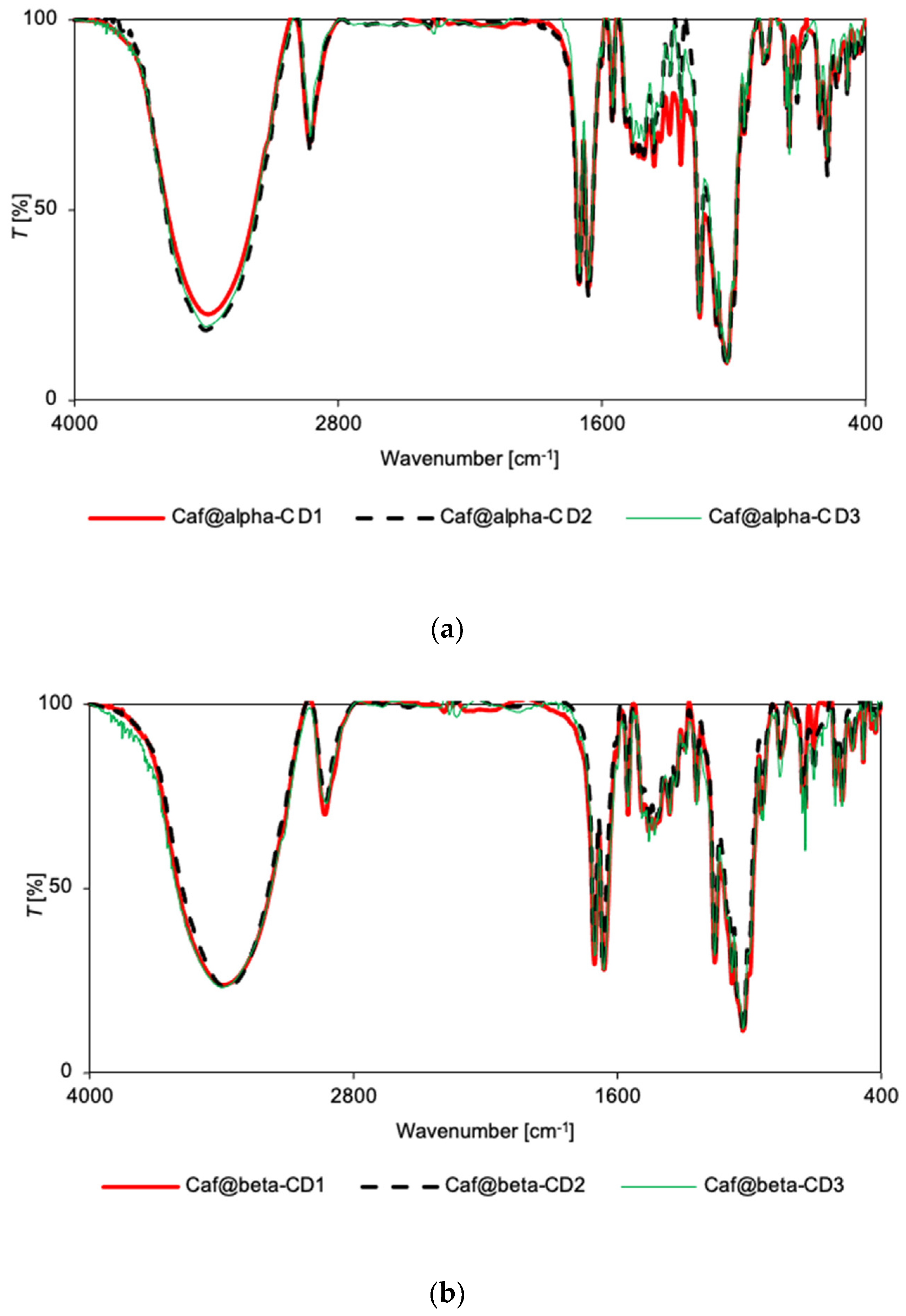
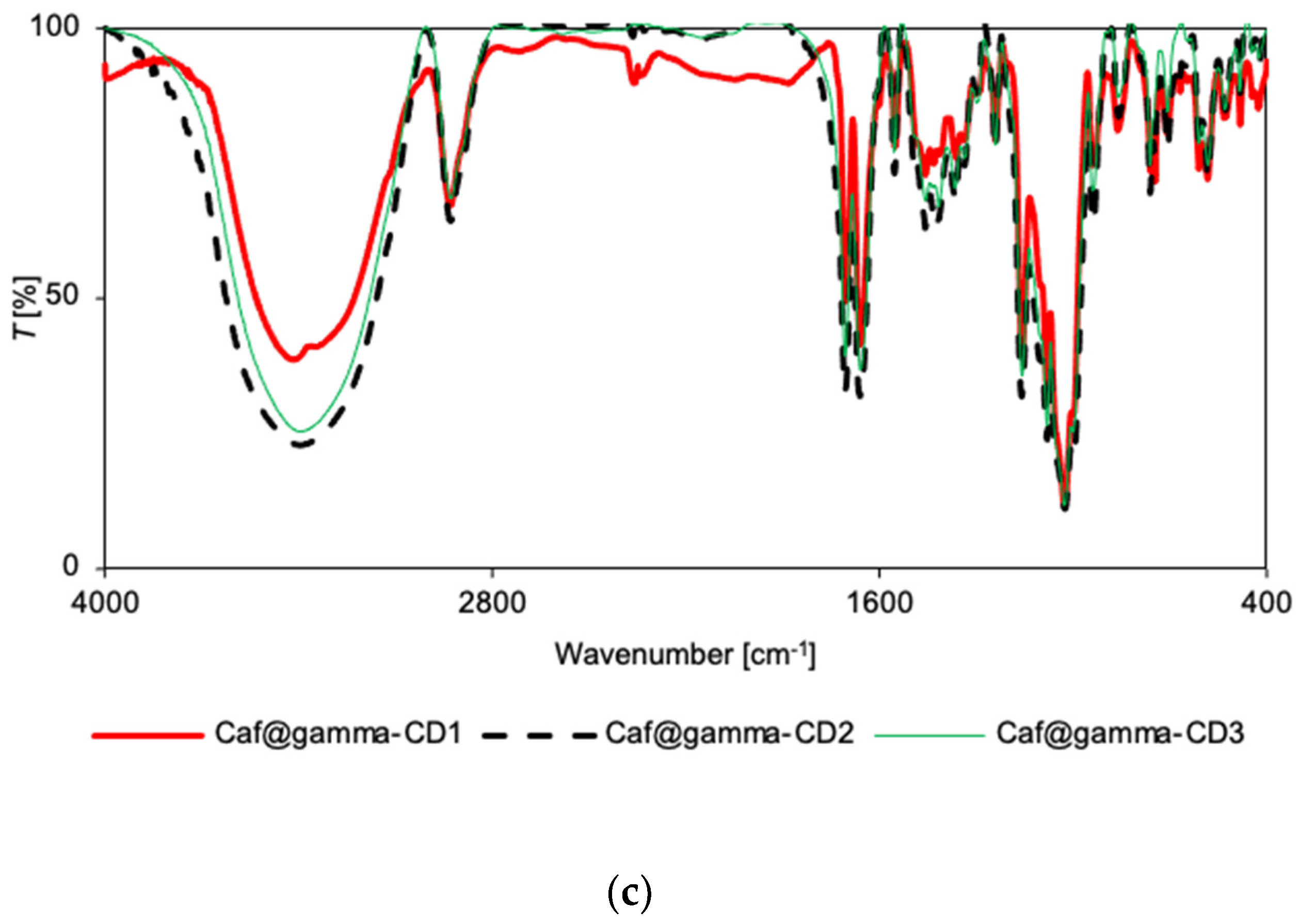
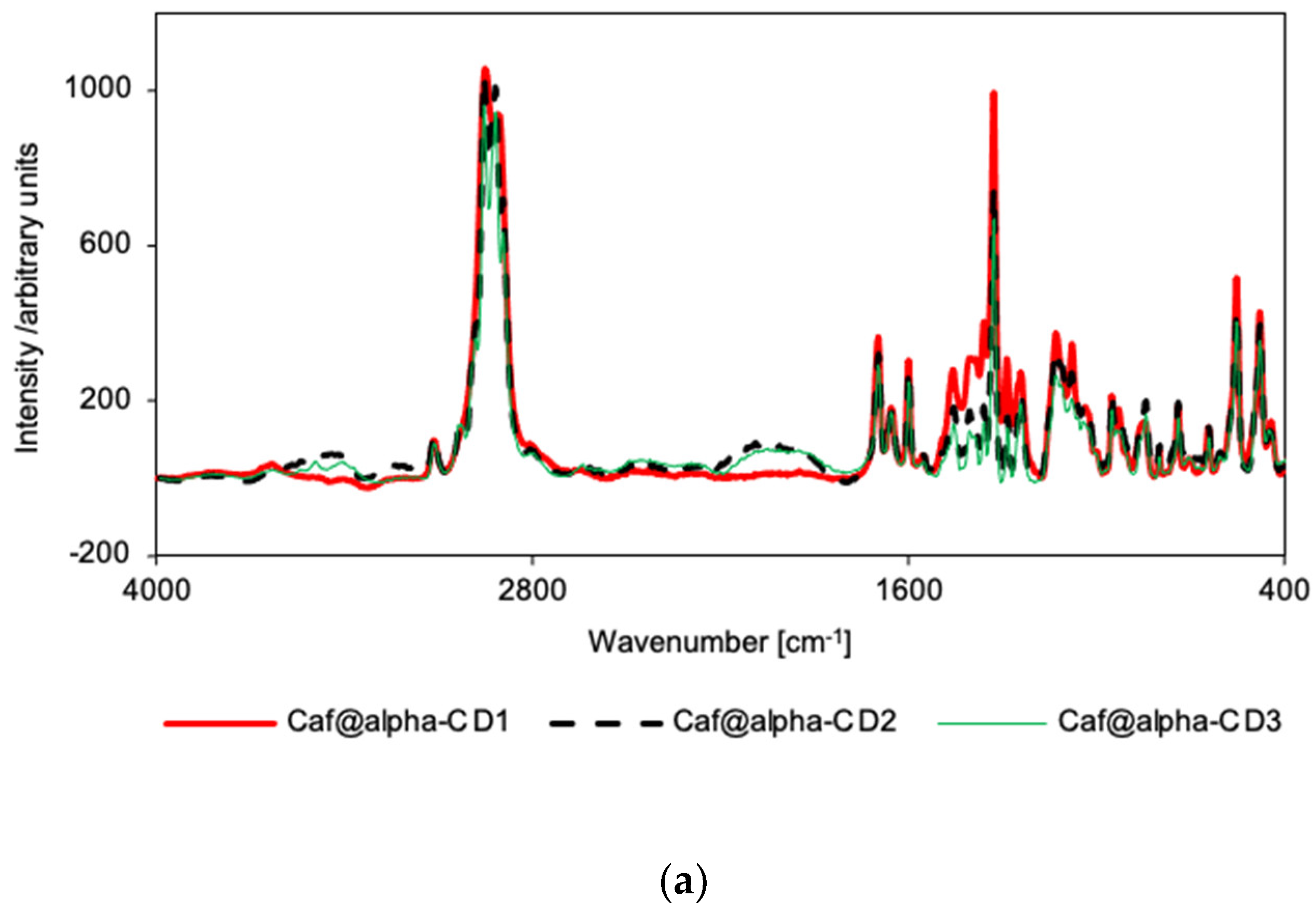
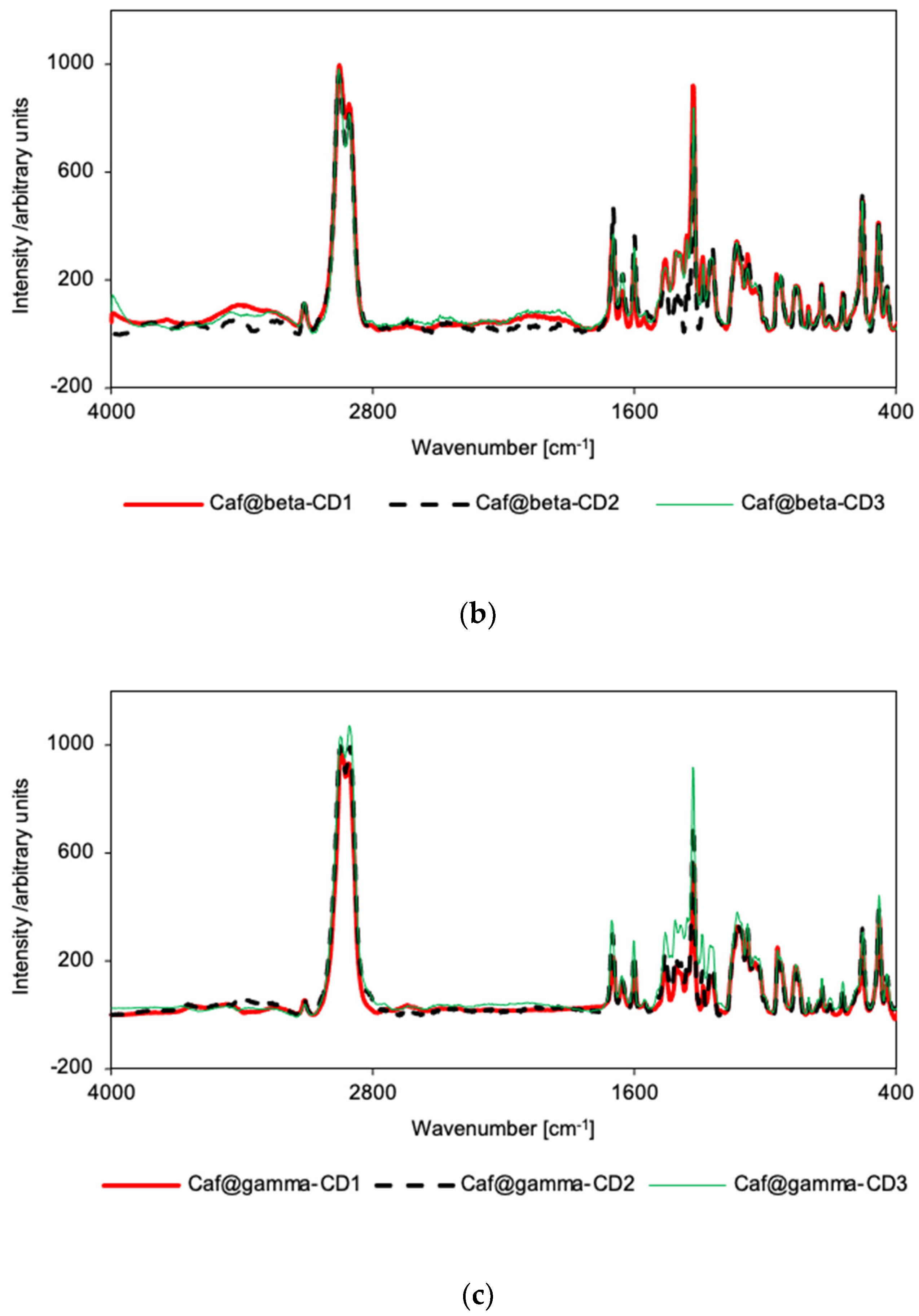
2.4. Vibrational Spectroscopy
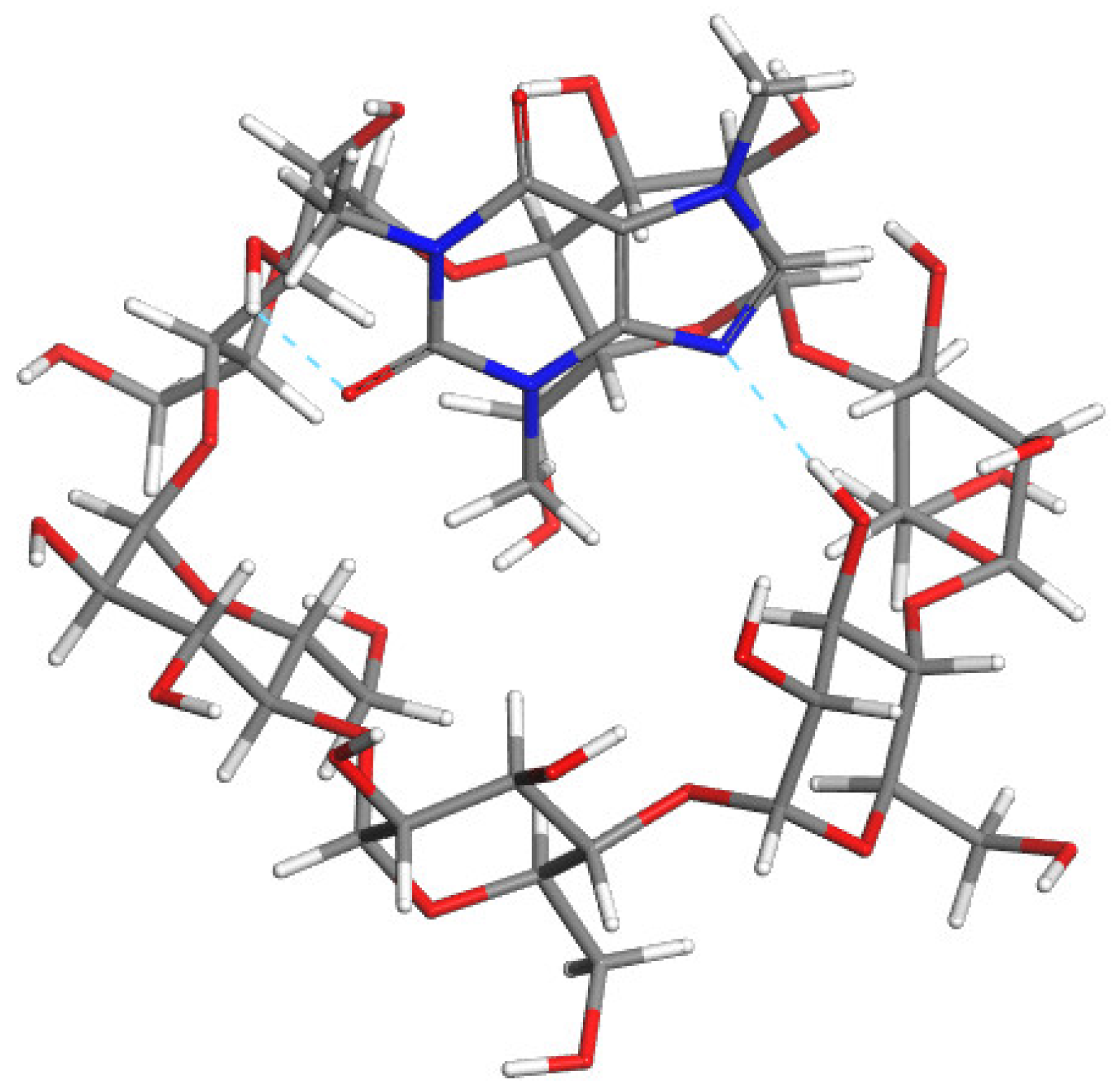
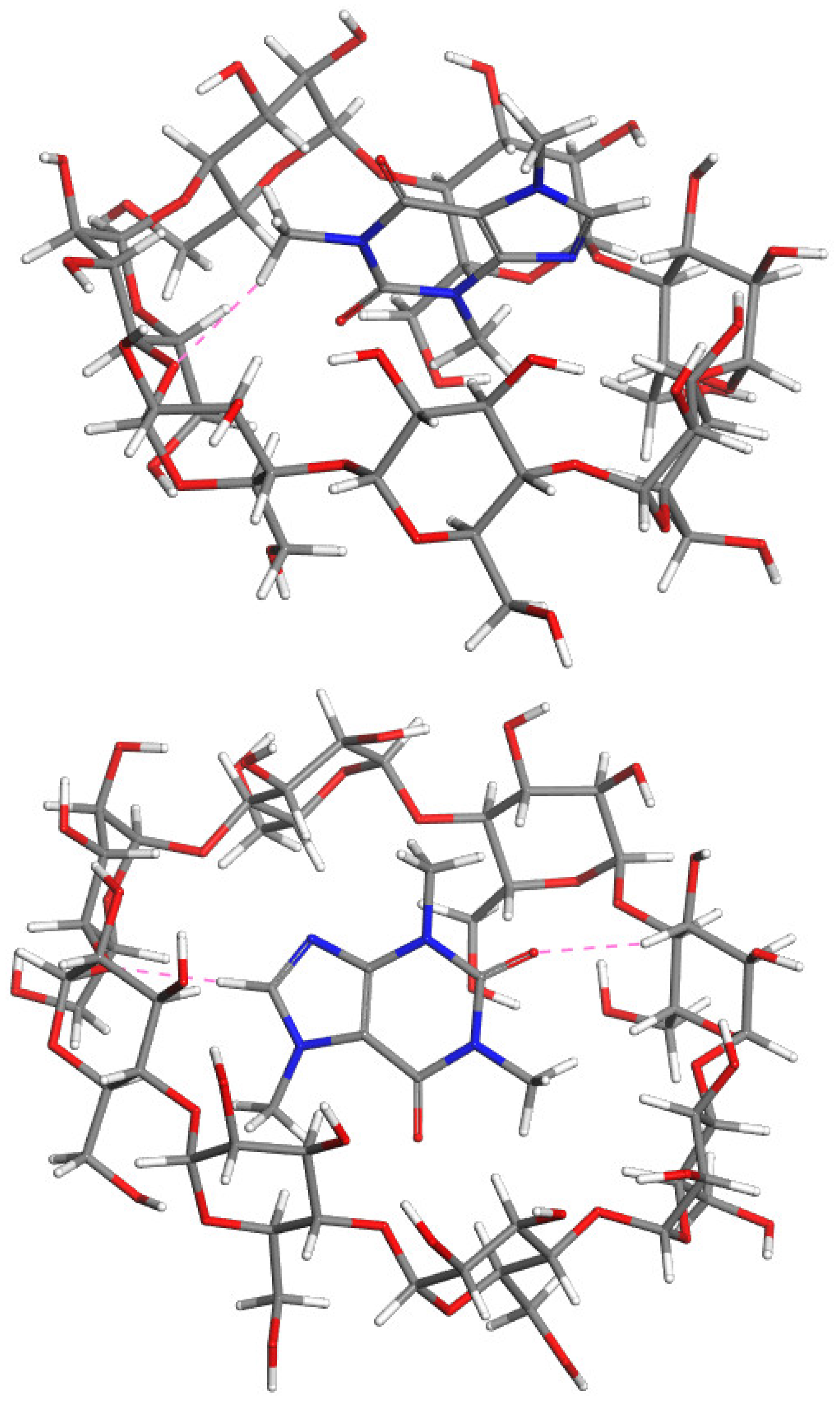
2.5. Molecular Modeling
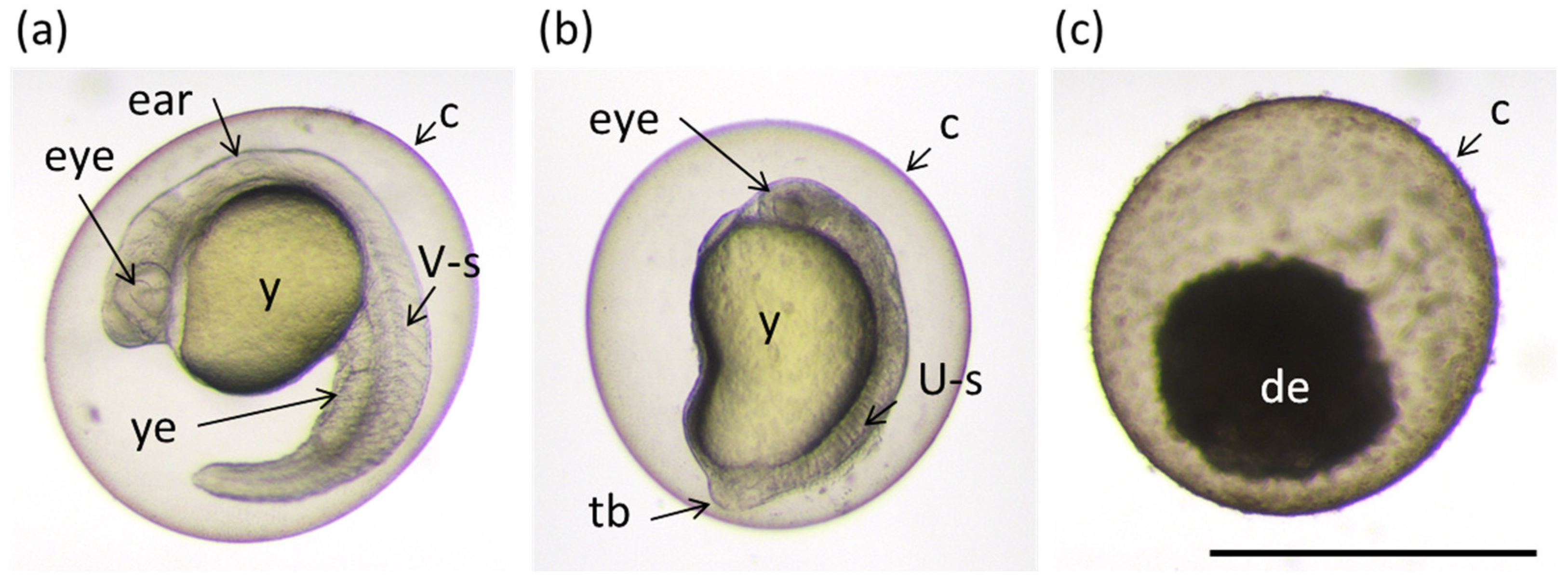
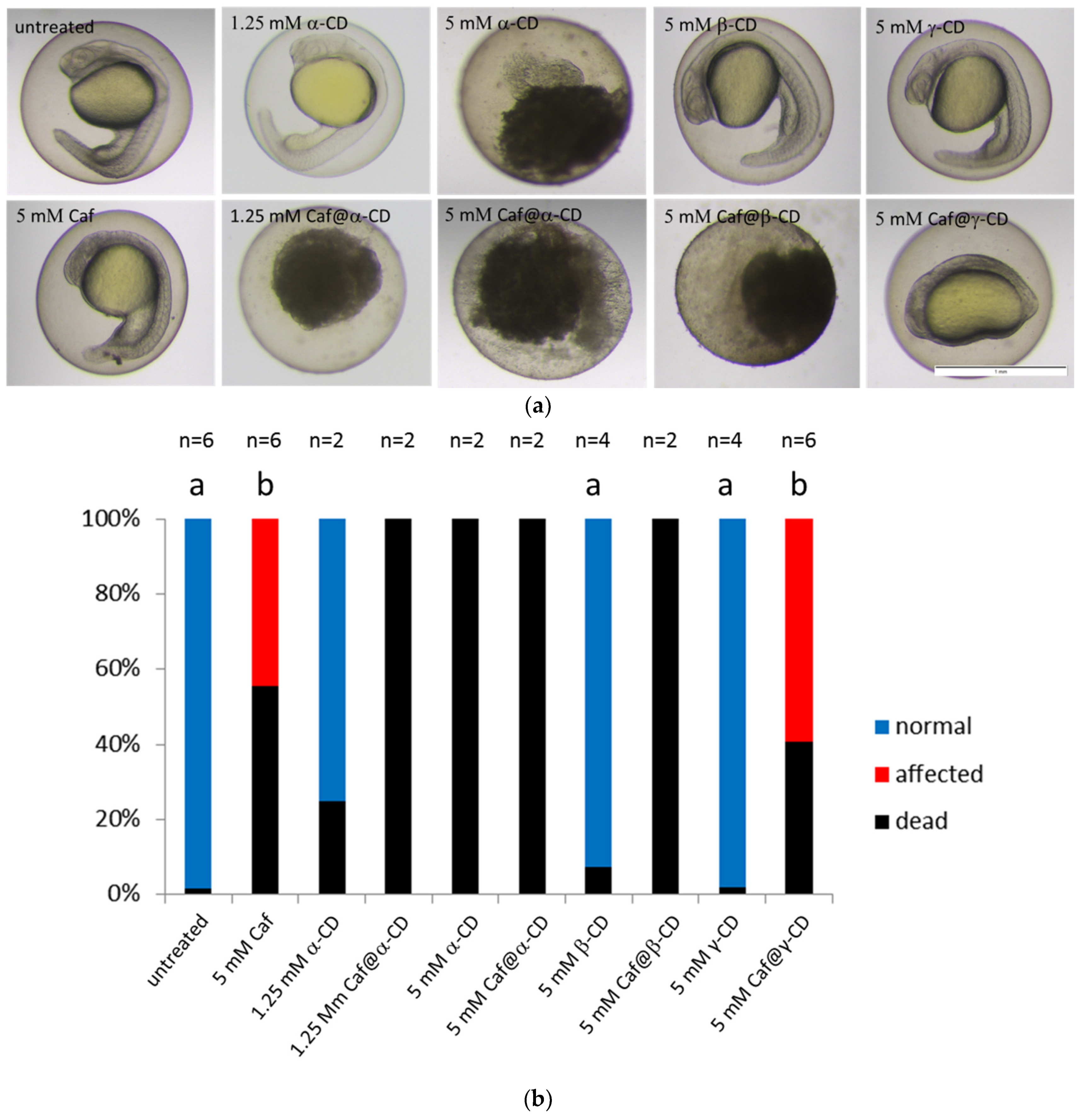
2.6. Toxicity
2.7. Caffeine Levels in the Zebrafish Embryos
3. Materials and Methods
3.1. Materials
3.2. Synthesis
3.3. PXRD
3.4. DSC
3.5. FT-IR
3.6. Raman Spectroscopy
3.7. Molecular Modeling
3.8. Toxicity
3.9. Caffeine Uptake
3.10. Ethical Statement
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Heckman, M.A.; Weil, J.; De Mejia, E.G. Caffeine (1, 3, 7-trimethylxanthine) in Foods: A Comprehensive Review on Consumption, Functionality, Safety, and Regulatory Matters. J. Food Sci. 2010, 75, R77–R87. [Google Scholar] [CrossRef]
- Nehlig, A. Interindividual Differences in Caffeine Metabolism and Factors Driving Caffeine Consumption. Pharmacol. Rev. 2018, 70, 384–411. [Google Scholar] [CrossRef] [Green Version]
- Szejtli, J. Introduction and General Overview of Cyclodextrin Chemistry. Chem. Rev. 1998, 98, 1743–1754. [Google Scholar] [CrossRef]
- Duchêne, D. Cyclodextrins and Their Industrial Uses; Editions de Santé: Paris, France, 1987. [Google Scholar]
- Frömming, K.-H.; Szejtli, J. Cyclodextrins in Pharmacy; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Loftsson, T.; Duchêne, D. Cyclodextrins and their pharmaceutical applications. Int. J. Pharm. 2007, 329, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.-L.; Ding, L.-H.; Dong, C.; Niu, W.-P.; Shuang, S.-M. Study on inclusion complex of cyclodextrin with methyl xanthine derivatives by fluorimetry. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2003, 59, 2697–2703. [Google Scholar] [CrossRef]
- Mejri, M.; Bensouissi, A.; Aroulmoji, V.; Rogé, B. Hydration and self-association of caffeine molecules in aqueous solution: Comparative effects of sucrose and β-cyclodextrin. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2009, 73, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, O.V.; Terekhova, I.V.; Kumeev, R.S.; Al’per, G.A. Thermodynamic spectroscopic studies of interactions of alpha- and beta-cyclodextrins and 18-crown-6 ether with bases of nucleic acids. Russ. J. Phys. Chem. 2004, 78, 1488–1493. [Google Scholar]
- Rajendiran, N.; Mohandoss, T.; Thulasidhasan, J. Excimer emission of caffeine with α- and β-cyclodextrins: Spectral and molecular modelling studies. Phys. Chem. Liq. 2017, 55, 1–13. [Google Scholar] [CrossRef]
- Prabu, S.; Swaminathan, M.; Sivakumar, K.; Rajamohan, R. Preparation, characterization and molecular modeling studies of the inclusion complex of Caffeine with Beta-cyclodextrin. J. Mol. Struct. 2015, 1099, 616–624. [Google Scholar] [CrossRef]
- Másson, M.; Loftsson, T.; Stefansson, E. How Do Cyclodextrins Enhance Drug Permeability through Biological Membranes? Springer: Berlin/Heidelberg, Germany, 1999; pp. 363–366. [Google Scholar]
- Másson, M.; Loftsson, T.; Másson, G.; Stefánsson, E. Cyclodextrins as permeation enhancers: Some theoretical evaluations and in vitro testing. J. Control. Release 1999, 59, 107–118. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, B.; Sun, J. The Solubility-Permeability Trade-Off of Progesterone with Cyclodextrins Under Physiological Conditions: Experimental Observations and Computer Simulations. J. Pharm. Sci. 2018, 107, 488–494. [Google Scholar] [CrossRef]
- Muankaew, C.; Loftsson, T. Cyclodextrin-Based Formulations: A Non-Invasive Platform for Targeted Drug Delivery. Basic Clin. Pharmacol. Toxicol. 2017, 122, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Bradford, Y.M.; Toro, S.; Ramachandran, S.; Ruzicka, L.; Howe, D.G.; Eagle, A.; Kalita, P.; Martin, R.; Moxon, S.A.T.; Schaper, K.; et al. Zebrafish Models of Human Disease: Gaining Insight into Human Disease at ZFIN. ILAR J. 2017, 58, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, H.C.; Vacca, I.; Schoenmacker, G.; Cleal, M.; Tochwin, A.; O’Connor, B.; Young, A.M.; Vasquez, A.A.; Winter, M.J.; Parker, M.O.; et al. Screening for drugs to reduce zebrafish aggression identifies caffeine and sildenafil. Eur. Neuropsychopharmacol. 2020, 30, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Lantz-McPeak, S.; Guo, X.; Cuevas, E.; Dumas, M.; Newport, G.D.; Ali, S.F.; Paule, M.G.; Kanungo, J. Developmental toxicity assay using high content screening of zebrafish embryos. J. Appl. Toxicol. 2015, 35, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Maes, J.; Verlooy, L.; Buenafe, O.E.; De Witte, P.A.M.; Esguerra, C.V.; Crawford, A.D. Evaluation of 14 Organic Solvents and Carriers for Screening Applications in Zebrafish Embryos and Larvae. PLoS ONE 2012, 7, e43850. [Google Scholar] [CrossRef]
- Da Silva, M.C.G.; da Silva, J.F.; Santos, T.P.; da Silva, N.P.C.; dos Santos, A.R.; de Andrade, A.L.C.; Souza, E.H.L.D.S.; Cadena, M.R.S.; de Sá, F.B.; Junior, V.A.D.S.; et al. The complexation of steroid hormones into cyclodextrin alters the toxic effects on the biological parameters of zebrafish (Danio rerio). Chemosphere 2019, 214, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Aigner, Z.; Berkesi, O.; Farkas, G.; Szabó-Révész, P. DSC, X-ray and FTIR studies of a gemfibrozil/dimethyl-β-cyclodextrin inclusion complex produced by co-grinding. J. Pharm. Biomed. Anal. 2012, 57, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-Y.; Hsu, C.-H.; Sheu, M.-T. Curve-fitting FTIR studies of loratadine/hydroxypropyl-β-cyclodextrin inclusion complex induced by co-grinding process. J. Pharm. Biomed. Anal. 2010, 53, 799–803. [Google Scholar] [CrossRef]
- Mura, P. Analytical techniques for characterization of cyclodextrin complexes in the solid state: A review. J. Pharm. Biomed. Anal. 2015, 113, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Mura, P.; Bettinetti, G.P.; Faucci, M.T.; Sorrenti, M.; Negri, A. Interaction of Naproxen with Crystalline and Amorphous Methylated β-Cyclodextrin in the Liquid and Solid State. Supramol. Chem. 2001, 12, 379–389. [Google Scholar] [CrossRef]
- Figueiras, A.; Carvalho, R.A.; Ribeiro, L.; Torres-Labandeira, J.J.; Veiga, F.J.B. Solid-state characterization and dissolution profiles of the inclusion complexes of omeprazole with native and chemically modified β-cyclodextrin. Eur. J. Pharm. Biopharm. 2007, 67, 531–539. [Google Scholar] [CrossRef] [PubMed]
- McMullan, R.; Saenger, W.; Fayos, J.; Mootz, D. Topography of cyclodextrin inclusion complexes. Carbohydr. Res. 1973, 31, 211–227. [Google Scholar] [CrossRef]
- Christoforides, E.; Papaioannou, A.; Bethanis, K. Crystal structure of the inclusion complex of cholesterol in β-cyclodextrin and molecular dynamics studies. Beilstein J. Org. Chem. 2018, 14, 838–848. [Google Scholar] [CrossRef]
- Ding, J.; Steiner, T.; Saenger, W. Structure of the γ-cyclodextrin–1-propanol–17H2O inclusion complex. Acta Crystallogr. Sect. B Struct. Sci. 1991, 47, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Mura, P.; Zerrouk, N.; Faucci, M.; Maestrelli, F.; Chemtob, C. Comparative study of ibuproxam complexation with amorphous β-cyclodextrin derivatives in solution and in the solid state. Eur. J. Pharm. Biopharm. 2002, 54, 181–191. [Google Scholar] [CrossRef]
- Bettinetti, G.; Gazzaniga, A.; Mura, P.; Giordano, F.; Setti, M. Thermal behaviour and dissolution properties of naproxen in combinations with chemically modified ß-Cyclodextrins. Drug Dev. Ind. Pharm. 1992, 18, 39–53. [Google Scholar] [CrossRef]
- Ucun, F.; Sağlam, A.; Güçlü, V. Molecular structures and vibrational frequencies of xanthine and its methyl derivatives (caffeine and theobromine) by ab initio Hartree–Fock and density functional theory calculations. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2007, 67, 342–349. [Google Scholar] [CrossRef]
- Nolasco, M.M.; Amado, A.M.; Ribeiro-Claro, P.J.A. Computationally-Assisted Approach to the Vibrational Spectra of Molecular Crystals: Study of Hydrogen-Bonding and Pseudo-Polymorphism. ChemPhysChem 2006, 7, 2150–2161. [Google Scholar] [CrossRef] [Green Version]
- Rezende, B.A.; Cortes, S.F.; De Sousa, F.B.; Lula, I.S.; Schmitt, M.; Sinisterra, R.D.; Lemos, V.S. Complexation with β-cyclodextrin confers oral activity on the flavonoid dioclein. Int. J. Pharm. 2009, 367, 133–139. [Google Scholar] [CrossRef]
- Yadav, V.R.; Suresh, S.; Devi, K.; Yadav, S. Effect of Cyclodextrin Complexation of Curcumin on its Solubility and Antiangiogenic and Anti-inflammatory Activity in Rat Colitis Model. AAPS PharmSciTech 2009, 10, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Aree, T. Supramolecular Complexes of β-Cyclodextrin with Clomipramine and Doxepin: Effect of the Ring Substituent and Component of Drugs on Their Inclusion Topologies and Structural Flexibilities. Pharmaceuticals 2020, 13, 278. [Google Scholar] [CrossRef]
- Buta, M.C.; Toader, A.M.; Frecus, B.; Oprea, C.I.; Cimpoesu, F.; Ionita, G. Molecular and Supramolecular Interactions in Systems with Nitroxide-Based Radicals. Int. J. Mol. Sci. 2019, 20, 4733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghania, A.; Bahlakeha, G.; Ramezanzadehb, B. Beta-cyclodextrin-zinc acetylacetonate (β-CD@ZnA) inclusion complex formation as a sustainable/smart nanocarrier of corrosion inhibitors for a water-based siliconized composite film: Integrated experimental analysis and fundamental computational electronic/atomic-scale simulation. Compos. Part B Eng. 2020, 197, 108152. [Google Scholar] [CrossRef]
- Kerdpol, K.; Daengngern, R.; Sattayanon, C.; Namuangruk, S.; Rungrotmongkol, T.; Wolschann, P.; Kungwan, N.; Hannongbua, S. Effect of Water Microsolvation on the Excited-State Proton Transfer of 3-Hydroxyflavone Enclosed in γ-Cyclodextrin. Molecules 2021, 26, 843. [Google Scholar] [CrossRef]
- Oqmhula, K.; Hongo, K.; Maezono, R.; Ichibha, T. Ab Initio Evaluation of Complexation Energies for Cyclodextrin-Drug Inclusion Complexes. ACS Omega 2020, 5, 19371–19376. [Google Scholar] [CrossRef] [PubMed]
- Feng, B. Furrow-Specific Endocytosis during Cytokinesis of Zebrafish Blastomeres. Exp. Cell Res. 2002, 279, 14–20. [Google Scholar] [CrossRef]
- Radi, M.; Evensen, L.; Dreassi, E.; Zamperini, C.; Caporicci, M.; Falchi, F.; Musumeci, F.; Schenone, S.; Lorens, J.B.; Botta, M. A combined targeted/phenotypic approach for the identification of new antiangiogenics agents active on a zebrafish model: From in silico screening to cyclodextrin formulation. Bioorg. Med. Chem. Lett. 2012, 22, 5579–5583. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-Y.; Ma, K.; Cheng, R.; She, X.-J.; Zhang, Y.-W.; Wang, C.-F.; Chen, S.; Xu, C. Host-guest supramolecular assembly directing beta-cyclodextrin based nanocrystals towards their robust performances. J. Hazard. Mater. 2019, 361, 329–337. [Google Scholar] [CrossRef]
- Geng, Q.; Xie, J.; Wang, X.; Cai, M.; Ma, H.; Ni, H. Preparation and Characterization of Butachlor/(2-Hydroxypropyl)-β-cyclodextrin Inclusion Complex: Improve Soil Mobility and Herbicidal Activity and Decrease Fish Toxicity. J. Agric. Food Chem. 2018, 66, 12198–12205. [Google Scholar] [CrossRef]
- Saokham, P.; Muankaew, C.; Jansook, P.; Loftsson, T. Solubility of Cyclodextrins and Drug/Cyclodextrin Complexes. Molecules 2018, 23, 1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groom, C.R.; Bruno, I.J.; Lightfoot, M.P.; Ward, S.C. The Cambridge Structural Database. Acta Crystallogr. Sect. B Struct. Sci. Cryst. Eng. Mater. 2016, 72, 171–179. [Google Scholar] [CrossRef]
- Enright, G.D.; Terskikh, V.V.; Brouwer, A.D.H.; Ripmeester, J.A. The Structure of Two Anhydrous Polymorphs of Caffeine from Single-Crystal Diffraction and Ultrahigh-Field Solid-State13C NMR Spectroscopy. Cryst. Growth Des. 2007, 7, 1406–1410. [Google Scholar] [CrossRef]
- Manor, P.C.; Saenger, W. Topography of cyclodextrin inclusion complexes. III. Crystal and molecular structure of cyclohexaamylose hexahydrate, the water dimer inclusion complex. J. Am. Chem. Soc. 1974, 96, 3630–3639. [Google Scholar] [CrossRef]
- Lindner, K.; Saenger, W. Crystal and molecular structure of cyclohepta-amylose dodecahydrate. Carbohydr. Res. 1982, 99, 103–115. [Google Scholar] [CrossRef]
- Harata, K. The Structure of the Cyclodextrin Complex. XX. Crystal Structure of Uncomplexed Hydrated γ-Cyclodextrin. Bull. Chem. Soc. Jpn. 1987, 60, 2763–2767. [Google Scholar] [CrossRef] [Green Version]
- Materials Studio, 8.0; BIOVIA, Dassault Systèmes: San Diego, CA, USA, 2020.
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16 Rev. C.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Barone, V.; Cossi, M.; Tomasi, J. A new definition of cavities for the computation of solvation free energies by the polarizable continuum model. J. Chem. Phys. 1997, 107, 3210–3221. [Google Scholar] [CrossRef]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) Test. OECD iLibrary 2013. [Google Scholar] [CrossRef]
- Brusnikina, M.; Silyukov, O.; Chislov, M.; Volkova, T.; Proshin, A.; Mazur, A.; Tolstoy, P.; Terekhova, I. Effect of cyclodextrin complexation on solubility of novel anti-Alzheimer 1,2,4-thiadiazole derivative. J. Therm. Anal. Calorim. 2017, 130, 443–450. [Google Scholar] [CrossRef]
- Li, S.; Lin, X.; Xu, K.; He, J.; Yang, H.; Li, H. Co-grinding Effect on Crystalline Zaltoprofen with β-cyclodextrin/Cucurbit [7]uril in Tablet Formulation. Sci. Rep. 2017, 7, 45984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Promzeleva, M.; Volkova, T.V.; Proshin, A.N.; Siluykov, O.; Mazur, A.; Tolstoy, P.M.; Ivanov, S.P.; Kamilov, F.; Terekhova, I.V. Improved Biopharmaceutical Properties of Oral Formulations of 1,2,4-Thiadiazole Derivative with Cyclodextrins: In Vitro and in Vivo Evaluation. ACS Biomater. Sci. Eng. 2017, 4, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Jug, M.; Mura, P.A. Grinding as Solvent-Free Green Chemistry Approach for Cyclodextrin Inclusion Complex Preparation in the Solid State. Pharmaceutics 2018, 10, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance (Method of Preparation) | α-CD | β-CD | γ-CD | |
|---|---|---|---|---|
| single component (ground, dry) | α-CD1 | β-CD1 | γ-CD1 | |
| single component (deposited from soln.) | α-CD3 | β-CD3 | γ-CD3 | |
| putative complex (coground, dry) | Caf@α-CD1 | Caf@β-CD1 | Caf@γ-CD1 | |
| putative complex (coground, w/water drops) | Caf@α-CD2 | Caf@β-CD2 | Caf@γ-CD2 | |
| putative complex (deposited from soln.) | Caf@α-CD3 | Caf@β-CD3 | Caf@γ-CD3 | |
| mixture (from dried CD) | Caf+α-CD1 | Caf+β-CD1 | Caf+γ-CD1 | |
| mixture (from deposited caffeine and CD) | Caf+α-CD3 | Caf+β-CD3 | Caf+γ-CD3 | |
| caffeine (as supplied) | Caf1 | caffeine (deposited from soln.) | Caf3 | |
| Substance | Temperatures and Heats | Figures (Supplementary Material) | ||||
|---|---|---|---|---|---|---|
| Caf1 | 148-169-159 (31) | 233-248-239 (195) | S4, S9, and S14 | |||
| Caf3 | 135-165-152 (24.5) | 230-244-238 (193) | S6, S11, and S16 | |||
| α-CD1 | 66-86-76 (16) | 157-180-158 (8) | 180-241-184 (270) | S4 | ||
| Caf+α-CD1 | 57-70-63 (1.9) | 133-174-152 (124) | 234-242-236 (23) | S4 and S5 | ||
| Caf@α-CD1 | 53-65-50 (0.5) | 139-170-148 (11) | 203-215-209 (11) * | 230-241-234 (34) | S5 and S8 | |
| Caf@α-CD2 | 83-104-92 (30) | 152-172-156 (36) | 176-204-182 (143) | 224-239-233 (28) | S8 | |
| α-CD3 | 78-99-89 (52) | 138-140-143 (2.3) | 174-222-176 (256) | S6 | ||
| Caf+α-CD3 | 78-98-88 (38) | 99-106-101 (2.2) | 141-147-142 (2.4) | 162-225-166 (267) | 229-245-235 (28) | S6 and S7 |
| Caf@α-CD3 | 78-94-87 (34) | 145-150-146 (2.3) | 154-184-160 (123) | 184-209-199 (29) | 223-237-231 (21) | S7 and S8 |
| β-CD1 | 162-187-163 (255) | S9 | ||||
| Caf+β-CD1 | 68-150-123 (310) | 104-150-123 (157) | 154-168-161 (1.8) | 235-241-237 (4.4) | 268-280-270 (16) | S9 and S10 |
| Caf@β-CD1 | 70-117-170 (265) | 208-224-209 (13) * | 225-243-232 (2.4) * | 250-279-265 (22) | S10 and S13 | |
| Caf@β-CD2 | 145-161-148 (44) | 178-196-175 (338) | 230-238-233 (9) | S13 | ||
| β-CD3 | 142-151-144 (7.5) | 175-220-192 (493) | S11 | |||
| Caf+β-CD3 | 141-188-143 (476) | 233-240-236 (8.6) | 257-271-265 (7) | S11 and S12 | ||
| Caf@β-CD3 | 147-171-153 (35) | 180-202-182 (250) | 232-239-235 (4) | S12 and S13 | ||
| γ-CD1 | 40-53-47 (1.7) | 184-189-185 (2.2) | 221-251-225 (80) | S14 | ||
| Caf+γ-CD1 | 150-161-152 (5) | 161-181-164 (89) | S14 and S15 | |||
| Caf@γ-CD1 | 163-174-164 (12) | 205-221-207(106) | S15 and S18 | |||
| Caf@γ-CD2 | 178-208-183 (178) | S18 | ||||
| γ-CD3 | 104-115-108 (4.1) | 132-162-147 (23.5) | 174-217-177 (413) | S16 | ||
| Caf+γ-CD3 | 96-110-105 (9.4) | 119-131-126 (7) | 148-157-149 (11) | 172-194-173 (290) | S16 and S17 | |
| Caf@γ-CD3 | 168-199-172 (150) | S17 and S18 | ||||
| ∆Ebind (kcal-mol−1) | ∆Ebind/PCM (kcal-mol−1) | |
|---|---|---|
| Caf@α-CD | −12.82 | −8.56 |
| Caf@β-CD | −25.88 | −20.04 |
| Caf@γ-CD | −30.08 | −34.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szmeja, S.; Gubica, T.; Ostrowski, A.; Zalewska, A.; Szeleszczuk, Ł.; Zawada, K.; Zielińska-Pisklak, M.; Skowronek, K.; Wiweger, M. Caffeine-Cyclodextrin Complexes as Solids: Synthesis, Biological and Physicochemical Characterization. Int. J. Mol. Sci. 2021, 22, 4191. https://doi.org/10.3390/ijms22084191
Szmeja S, Gubica T, Ostrowski A, Zalewska A, Szeleszczuk Ł, Zawada K, Zielińska-Pisklak M, Skowronek K, Wiweger M. Caffeine-Cyclodextrin Complexes as Solids: Synthesis, Biological and Physicochemical Characterization. International Journal of Molecular Sciences. 2021; 22(8):4191. https://doi.org/10.3390/ijms22084191
Chicago/Turabian StyleSzmeja, Sebastian, Tomasz Gubica, Andrzej Ostrowski, Aldona Zalewska, Łukasz Szeleszczuk, Katarzyna Zawada, Monika Zielińska-Pisklak, Krzysztof Skowronek, and Małgorzata Wiweger. 2021. "Caffeine-Cyclodextrin Complexes as Solids: Synthesis, Biological and Physicochemical Characterization" International Journal of Molecular Sciences 22, no. 8: 4191. https://doi.org/10.3390/ijms22084191
APA StyleSzmeja, S., Gubica, T., Ostrowski, A., Zalewska, A., Szeleszczuk, Ł., Zawada, K., Zielińska-Pisklak, M., Skowronek, K., & Wiweger, M. (2021). Caffeine-Cyclodextrin Complexes as Solids: Synthesis, Biological and Physicochemical Characterization. International Journal of Molecular Sciences, 22(8), 4191. https://doi.org/10.3390/ijms22084191