
Estrogen-Related Receptor Influences the Hemolymph Glucose Content by Regulating Midgut Trehalase Gene Expression in the Last Instar Larvae of Bombyx mori

 and
and 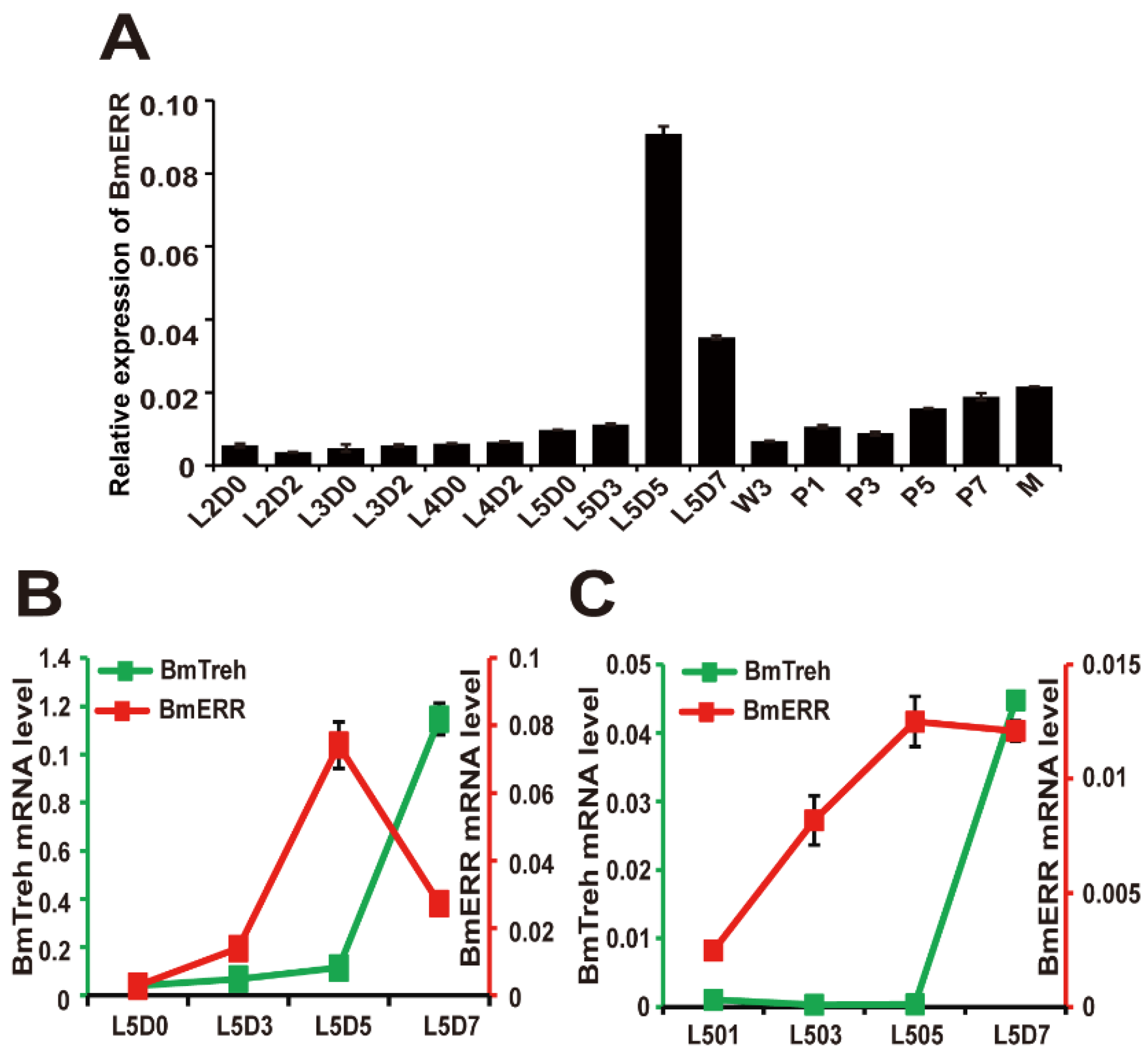{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Expression Pattern of ERR and Treh in Silkworms
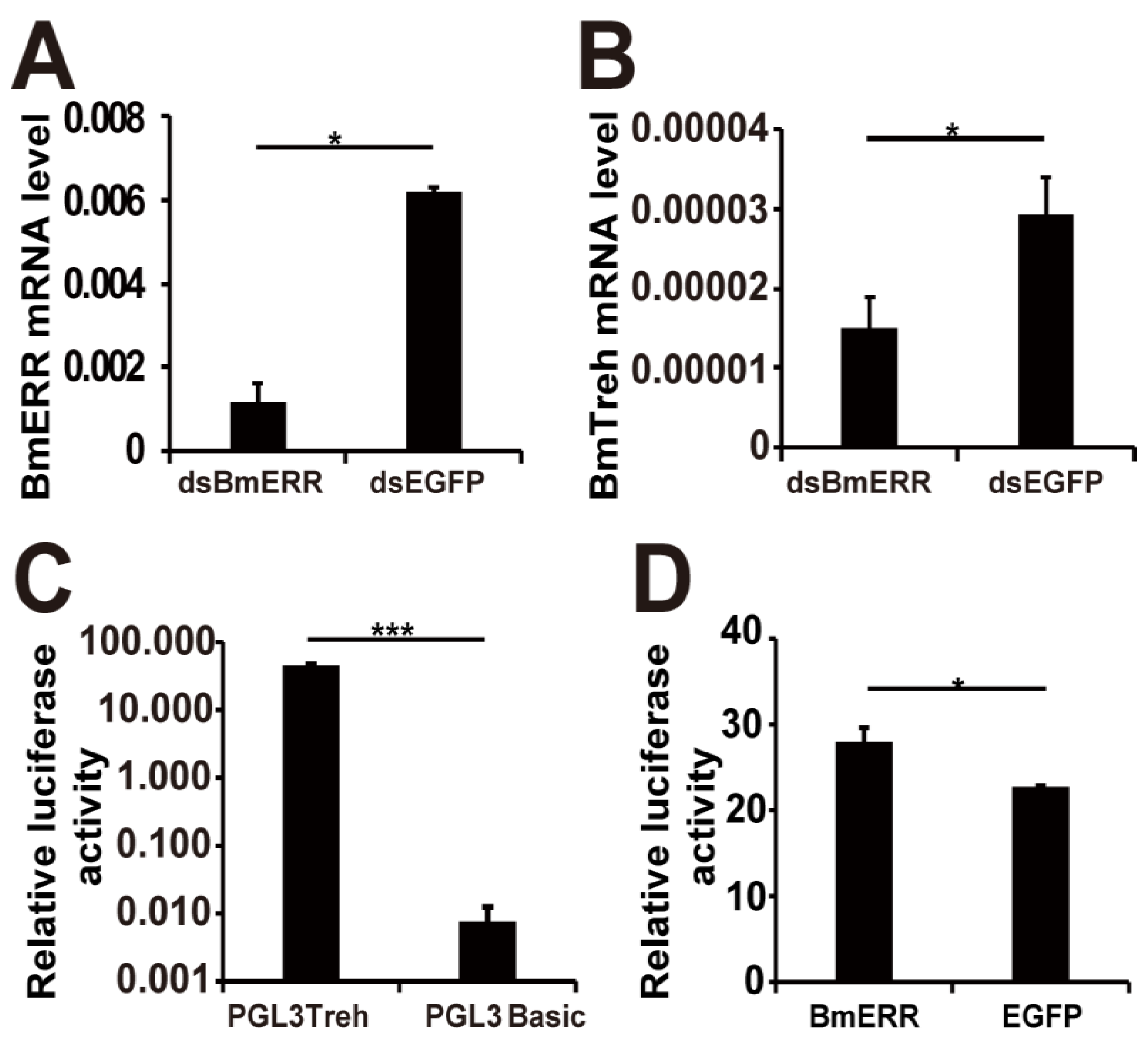
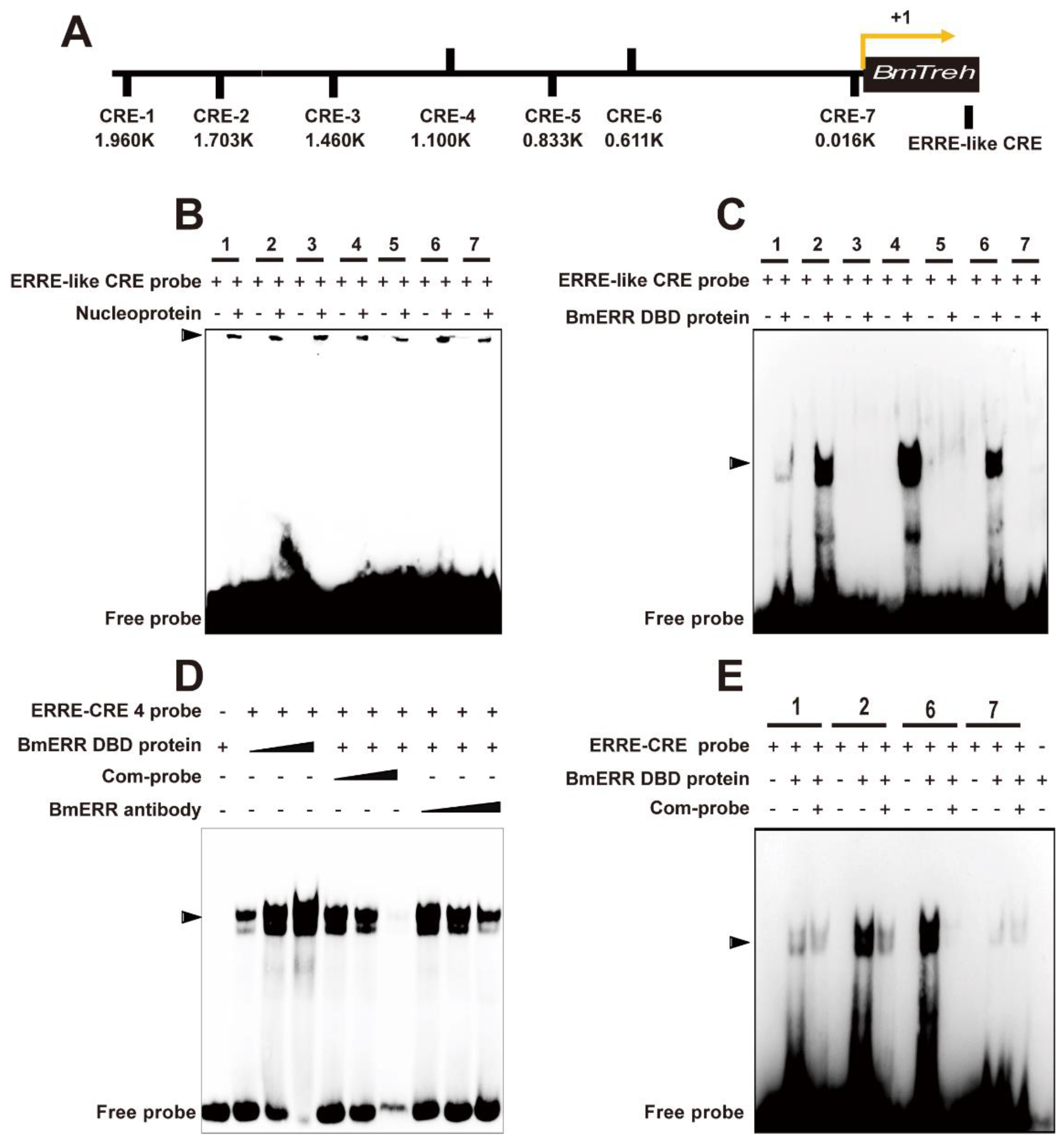
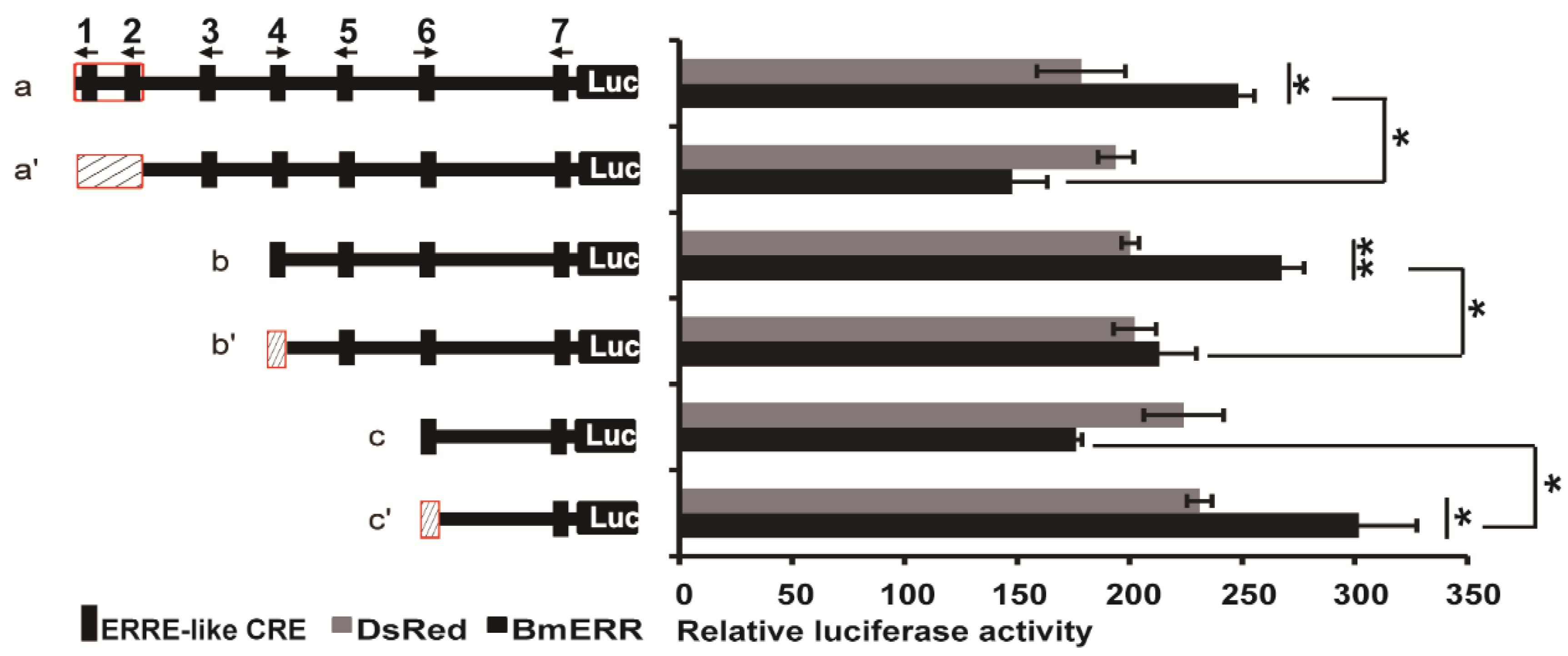
2.2. BmERR Regulated Treh Expression
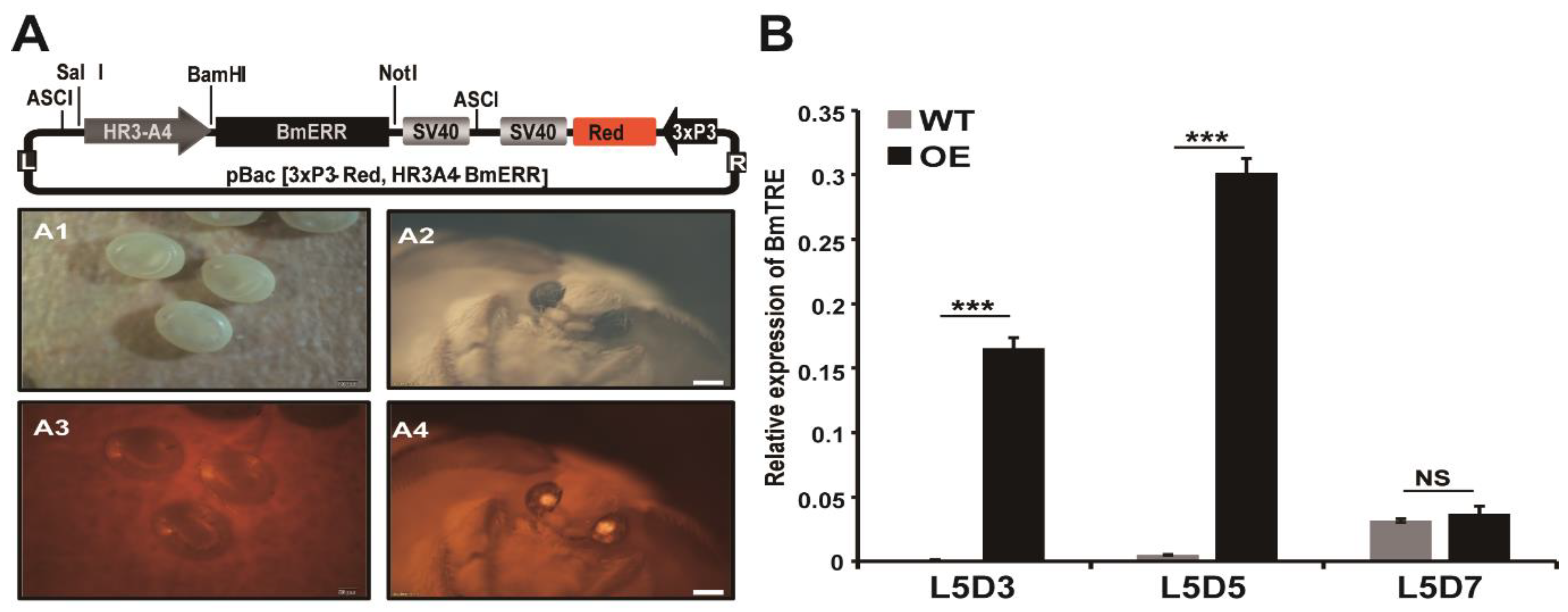
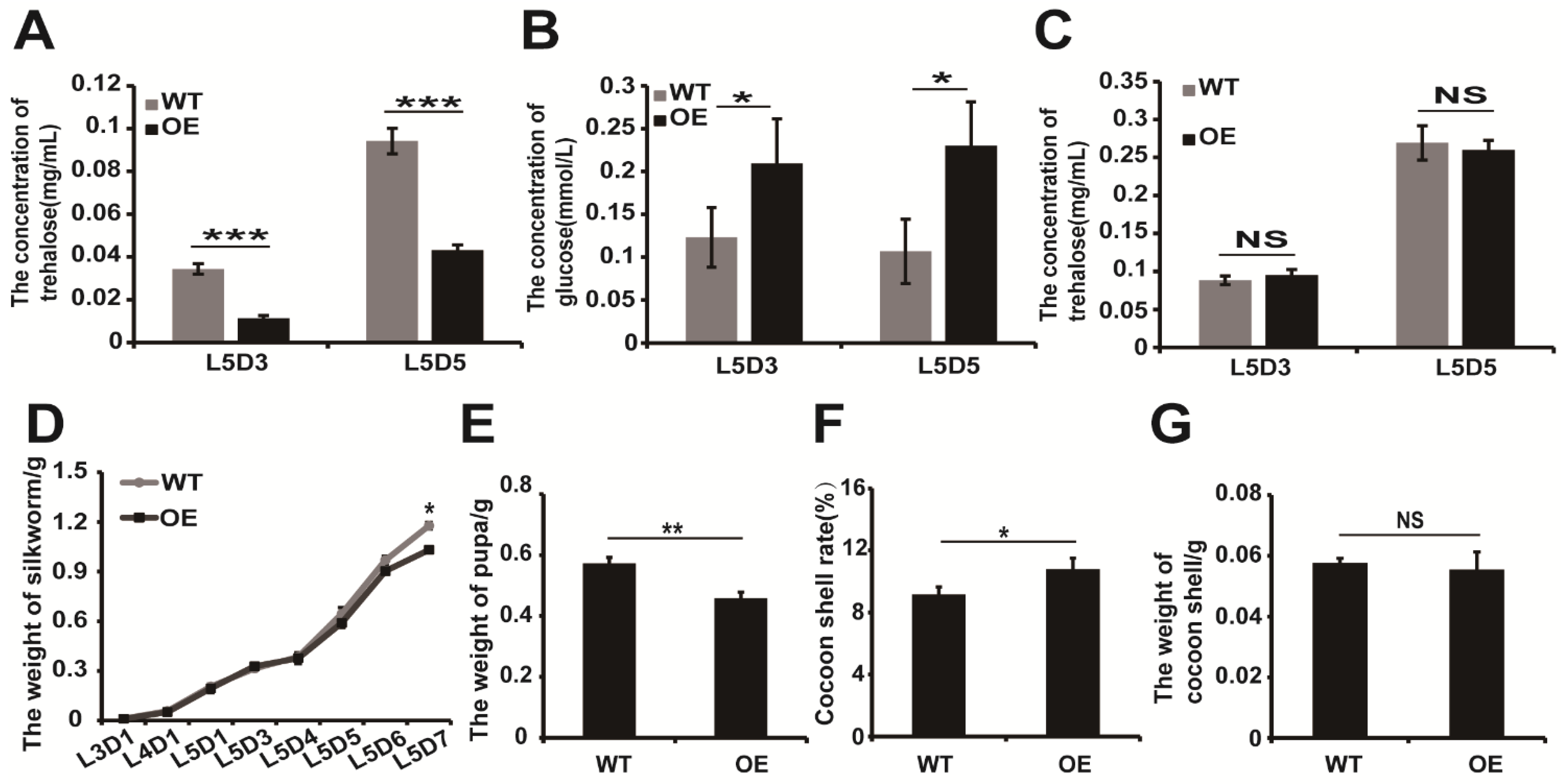
2.3. Overexpression of BmERR in Transgenic Species Increases the Expression of BmTreh and Hemolymph Glucose Content
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. RNA/DNA Extraction and cDNA Synthesis
4.3. qRT-PCR
4.4. Bioinformatics Analysis and Vector Construction
4.5. Cell Transfection, Luciferase Assay, and Double-Stranded RNA Interference Assay
4.6. EMSA
4.7. Construction of Transgenic Silkworms
4.8. Weight Statistics
4.9. Measurement of Trehalose and Glucose Levels
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Horard, B.; Vanacker, J.M. Estrogen receptor-related receptors: Orphan receptors desperately seeking a ligand. J. Mol. Endocrinol. 2003, 31, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giguere, V.; Yang, N.; Segui, P.; Evans, R.M. Identification of a New Class of Steroid-Hormone Receptors. Nature 1988, 331, 91–94. [Google Scholar] [CrossRef]
- Giguere, V. Orphan nuclear receptors: From gene to function. Endocr. Rev. 1999, 20, 689–725. [Google Scholar] [CrossRef]
- Bardet, P.L.; Obrecht-Pflumio, S.; Thisse, C.; Laudet, V.; Thisse, B.; Vanacker, J.M. Cloning and developmental expression of five estrogen-receptor related genes in the zebrafish. Dev. Genes Evol. 2004, 214, 240–249. [Google Scholar]
- Bardet, P.L.; Schubert, M.; Horard, B.; Holland, L.Z.; Laudet, V.; Holland, N.D.; Vanacker, J.M. Expression of estrogen-receptor related receptors in amphioxus and zebrafish: Implications for the evolution of posterior brain segmentation at the invertebrate-to-vertebrate transition. Evol. Dev. 2005, 7, 223–233. [Google Scholar] [CrossRef]
- Dehal, P.; Satou, Y.; Campbell, R.K.; Chapman, J.; Degnan, B.; De Tomaso, A.; Davidson, B.; Di Gregorio, A.; Gelpke, M.; Goodstein, D.M.; et al. The draft genome of Ciona intestinalis: Insights into chordate and vertebrate origins. Science 2002, 298, 2157–2167. [Google Scholar] [CrossRef] [Green Version]
- Howard-Ashby, M.; Materna, S.C.; Brown, C.T.; Chen, L.; Cameron, R.A.; Davidson, E.H. Gene families encoding transcription factors expressed in early development of Strongylocentrotus purpuratus. Dev. Biol. 2006, 300, 90–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasawa, K.; Treen, N.; Kondo, R.; Otoki, Y.; Itoh, N.; Rotchell, J.M.; Osada, M. Molecular characterization of an estrogen receptor and estrogen-related receptor and their autoregulatory capabilities in two Mytilus species. Gene 2015, 564, 153–159. [Google Scholar] [CrossRef]
- Bardet, P.L.; Laudet, V.; Vanacker, J.M. Studying non-mammalian models? Not a fool’s ERRand! Trends Endocrinol. Metab. TEM 2006, 17, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desvergne, B.; Michalik, L.; Wahli, W. Transcriptional regulation of metabolism. Physiol. Rev. 2006, 86, 465–514. [Google Scholar] [CrossRef]
- Giguere, V. Transcriptional Control of Energy Homeostasis by the Estrogen-Related Receptors. Endocr. Rev. 2008, 29, 677–696. [Google Scholar] [CrossRef] [Green Version]
- Greschik, H.; Wurtz, J.M.; Sanglier, S.; Bourguet, W.; van Dorsselaer, A.; Moras, D.; Renaud, J.P. Structural and functional evidence for ligand-independent transcriptional activation by the estrogen-related receptor 3. Mol. Cell 2002, 9, 303–313. [Google Scholar] [CrossRef]
- Sladek, R.; Giguere, V. Orphan nuclear receptors: An emerging family of metabolic regulators. Adv. Pharmacol. 2000, 47, 23–87. [Google Scholar]
- Hummasti, S.; Tontonoz, P. Adopting new orphans into the family of metabolic regulators. Mol. Endocrinol. 2008, 22, 1743–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, A.M.; Giguere, V. The NR3B subgroup: An ovERRview. Nucl. Recept. Signal. 2007, 5, e009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Lin, T.; Kamarajugadda, S.; Lu, J. Regulation of glycolysis and the Warburg effect by estrogen-related receptors. Oncogene 2013, 32, 2079–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennessen, J.M.; Baker, K.D.; Lam, G.; Evans, J.; Thummel, C.S. The Drosophila Estrogen-Related Receptor Directs a Metabolic Switch that Supports Developmental Growth. Cell Metab. 2011, 13, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Thornton, J.W.; Need, E.; Crews, D. Resurrecting the ancestral steroid receptor: Ancient origin of estrogen signaling. Science 2003, 301, 1714–1717. [Google Scholar] [CrossRef] [Green Version]
- Bozzolan, F.; Durand, N.; Demondion, E.; Bourgeois, T.; Gassias, E.; Debernard, S. Evidence for a role of oestrogen receptor-related receptor in the regulation of male sexual behaviour in the moth Agrotis ipsilon. Insect Mol. Biol. 2017, 26, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Xi, G.S. Morphological changes in different caste adult ant specificities of Polyrhachis vicina Roger (Hymenoptera, Formicidae) caused in estrogen-related receptor. Gen. Comp. Endocrinol. 2018, 266, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Pandey, A.K.; Gupta, S.; Kumar, A.; Khanna, P.; Shankar, J.; Ram, K.R. Estrogen related receptor is required for the testicular development and for the normal sperm axoneme/mitochondrial derivatives in Drosophila males. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Kwak, I.S. Molecular effects of endocrine-disrupting chemicals on the Chironomus riparius estrogen-related receptor gene. Chemosphere 2010, 79, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Paz, P.; Morales, M.; Urien, J.; Morcillo, G.; Martinez-Guitarte, J.L. Endocrine-related genes are altered by antibacterial agent triclosan in Chironomus riparius aquatic larvae. Ecotoxicol. Environ. Saf. 2017, 140, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, M.; Zhang, G.; Liu, F.; Wang, H.; Guo, X.; Xu, B. Molecular cloning, expression, and stress response of the estrogen-related receptor gene (AccERR) from Apis cerana cerana. Die Nat. 2016, 103, 24. [Google Scholar] [CrossRef]
- In, S.; Cho, H.; Lee, K.W.; Won, E.J.; Lee, Y.M. Cloning and molecular characterization of estrogen-related receptor (ERR) and vitellogenin genes in the brackish water flea Diaphanosoma celebensis exposed to bisphenol A and its structural analogues. Mar. Pollut. Bull. 2020, 154, 111063. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Wu, J.; Han, C.; Liu, H.; Xu, Y.; Zhang, H.; Lin, Y.; Xia, Q. Oestrogen-related receptor reduces vitellogenin expression by crosstalk with the ecdysone receptor pathway in female silkworm, Bombyx mori. Insect Mol. Biol. 2018, 27, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, E.V.; Mazina, M.Y.; Krasnov, A.N.; Vorobyeva, N.E. The Drosophila nuclear receptors EcR and ERR jointly regulate the expression of genes involved in carbohydrate metabolism. Insect Biochem. Mol. 2019, 112, 103184. [Google Scholar] [CrossRef]
- Liu, T.; Li, Y.; Zhao, X.; Zhang, M.; Gu, W. Ethylparaben affects lifespan, fecundity, and the expression levels of ERR, EcR and YPR in Drosophila melanogaster. J. Insect Physiol. 2014, 71, 1–7. [Google Scholar] [CrossRef]
- Mazina, M.Y.; Kocheryzhkina, E.V.; Nikolenko, J.V.; Krasnov, A.N.; Georgieva, S.G.; Vorobyeva, N.E. Nuclear receptors EcR, Usp, E75, DHR3, and ERR regulate transcription of ecdysone cascade genes. Dokl. Biochem. Biophys. 2017, 473, 145–147. [Google Scholar] [CrossRef]
- Xi, G.S.; Ying, Q.Q.; Xie, H. The functional relevance of oestrogen-related receptor gene to ultraspiracle gene in regulating the development of Polyrhachis vicina (Hymenoptera: Formicidae). Orient. Insects 2019, 53, 362–378. [Google Scholar] [CrossRef]
- Jin, W.J.; Jia, Y.S.; Tan, E.; Xi, G.S. Relevance of estrogen-related receptor gene and ecdysone receptor gene in adult testis of the cricket Teleogryllus emma (Orthoptera: Gryllidae). Sci. Nat. 2017, 104, 1–7. [Google Scholar] [CrossRef]
- He, H.; Xi, G.; Lu, X. Molecular cloning, characterization, and expression analysis of an estrogen receptor-related receptor homologue in the cricket, Teleogryllus emma. J. Insect Sci. 2010, 10, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, W.; Wu, J.; Shen, G.; Zhang, H.; Liu, H.; Xu, Y.; Gu, J.; Jia, L.; Lin, Y.; Xia, Q. Estrogen-related receptor participates in regulating glycolysis and influences embryonic development in silkworm Bombyx mori. Insect Mol. Biol. 2020, 29, 160–169. [Google Scholar] [CrossRef]
- Li, Y.; Padmanabha, D.; Gentile, L.B.; Dumur, C.I.; Beckstead, R.B.; Baker, K.D. HIF- and non-HIF-regulated hypoxic responses require the estrogen-related receptor in Drosophila melanogaster. PLoS Genet. 2013, 9, e1003230. [Google Scholar] [CrossRef]
- Beebe, K.; Robins, M.M.; Hernandez, E.J.; Lam, G.; Horner, M.A.; Thummel, C.S. Drosophila estrogen-related receptor directs a transcriptional switch that supports adult glycolysis and lipogenesis. Genes Dev. 2020, 34, 701–714. [Google Scholar] [CrossRef]
- Park, W.R.; Lim, D.J.; Sang, H.; Kim, E.; Moon, J.H.; Choi, H.S.; Kim, I.S.; Kim, D.K. Aphid estrogen-related receptor controls glycolytic gene expression and fecundity. Insect Biochem. Mol. Biol. 2021, 130, 103529. [Google Scholar] [CrossRef]
- Suetsugu, Y.; Futahashi, R.; Kanamori, H.; Kadono-Okuda, K.; Sasanuma, S.; Narukawa, J.; Ajimura, M.; Jouraku, A.; Namiki, N.; Shimomura, M.; et al. Large Scale Full-Length cDNA Sequencing Reveals a Unique Genomic Landscape in a Lepidopteran Model Insect, Bombyx mori. G3-Genes Genom Genet 2013, 3, 1481–1492. [Google Scholar] [CrossRef] [Green Version]
- Xia, Q.; Zhou, Z.; Lu, C.; Cheng, D.; Dai, F.; Li, B.; Zhao, P.; Zha, X.; Cheng, T.; Chai, C.; et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar]
- Xia, Q.Y.; Li, S.; Feng, Q.L. Advances in Silkworm Studies Accelerated by the Genome Sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef] [PubMed]
- Nwibo, D.D.; Hamamoto, H.; Matsumoto, Y.; Kaito, C.; Sekimizu, K. Current use of silkworm larvae (Bombyx mori) as an animal model in pharmaco-medical research. Drug Discov. Ther. 2015, 9, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Zhu, F.; Chen, K. Silkworm: A Promising Model Organism in Life Science. J. Insect Sci. 2017, 17, 97. [Google Scholar]
- Xia, Q.Y.; Wang, J.; Zhou, Z.Y.; Li, R.Q.; Fan, W.; Cheng, D.J.; Cheng, T.C.; Qin, J.J.; Duan, J.; Xu, H.F.; et al. The genome of a lepidopteran model insect, the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1036–1045. [Google Scholar]
- Wang, X.; Li, Y.; Xie, K.; Yi, Q.Y.; Chen, Q.M.; Wang, X.H.; Shen, H.; Xia, Q.Y.; Zhao, P. Ca2+ and endoplasmic reticulum Ca2+-ATPase regulate the formation of silk fibers with favorable mechanical properties. J. Insect Physiol. 2015, 73, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Liu, Q.; Xia, Q.; Zhao, P. Proteome profile of spinneret from the silkworm, Bombyx mori. Proteomics 2017, 17, 1600301. [Google Scholar] [CrossRef]
- Egorova, T.A.; Smolin, A.N. Study of Trehalose Metabolism in Organism of Oak Silkworm. Biochem. Mosc. 1962, 27, 696. [Google Scholar]
- Egorova, T.A.; Smolin, A.N. Trehalose in Organism of Oak Silkworm in Different Stages of Its Development. Biochem. Mosc. 1962, 27, 407. [Google Scholar]
- Thompson, S.N. Trehalose—The insect ‘blood’ sugar. Adv. Insect Physiol. 2003, 31, 205–285. [Google Scholar]
- Horie, Y. Blood Trehalose and Fat-Body Glycogen in the Silkworm, Bombyx-Mori. Nature 1960, 188, 583–584. [Google Scholar] [CrossRef] [PubMed]
- Mitsumasu, K.; Azuma, M.; Niimi, T.; Yamashita, O.; Yaginuma, T. Membrane-penetrating trehalase from silkworm Bombyx mori. Molecular cloning and localization in larval midgut. Insect Mol. Biol. 2005, 14, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Saito, S. Trehalose in the Body Fluid of the Silkworm, Bombyx-Mori L. J. Insect Physiol. 1963, 9, 509–519. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, G.; Wu, J.; Lin, Y.; Hua, X.; Xia, Q.; Zhao, P. Estrogen-Related Receptor Influences the Hemolymph Glucose Content by Regulating Midgut Trehalase Gene Expression in the Last Instar Larvae of Bombyx mori. Int. J. Mol. Sci. 2021, 22, 4343. https://doi.org/10.3390/ijms22094343
Shen G, Wu J, Lin Y, Hua X, Xia Q, Zhao P. Estrogen-Related Receptor Influences the Hemolymph Glucose Content by Regulating Midgut Trehalase Gene Expression in the Last Instar Larvae of Bombyx mori. International Journal of Molecular Sciences. 2021; 22(9):4343. https://doi.org/10.3390/ijms22094343
Chicago/Turabian StyleShen, Guanwang, Jinxin Wu, Ying Lin, Xiaoting Hua, Qingyou Xia, and Ping Zhao. 2021. "Estrogen-Related Receptor Influences the Hemolymph Glucose Content by Regulating Midgut Trehalase Gene Expression in the Last Instar Larvae of Bombyx mori" International Journal of Molecular Sciences 22, no. 9: 4343. https://doi.org/10.3390/ijms22094343
APA StyleShen, G., Wu, J., Lin, Y., Hua, X., Xia, Q., & Zhao, P. (2021). Estrogen-Related Receptor Influences the Hemolymph Glucose Content by Regulating Midgut Trehalase Gene Expression in the Last Instar Larvae of Bombyx mori. International Journal of Molecular Sciences, 22(9), 4343. https://doi.org/10.3390/ijms22094343