
Insights into the Pathophysiology of Psychiatric Symptoms in Central Nervous System Disorders: Implications for Early and Differential Diagnosis

 , , , ,
, , , , 

{kind=link}
Abstract
:1. Introduction
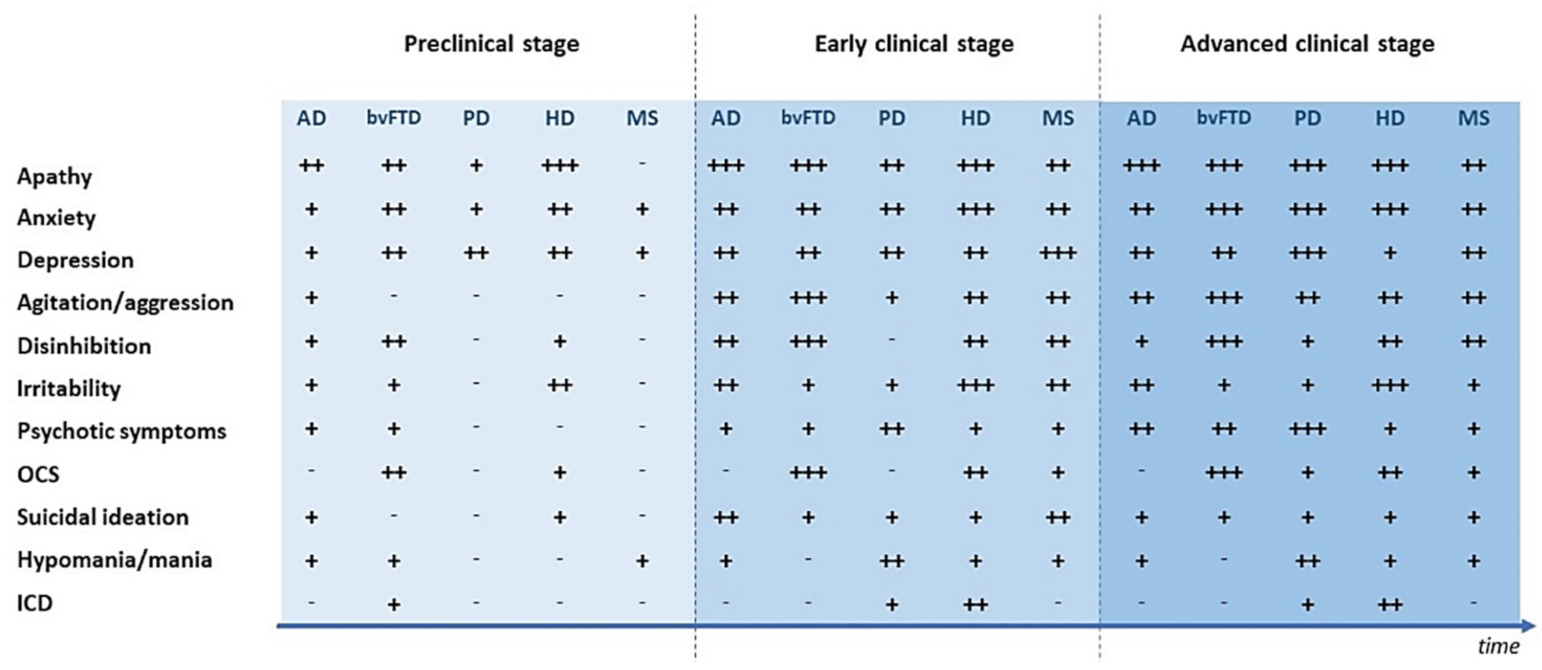
2. Psychiatric Symptoms in the Clinical Course of Neurodegenerative and Neuroinflammatory Diseases
2.1. Psychiatric Symptoms in the Preclinical Phase of Neurological Diseases
2.2. Psychiatric Symptoms in the Early Clinical Stages of Neurological Diseases
2.3. Psychiatric Symptoms in the Advanced Clinical Stages of Neurological Diseases
3. Brain Circuits Involved in Psychiatric Symptoms Occurring in CNS Disorders
3.1. Alzheimer’s Disease
3.2. Behavioral Variant of Fronto-Temporal Dementia
3.3. Parkinson’s Disease
3.4. Huntington’s Disease
3.5. Multiple Sclerosis
3.6. Future Perspectives
4. Molecular and Electrophysiological Correlates of Synaptic Dysfunction Underlying Psychiatric Symptoms in CNS Disorders
4.1. Synaptic Dysfunctions in CNS Disorders
4.2. Synaptic Dysfunctions and Psychiatric Disorders
4.3. Synaptic Dysfunctions, Neuroinflammation, and Psychiatric Disorders
4.4. Future Perspectives
5. Body Fluids Biomarkers and Psychiatric Symptoms in Neurodegenerative and Neuroinflammatory Diseases
5.1. Biomarkers of Amyloidosis and Tauopathy
5.2. Biomarkers of Ubiquitinopathy
5.3. Biomarkers of Synucleinopathy
5.4. Biomarkers of Neuroinflammation
5.5. Neurotransmitter Biomarkers
5.6. Biomarkers of Axonal Damage in the Early Detection of CNS Disorders in Patients with Psychiatric Symptoms
5.7. Future Perspectives on Synaptic, Glial and Neurotransmitter Biomarkers
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hesdorffer, D.C. Comorbidity between neurological illness and psychiatric disorders. CNS Spectr. 2016, 21, 230–238. [Google Scholar] [CrossRef]
- Sparaco, M.; Lavorgna, L.; Bonavita, S. Psychiatric disorders in multiple sclerosis. J. Neurol. 2019, 268, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Starkstein, S.E.; Jorge, R.; Mizrahi, R.; Robinson, R.G. The construct of minor and major depression in Alzheimer’s disease. Am. J. Psychiatry 2005, 162, 2086–2093. [Google Scholar] [CrossRef] [PubMed]
- Marrie, R.A.; Reingold, S.; Cohen, J.; Stuve, O.; Trojano, M.; Sorensen, P.S.; Cutter, G.; Reider, N. The incidence and prevalence of psychiatric disorders in multiple sclerosis: A systematic review. Mult. Scler. J. 2015, 21, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Paoli, R.A.; Botturi, A.; Ciammola, A.; Silani, V.; Prunas, C.; Lucchiari, C.; Zugno, E.; Caletti, E. Neuropsychiatric burden in Huntington’s disease. Brain Sci. 2017, 7, 67. [Google Scholar] [CrossRef]
- Galts, C.P.C.; Bettio, L.E.B.; Jewett, D.C.; Yang, C.C.; Brocardo, P.S.; Rodrigues, A.L.S.; Thacker, J.S.; Gil-Mohapel, J. Depression in neurodegenerative diseases: Common mechanisms and current treatment options. Neurosci. Biobehav. Rev. 2019, 102, 56–84. [Google Scholar] [CrossRef] [PubMed]
- Orr, J.; Bernstein, C.N.; Graff, L.A.; Patten, S.B.; Bolton, J.M.; Sareen, J.; Marriott, J.J.; Fisk, J.D.; Marrie, R.A. Factors associated with perceived need for mental health care in multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 25, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.F.; Tan, L.; Wang, H.F.; Jiang, T.; Tan, M.S.; Tan, L.; Xu, W.; Li, J.Q.; Wang, J.; Lai, T.J.; et al. The prevalence of neuropsychiatric symptoms in Alzheimer’s disease: Systematic review and meta-analysis. J. Affect. Disord. 2016, 190, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Husain, M. Transdiagnostic neurology: Neuropsychiatric symptoms in neurodegenerative diseases. Brain 2017, 140, 1535–1536. [Google Scholar] [CrossRef]
- Rod, N.H.; Bordelon, Y.; Thompson, A.; Marcotte, E.; Ritz, B. Major life events and development of major depression in Parkinson’s disease patients. Eur. J. Neurol. 2013, 20, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Solaro, C.; Gamberini, G.; Masuccio, F.G. Depression in Multiple Sclerosis: Epidemiology, Aetiology, Diagnosis and Treatment. CNS Drugs 2018, 32, 117–133. [Google Scholar] [CrossRef]
- Linnemann, C.; Lang, U.E. Pathways Connecting Late-Life Depression and Dementia. Front. Pharmacol. 2020, 11, 279. [Google Scholar] [CrossRef]
- Lorefice, L.; Fenu, G.; Fois, M.; Frau, J.; Coghe, G.; Marrosu, M.G.; Cocco, E. Pulse steroid therapy in multiple sclerosis and mood changes: An exploratory prospective study. Mult. Scler. Relat. Disord. 2018, 20, 104–108. [Google Scholar] [CrossRef]
- Weintraub, D.; Papay, K.; Siderowf, A. Screening for impulse control symptoms in patients with de novo Parkinson disease: A casecontrol study. Neurology 2013, 80, 176–180. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Kumar, P.; Khan, S.; Gordon, D.K.; Khan, S. Similarities Between Depression and Neurodegenerative Diseases: Pathophysiology, Challenges in Diagnosis and Treatment Options. Cureus 2020, 12, e11613. [Google Scholar] [CrossRef]
- Szatmari, S.; Ajtay, A.; Oberfrank, F.; Dobi, B.; Bereczki, D. The prevalence of psychiatric symptoms before the diagnosis of Parkinson’s disease in a nationwide cohort: A comparison to patients with cerebral infarction. PLoS ONE 2020, 15, e0236728. [Google Scholar] [CrossRef]
- Lleó, A.; Parnetti, L.; Belbin, O.; Wiltfang, J. Has the time arrived for cerebrospinal fluid biomarkers in psychiatric disorders? Clin. Chim. Acta 2019, 491, 81–84. [Google Scholar] [CrossRef]
- Gaetani, L.; Paolini Paoletti, F.; Bellomo, G.; Mancini, A.; Simoni, S.; Di Filippo, M.; Parnetti, L. CSF and Blood Biomarkers in Neuroinflammatory and Neurodegenerative Diseases: Implications for Treatment. Trends Pharmacol. Sci. 2020, 41, 1023–1037. [Google Scholar] [CrossRef]
- Teunissen, C.E.; Otto, M.; Engelborghs, S.; Herukka, S.K.; Lehmann, S.; Lewczuk, P.; Lleó, A.; Perret-Liaudet, A.; Tumani, H.; Turner, M.R.; et al. White paper by the Society for CSF Analysis and Clinical Neurochemistry: Overcoming barriers in biomarker development and clinical translation. Alzheimer’s Res. Ther. 2018, 10, 30. [Google Scholar] [CrossRef]
- Lewczuk, P.; Riederer, P.; O’Bryant, S.E.; Verbeek, M.M.; Dubois, B.; Visser, P.J.; Jellinger, K.A.; Engelborghs, S.; Ramirez, A.; Parnetti, L.; et al. Cerebrospinal fluid and blood biomarkers for neurodegenerative dementias: An update of the Consensus of the Task Force on Biological Markers in Psychiatry of the World Federation of Societies of Biological Psychiatry. World J. Biol. Psychiatry 2018, 19, 244–328. [Google Scholar] [CrossRef]
- Trojano, L.; Papagno, C. Cognitive and behavioral disorders in Parkinson’s disease: An update. II: Behavioral disorders. Neurol. Sci. 2018, 39, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Silveira, C.; Guedes, R.; Maia, D.; Curral, R.; Coelho, R. Neuropsychiatric symptoms of multiple sclerosis: State of the art. Psychiatry Investig. 2019, 16, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Escudero, J.M.S.; Beltrán, J.; Palacios, Á.; Chimbí, C.M.; Matallana, D.; Reyes, P.; Perez-Sola, V.; Santamaría-García, H. Neuropsychiatric symptoms as predictors of clinical course in neurodegeneration. A longitudinal study. Front. Aging Neurosci. 2019, 11, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loi, S.M.; Walterfang, M.; Velakoulis, D.; Looi, J.C.L. Huntington’s disease: Managing neuropsychiatric symptoms in Huntington’s disease. Australas. Psychiatry 2018, 26, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, K.; Sgambato, V. Neuropsychiatric Disorders in Parkinson’s Disease: What Do We Know About the Role of Dopaminergic and Non-dopaminergic Systems? Front. Neurosci. 2020, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Dubovsky, S.L. Mania. Contin. Lifelong Learn. Neurol. 2015, 21, 737–755. [Google Scholar] [CrossRef]
- Rothenberg, K.G.; Rajaram, R. Advances in Management of Psychosis in Neurodegenerative Diseases. Curr. Treat. Options Neurol. 2019, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- Masters, M.C.; Morris, J.C.; Roe, C.M. “Noncognitive” symptoms of early Alzheimer disease: A longitudinal analysis. Neurology 2015, 84, 617–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poletti, M.; Bonuccelli, U. Psychopathological disorders in Parkinson’s disease. J. Psychopathol. 2011, 17, 13–21. [Google Scholar]
- Patten, S.B.; Marrie, R.A.; Carta, M.G. Depression in multiple sclerosis. Int. Rev. Psychiatry 2017, 29, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Wise, E.A.; Rosenberg, P.B.; Lyketsos, C.G.; Leoutsakos, J.M. Time course of neuropsychiatric symptoms and cognitive diagnosis in National Alzheimer’s Coordinating Centers volunteers. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2019, 11, 333–339. [Google Scholar] [CrossRef]
- Benussi, A.; Premi, E.; Gazzina, S.; Brattini, C.; Bonomi, E.; Alberici, A.; Jiskoot, L.; van Swieten, J.C.; Sanchez-Valle, R.; Moreno, F.; et al. Progression of Behavioral Disturbances and Neuropsychiatric Symptoms in Patients with Genetic Frontotemporal Dementia. JAMA Netw. Open 2021, 4, e2030194. [Google Scholar] [CrossRef] [PubMed]
- Ismail, Z.; Gatchel, J.; Bateman, D.R.; Barcelos-Ferreira, R.; Chantillon, M.; Jaeger, J.; Donovan, N.J.; Mortby, M.E. Affective and emotional dysregulation as pre-dementia risk markers: Exploring the mild behavioral impairment symptoms of depression, anxiety, irritability, and euphoria. Int. Psychogeriatr. 2018, 30, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Seritan, A.L.; Rienas, C.; Duong, T.; Delucchi, K.; Ostrem, J.L. Ages at onset of anxiety and depressive disorders in parkinson’s disease. J. Neuropsychiatry Clin. Neurosci. 2019, 31, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.S.; Huey, E.D.; Thorne, D.Z. The Confluence of Psychiatric Symptoms and Neurodegenerative Disease: Impact on Genetic Counseling. J. Genet. Couns. 2017, 26, 435–441. [Google Scholar] [CrossRef]
- Heinzel, S.; Berg, D.; Gasser, T.; Chen, H.; Yao, C.; Postuma, R.B. Update of the MDS research criteria for prodromal Parkinson’s disease. Mov. Disord. 2019, 34, 1464–1470. [Google Scholar] [CrossRef]
- Giagkou, N.; Maraki, M.I.; Yannakoulia, M.; Kosmidis, M.H.; Dardiotis, E.; Hadjigeorgiou, G.M.; Sakka, P.; Ntanasi, E.; Anastasiou, C.A.; Xiromerisiou, G.; et al. A Prospective Validation of the Updated Movement Disorders Society Research Criteria for Prodromal Parkinson’s Disease. Mov. Disord. 2020, 35, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Woolley, J.D.; Khan, B.K.; Murthy, N.K.; Miller, B.L.; Rankin, K.P. The diagnostic challenge of psychiatric symptoms in neurodegenerative disease: Rates of and risk factors for prior psychiatric diagnosis in patients with early neurodegenerative disease. J. Clin. Psychiatry 2011, 72, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, H.; Ganesh, A.; Lau, R.; Lysack, J.; Ismail, Z. Cholinesterase-inhibitor associated mania: A case report and literature review. Can. J. Neurol. Sci. 2014, 41, 278–280. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.; O’Donoghue, S.; Counihan, T.; McDonald, C.; Calabresi, P.A.; Ahmed, M.A.S.; Kaplin, A.; Hallahan, B. Neuropsychiatric syndromes of multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2017, 88, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, F.L.A.; Ng, B.C.; Wijnands, J.M.A.; Kingwell, E.; Marrie, R.A.; Tremlett, H. A systematic review of morbidities suggestive of the multiple sclerosis prodrome. Expert Rev. Neurother. 2020, 20, 799–819. [Google Scholar] [CrossRef]
- Lo Fermo, S.; Barone, R.; Patti, F.; Laisa, P.; Cavallaro, T.L.; Nicoletti, A.; Zappia, M. Outcome of psychiatric symptoms presenting at onset of multiple sclerosis: A retrospective study. Mult. Scler. 2010, 16, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Wijnands, J.M.A.; Zhu, F.; Kingwell, E.; Zhao, Y.; Ekuma, O.; Lu, X.; Evans, C.; Fisk, J.D.; Marrie, R.A.; Tremlett, H. Five years before multiple sclerosis onset: Phenotyping the prodrome. Mult. Scler. J. 2019, 25, 1092–1101. [Google Scholar] [CrossRef]
- Hoang, H.; Laursen, B.; Stenager, E.N.; Stenager, E. Psychiatric co-morbidity in multiple sclerosis: The risk of depression and anxiety before and after MS diagnosis. Mult. Scler. J. 2016, 22, 347–353. [Google Scholar] [CrossRef]
- Disanto, G.; Zecca, C.; MacLachlan, S.; Sacco, R.; Handunnetthi, L.; Meier, U.C.; Simpson, A.; McDonald, L.; Rossi, A.; Benkert, P.; et al. Prodromal symptoms of multiple sclerosis in primary care. Ann. Neurol. 2018, 83, 1162–1173. [Google Scholar] [CrossRef]
- Carta, M.G.; Moro, M.F.; Lorefice, L.; Picardi, A.; Trincas, G.; Fenu, G.; Cocco, E.; Floris, F.; Bessonov, D.; Akiskal, H.S.; et al. Multiple sclerosis and bipolar disorders: The burden of comorbidity and its consequences on quality of life. J. Affect. Disord. 2014, 167, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Cortés, N.; Andrade, V.; Maccioni, R.B. Behavioral and Neuropsychiatric Disorders in Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 899–910. [Google Scholar] [CrossRef] [Green Version]
- Vik-Mo, A.O.; Giil, L.M.; Borda, M.G.; Ballard, C.; Aarsland, D. The individual course of neuropsychiatric symptoms in people with Alzheimer’s and Lewy body dementia: 12-year longitudinal cohort study. Br. J. Psychiatry 2020, 216, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Marin, R.S. Apathy: A neuropsychiatric syndrome. J. Neuropsychiatry Clin. Neurosci. 1991, 3, 243–254. [Google Scholar] [CrossRef]
- Boyle, P.A.; Malloy, P.F. Treating apathy in Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2004, 17, 91–99. [Google Scholar] [CrossRef]
- Buettner, L.; Fitzsimmons, S.; Atav, S.; Sink, K. Cognitive stimulation for apathy in probable early-stage Alzheimer’s. J. Aging Res. 2011, 2011, 480890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumfor, F.; Zhen, A.; Hodges, J.R.; Piguet, O.; Irish, M. Apathy in Alzheimer’s disease and frontotemporal dementia: Distinct clinical profiles and neural correlates. Cortex 2018, 103, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Irish, M.; Hodges, J.R.; Piguet, O.; Kumfor, F. Disease-specific profiles of apathy in Alzheimer’s disease and behavioural-variant frontotemporal dementia differ across the disease course. J. Neurol. 2020, 267, 1086–1096. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.D.; Henley, S.M.D.; Suárez-González, A. A systematic review of the prevalence of depression, anxiety, and apathy in frontotemporal dementia, atypical and young-onset Alzheimer’s disease, and inherited dementia. Int. Psychogeriatr. 2020, 20, 1–20. [Google Scholar] [CrossRef]
- Radakovic, R.; Colville, S.; Cranley, D.; Starr, J.M.; Pal, S.; Abrahams, S. Multidimensional Apathy in Behavioral Variant Frontotemporal Dementia, Primary Progressive Aphasia, and Alzheimer Disease. J. Geriatr. Psychiatry Neurol. 2020. [Google Scholar] [CrossRef]
- Radakovic, R.; Davenport, R.; Starr, J.M.; Abrahams, S. Apathy dimensions in Parkinson’s disease. Int. J. Geriatr. Psychiatry 2018, 33, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, G.; Vitale, C.; Trojano, L.; Picillo, M.; Moccia, M.; Pisano, G.; Pezzella, D.; Cuoco, S.; Erro, R.; Longo, K.; et al. Relationship between apathy and cognitive dysfunctions in de novo untreated Parkinson’s disease: A prospective longitudinal study. Eur. J. Neurol. 2015, 22, 253–260. [Google Scholar] [CrossRef]
- Pagonabarraga, J.; Kulisevsky, J.; Strafella, A.P.; Krack, P. Apathy in Parkinson’s disease: Clinical features, neural substrates, diagnosis, and treatment. Lancet Neurol. 2015, 14, 518–531. [Google Scholar] [CrossRef] [Green Version]
- Vergallo, A.; Giampietri, L.; Pagni, C.; Giorgi, F.S.; Nicoletti, V.; Miccoli, M.; Libertini, P.; Petrozzi, L.; Bonuccelli, U.; Tognoni, G. Association Between CSF Beta-Amyloid and Apathy in Early-Stage Alzheimer Disease. J. Geriatr. Psychiatry Neurol. 2019, 32, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Shail, M.S. Neuropsychiatry in Demyelination Disease: Using Depression as a Prodrome for Early Diagnosis and Treatment of Multiple Sclerosis. Cureus 2017, 9, e1813. [Google Scholar] [CrossRef] [Green Version]
- Possa, M.F.; Minacapelli, E.; Canale, S.; Comi, G.; Martinelli, V.; Falautano, M. The first year after diagnosis: Psychological impact on people with multiple sclerosis. Psychol. Health Med. 2017, 22, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Grochmal-Bach, B.; Bidzan, L.; Pachalska, M.; Bidzan, M.; Łukaszewska, B.; Pufal, A. Aggressive and impulsive behaviors in Frontotemporal dementia and Alzheimer’s disease. Med. Sci. Monit. 2009, 15, CR248-54. [Google Scholar] [PubMed]
- Kumar, A.; Tsao, J.W. Alzheimer Disease: REVUE; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Perry, D.C.; Whitwell, J.L.; Boeve, B.F.; Pankratz, V.S.; Knopman, D.S.; Petersen, R.C.; Jack, C.R.; Josephs, K.A. Voxel-based morphometry in patients with obsessive-compulsive behaviors in behavioral variant frontotemporal dementia. Eur. J. Neurol. 2012, 19, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, M.A.; Shnall, A.; Onyike, C.U.; Huey, E.D. Management of frontotemporal dementia in mental health and multidisciplinary settings. Int. Rev. Psychiatry 2013, 25, 230–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Riva, P.; Smith, K.; Xie, S.X.; Weintraub, D. Course of psychiatric symptoms and global cognition in early Parkinson disease. Neurology 2014, 83, 1096–1103. [Google Scholar] [CrossRef]
- Ecker, D.; Unrath, A.; Kassubek, J.; Sabolek, M. Dopamine Agonists and their risk to induce psychotic episodes in Parkinson’s disease: A case-control study. BMC Neurol. 2009, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weintraub, D.; David, A.S.; Evans, A.H.; Grant, J.E.; Stacy, M. Clinical spectrum of impulse control disorders in Parkinson’s disease. Mov. Disord. 2015, 30, 121–127. [Google Scholar] [CrossRef]
- Van Duijn, E.; Craufurd, D.; Hubers, A.A.M.; Giltay, E.J.; Bonelli, R.; Rickards, H.; Anderson, K.E.; Van Walsem, M.R.; Van Der Mast, R.C.; Orth, M.; et al. Neuropsychiatric symptoms in a European Huntington’s disease cohort (REGISTRY). J. Neurol. Neurosurg. Psychiatry 2014, 85, 1411–1418. [Google Scholar] [CrossRef] [Green Version]
- Hartoonian, N.; Terrill, A.L.; Beier, M.L.; Turner, A.P.; Day, M.A.; Alschuler, K.N. Predictors of anxiety in multiple sclerosis. Rehabil. Psychol. 2015, 60, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.M.W.; Salem, R.; Amtmann, D. Somatic symptoms of depression and anxiety in people with multiple sclerosis. Int. J. MS Care 2018, 20, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Thériault, R.K.; Manduca, J.D.; Perreault, M.L. Sex differences in innate and adaptive neural oscillatory patterns predict resilience and susceptibility to chronic stress in rats. bioRxiv 2021, 46, E258–E270. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Palleiro, P.; Rivera-Baltanás, T.; Rodrigues-Amorim, D.; Fernández-Gil, S.; del Carmen Vallejo-Curto, M.; Álvarez-Ariza, M.; López, M.; Rodriguez-Jamardo, C.; Luis Benavente, J.; de las Heras, E.; et al. Brainwaves Oscillations as a Potential Biomarker for Major Depression Disorder Risk. Clin. EEG Neurosci. 2020, 51, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Deshpande, G.; Zhao, S.; Liu, J.; Hu, X.; Li, K. Altered directional connectivity between emotion network and motor network in Parkinson’s disease with depression. Medicine 2016, 95, e4222. [Google Scholar] [CrossRef] [PubMed]
- Lucas-Jiménez, O.; Ojeda, N.; Peña, J.; Cabrera-Zubizarreta, A.; Díez-Cirarda, M.; Gómez-Esteban, J.C.; Gómez-Beldarrain, M.Á.; Ibarretxe-Bilbao, N. Apathy and brain alterations in Parkinson’s disease: A multimodal imaging study. Ann. Clin. Transl. Neurol. 2018, 5, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Manduca, J.D.; Thériault, R.K.; Williams, O.O.F.; Rasmussen, D.J.; Perreault, M.L. Transient Dose-dependent Effects of Ketamine on Neural Oscillatory Activity in Wistar-Kyoto Rats. Neuroscience 2020, 441, 161–175. [Google Scholar] [CrossRef]
- Babiloni, C.; Pascarelli, M.T.; Lizio, R.; Noce, G.; Lopez, S.; Rizzo, M.; Ferri, R.; Soricelli, A.; Nobili, F.; Arnaldi, D.; et al. Abnormal cortical neural synchronization mechanisms in quiet wakefulness are related to motor deficits, cognitive symptoms, and visual hallucinations in Parkinson’s disease patients: An electroencephalographic study. Neurobiol. Aging 2020, 91, 88–111. [Google Scholar] [CrossRef]
- Thériault, R.K.; Perreault, M.L. Hormonal regulation of circuit function: Sex, systems and depression. Biol. Sex Differ. 2019, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Dauwan, M.; Hoff, J.I.; Vriens, E.M.; Hillebrand, A.; Stam, C.J.; Sommer, I.E. Aberrant resting-state oscillatory brain activity in Parkinson’s disease patients with visual hallucinations: An MEG source-space study. NeuroImage Clin. 2019, 22. [Google Scholar] [CrossRef]
- Başar, E.; Femir, B.; Emek-Savaş, D.D.; Güntekin, B.; Yener, G.G. Increased long distance event-related gamma band connectivity in Alzheimer’s disease. NeuroImage Clin. 2017, 14, 580–590. [Google Scholar] [CrossRef]
- Ilhan Atagün, M.; Güntekin, B.; Tan, D.; Elif Tülay, E.; Başar, E. Lithium excessively enhances event related beta oscillations in patients with bipolar disorder. J. Affect. Disord. 2015, 170, 59–65. [Google Scholar] [CrossRef]
- Rosenberg, P.B.; Nowrangi, M.A.; Lyketsos, C.G. Neuropsychiatric symptoms in Alzheimer’s disease: What might be associated brain circuits? Mol. Asp. Med. 2015, 43–44, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Büyükgök, D.; Bayraktaroğlu, Z.; Seda Buker, H.; Işın Baral Kulaksızoğlu, M.; Hakan, G. Resting-state FMRI analysis in apathetic Alzheimer’s disease. Diagn. Interv. Radiol. 2020, 26, 363–369. [Google Scholar] [CrossRef]
- Dam, D.V.; Vermeiren, Y.; Dekker, A.D.; Naudé, P.J.W.; De Deyn, P.P. Neuropsychiatric Disturbances in Alzheimer’s Disease: What Have We Learned from Neuropathological Studies? Curr. Alzheimer Res. 2016, 13, 1145–1164. [Google Scholar] [CrossRef] [PubMed]
- Geda, Y.E.; Schneider, L.S.; Gitlin, L.N.; Miller, D.S.; Smith, G.S.; Bell, J.; Evans, J.; Lee, M.; Porsteinsson, A.; Lanctôt, K.L.; et al. Neuropsychiatric symptoms in Alzheimer’s disease: Past progress and anticipation of the future. Alzheimer’s Dement. 2013, 9, 602–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grupe, D.W.; Nitschke, J.B. Uncertainty and anticipation in anxiety: An integrated neurobiological and psychological perspective. Nat. Rev. Neurosci. 2013, 14, 488–501. [Google Scholar] [CrossRef]
- Apostolova, L.G.; Akopyan, G.G.; Partiali, N.; Steiner, C.A.; Dutton, R.A.; Hayashi, K.M.; Dinov, I.D.; Toga, A.W.; Cummings, J.L.; Thompson, P.M. Structural correlates of apathy in Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2007, 24, 91–97. [Google Scholar] [CrossRef]
- Stanton, B.R.; Leigh, P.N.; Howard, R.J.; Barker, G.J.; Brown, R.G. Behavioural and emotional symptoms of apathy are associated with distinct patterns of brain atrophy in neurodegenerative disorders. J. Neurol. 2013, 260, 2481–2490. [Google Scholar] [CrossRef]
- Starkstein, S.E.; Mizrahi, R.; Capizzano, A.A.; Acion, L.; Brockman, S.; Power, B.D. Neuroimaging Correlates of Apathy and Depression in Alzheimer’s Disease. J. Neuropsychiatr. 2009, 21, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Roberto, A.J.; Mohan, A.; Lorenzo, A.; Jones, K.; Carney, M.J.; Liogier-Weyback, L.; Hwang, S.; Lapidus, K.A.B. The significance of the Default Mode Network (DMN) in neurological and neuropsychiatric disorders: A review. Yale J. Biol. Med. 2016, 89, 49–57. [Google Scholar]
- Spreng, R.N.; Mar, R.A.; Kim, A.S.N. The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: A quantitative meta-analysis. J. Cogn. Neurosci. 2009, 21, 489–510. [Google Scholar] [CrossRef]
- Royall, D.R.; Palmer, R.F.; Vidoni, E.D.; Honea, R.A. The default mode network may be the key substrate of depressive symptom-related cognitive changes. J. Alzheimer’s Dis. 2013, 34, 547–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, C.E.; Donovan, N.J.; Guercio, B.J.; Wigman, S.E.; Schultz, A.P.; Amariglio, R.E.; Rentz, D.M.; Johnson, K.A.; Sperling, R.A.; Marshall, G.A. Neuropsychiatric symptoms and functional connectivity in mild cognitive impairment. J. Alzheimer’s Dis. 2015, 46, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Song, X.; Li, E.; Liu, J.; Yuan, Y.; Liu, W.; Liu, Y. Altered resting-state brain activity and connectivity in depressed Parkinson’s disease. PLoS ONE 2015, 10, e0131133. [Google Scholar] [CrossRef]
- Zahodne, L.B.; Gongvatana, A.; Cohen, R.A.; Ott, B.R.; Tremont, G. Are apathy and depression independently associated with longitudinal trajectories of cortical atrophy in mild cognitive impairment? Am. J. Geriatr. Psychiatry 2013, 21, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.J.; Lu, P.H.; Hua, X.; Lee, S.; Wu, S.; Nguyen, K.; Teng, E.; Leow, A.D.; Jack, C.R.; Toga, A.W.; et al. Depressive symptoms in mild cognitive impairment predict greater atrophy in alzheimer’s disease-related regions. Biol. Psychiatry 2012, 71, 814–821. [Google Scholar] [CrossRef] [Green Version]
- Sacuiu, S.; Insel, P.S.; Mueller, S.; Tosun, D.; Mattsson, N.; Jack, C.R.; DeCarli, C.; Petersen, R.; Aisen, P.S.; Weiner, M.W.; et al. Chronic depressive symptomatology in mild cognitive impairment is associated with frontal atrophy rate which hastens conversion to Alzheimer dementia. Am. J. Geriatr. Psychiatry 2016, 24, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Seeley, W.W. Network dysfunction in Alzheimer’s disease and frontotemporal dementia: Implications for psychiatry. Biol. Psychiatry 2014, 75, 565–573. [Google Scholar] [CrossRef]
- Mattsson, N.; Insel, P.S.; Nosheny, R.; Tosun, D.; Trojanowski, J.Q.; Shaw, L.M.; Jack, C.R.; Donohue, M.C.; Weiner, M.W. Emerging β-amyloid pathology and accelerated cortical atrophy. JAMA Neurol. 2014, 71, 725–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeley, W.W.; Allman, J.M.; Carlin, D.A.; Crawford, R.K.; Macedo, M.N.; Greicius, M.D.; DeArmond, S.J.; Miller, B.L. Divergent social functioning in behavioral variant frontotemporal dementia and Alzheimer disease: Reciprocal networks and neuronal evolution. Alzheimer Dis. Assoc. Disord. 2007, 21, S50–S57. [Google Scholar] [CrossRef]
- Uddin, L.Q. Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 2015, 16, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Hornberger, M.; Yew, B.; Gilardoni, S.; Mioshi, E.; Gleichgerrcht, E.; Manes, F.; Hodges, J.R. Ventromedial-frontopolar prefrontal cortex atrophy correlates with insight loss in frontotemporal dementia and Alzheimer’s disease. Hum. Brain Mapp. 2014, 35, 616–626. [Google Scholar] [CrossRef]
- Irish, M.; Piguet, O.; Hodges, J.R. Self-projection and the default network in frontotemporal dementia. Nat. Rev. Neurol. 2012, 8, 152–161. [Google Scholar] [CrossRef]
- Sheelakumari, R.; Bineesh, C.; Varghese, T.; Kesavadas, C.; Verghese, J.; Mathuranath, P.S. Neuroanatomical correlates of apathy and disinhibition in behavioural variant frontotemporal dementia. Brain Imaging Behav. 2020, 14, 2004–2011. [Google Scholar] [CrossRef] [PubMed]
- Basavaraju, R.; Feng, X.; France, J.; Huey, E.D.; Provenzano, F.A. Depression Is Associated With Preserved Cortical Thickness Relative to Apathy in Frontotemporal Dementia. J. Geriatr. Psychiatry Neurol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Malpetti, M.; Jones, P.S.; Tsvetanov, K.A.; Rittman, T.; van Swieten, J.C.; Borroni, B.; Sanchez-Valle, R.; Moreno, F.; Laforce, R.; Graff, C.; et al. Apathy in presymptomatic genetic frontotemporal dementia predicts cognitive decline and is driven by structural brain changes. Alzheimer’s Dement. 2020. [Google Scholar] [CrossRef]
- Price, J.L.; Drevets, W.C. Neurocircuitry of mood disorders. Neuropsychopharmacology 2010, 35, 192–216. [Google Scholar] [CrossRef] [PubMed]
- Ehgoetz Martens, K.A.; Lewis, S.J.G. Pathology of behavior in PD: What is known and what is not? J. Neurol. Sci. 2017, 374, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Castrioto, A.; Thobois, S.; Carnicella, S.; Maillet, A.; Krack, P. Emotional manifestations of PD: Neurobiological basis. Mov. Disord. 2016, 31, 1103–1113. [Google Scholar] [CrossRef]
- Ansari, M.; Moradi, S.A.; Sherbaf, F.G.; Hedayatnia, A.; Aarabi, M.H. Comparison of structural connectivity in Parkinson’s disease with depressive symptoms versus non-depressed: A diffusion MRI connectometry study. Int. Psychogeriatr. 2019, 31, 5–12. [Google Scholar] [CrossRef]
- De Micco, R.; Satolli, S.; Siciliano, M.; Di Nardo, F.; Caiazzo, G.; Russo, A.; Giordano, A.; Esposito, F.; Tedeschi, G.; Tessitore, A. Connectivity Correlates of Anxiety Symptoms in Drug-Naive Parkinson’s Disease Patients. Mov. Disord. 2020, 36, 96–105. [Google Scholar] [CrossRef]
- Carey, G.; Lopes, R.; Viard, R.; Betrouni, N.; Kuchcinski, G.; Devignes, Q.; Defebvre, L.; Leentjens, A.F.G.; Dujardin, K. Anxiety in Parkinson’s disease is associated with changes in the brain fear circuit. Park. Relat. Disord. 2020, 80, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Thobois, S.; Prange, S.; Sgambato-Faure, V.; Tremblay, L.; Broussolle, E. Imaging the Etiology of Apathy, Anxiety, and Depression in Parkinson’s Disease: Implication for Treatment. Curr. Neurol. Neurosci. Rep. 2017, 17, 76. [Google Scholar] [CrossRef] [PubMed]
- Dujardin, K.; Sockeel, P.; Devos, D.; Delliaux, M.; Krystkowiak, P.; Destée, A.; Defebvre, L. Characteristics of apathy in Parkinson’s disease. Mov. Disord. 2007, 22, 778–784. [Google Scholar] [CrossRef]
- Sun, H.H.; Pan, P.L.; Hu, J.B.; Chen, J.; Wang, X.Y.; Liu, C.F. Alterations of regional homogeneity in Parkinson’s disease with “pure” apathy: A resting-state fMRI study. J. Affect. Disord. 2020, 274, 792–798. [Google Scholar] [CrossRef]
- Imperiale, F.; Agosta, F.; Canu, E.; Markovic, V.; Inuggi, A.; Jecmenica-Lukic, M.; Tomic, A.; Copetti, M.; Basaia, S.; Kostic, V.S.; et al. Brain structural and functional signatures of impulsive-compulsive behaviours in Parkinson’s disease. Mol. Psychiatry 2018, 23, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Carriere, N.; Lopes, R.; Defebvre, L.; Delmaire, C.; Dujardin, K. Impaired corticostriatal connectivity in impulse control disorders in Parkinson’s disease. Neurology 2015, 84, 2116–2123. [Google Scholar] [CrossRef]
- Voon, V.; Gao, J.; Brezing, C.; Symmonds, M.; Ekanayake, V.; Fernandez, H.; Dolan, R.J.; Hallett, M. Dopamine agonists and risk: Impulse control disorders in Parkinson’s Disease. Brain 2011, 134, 1438–1446. [Google Scholar] [CrossRef]
- Aylward, E.H.; Sparks, B.F.; Field, K.M.; Yallapragada, V.; Shpritz, B.D.; Rosenblatt, A.; Brandt, J.; Gourley, L.M.; Liang, K.; Zhou, H.; et al. Onset and rate of striatal atrophy in preclinical Huntington disease. Neurology 2004, 63, 66–72. [Google Scholar] [CrossRef]
- Salloway, S.; Cummings, J. Subcortical disease and neuropsychiatric illness. J. Neuropsychiatry Clin. Neurosci. 1994, 6, 93–99. [Google Scholar] [CrossRef]
- Martínez-Horta, S.; Perez-Perez, J.; Sampedro, F.; Pagonabarraga, J.; Horta-Barba, A.; Carceller-Sindreu, M.; Gomez-Anson, B.; Lozano-Martinez, G.A.; Lopez-Mora, D.A.; Camacho, V.; et al. Structural and metabolic brain correlates of apathy in Huntington’s disease. Mov. Disord. 2018, 33, 1151–1159. [Google Scholar] [CrossRef]
- Baake, V.; Coppen, E.M.; van Duijn, E.; Dumas, E.M.; van den Bogaard, S.J.A.; Scahill, R.I.; Johnson, H.; Leavitt, B.; Durr, A.; Tabrizi, S.J.; et al. Apathy and atrophy of subcortical brain structures in Huntington’s disease: A two-year follow-up study. NeuroImage Clin. 2018, 19, 66–70. [Google Scholar] [CrossRef]
- Misiura, M.B.; Ciarochi, J.; Vaidya, J.; Bockholt, J.; Johnson, H.J.; Calhoun, V.D.; Paulsen, J.S.; Turner, J.A. Apathy Is Related to Cognitive Control and Striatum Volumes in Prodromal Huntington’s Disease. J. Int. Neuropsychol. Soc. 2019, 25, 462–469. [Google Scholar] [CrossRef]
- De Paepe, A.E.; Sierpowska, J.; Garcia-Gorro, C.; Martinez-Horta, S.; Perez-Perez, J.; Kulisevsky, J.; Rodriguez-Dechicha, N.; Vaquer, I.; Subira, S.; Calopa, M.; et al. White matter cortico-striatal tracts predict apathy subtypes in Huntington’s disease. NeuroImage Clin. 2019, 24, 101965. [Google Scholar] [CrossRef] [PubMed]
- McColgan, P.; Razi, A.; Gregory, S.; Seunarine, K.K.; Durr, A.; Roos, R.A.C.; Leavitt, B.R.; Scahill, R.I.; Clark, C.A.; Langbehn, D.R.; et al. Structural and functional brain network correlates of depressive symptoms in premanifest Huntington’s disease. Hum. Brain Mapp. 2017, 38, 2819–2829. [Google Scholar] [CrossRef] [Green Version]
- Sprengelmeyer, R.; Orth, M.; Müller, H.P.; Wolf, R.C.; Grön, G.; Depping, M.S.; Kassubek, J.; Justo, D.; Rees, E.M.; Haider, S.; et al. The neuroanatomy of subthreshold depressive symptoms in Huntington’s disease: A combined diffusion tensor imaging (DTI) and voxel-based morphometry (VBM) study. Psychol. Med. 2014, 44, 1867–1878. [Google Scholar] [CrossRef]
- Pape, K.; Tamouza, R.; Leboyer, M.; Zipp, F. Immunoneuropsychiatry—Novel perspectives on brain disorders. Nat. Rev. Neurol. 2019, 15, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Rocca, M.A.; Barkhof, F.; Brück, W.; Chen, J.T.; Comi, G.; DeLuca, G.; De Stefano, N.; Erickson, B.J.; Evangelou, N.; et al. Association between pathological and MRI findings in multiple sclerosis. Lancet Neurol. 2012, 11, 349–360. [Google Scholar] [CrossRef]
- Di Filippo, M.; Portaccio, E.; Mancini, A.; Calabresi, P. Multiple sclerosis and cognition: Synaptic failure and network dysfunction. Nat. Rev. Neurosci. 2018, 19, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, M.; Magliozzi, R.; Ciccarelli, O.; Geurts, J.J.G.; Reynolds, R.; Martin, R. Exploring the origins of grey matter damage in multiple sclerosis. Nat. Rev. Neurosci. 2015, 16, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Passamonti, L.; Cerasa, A.; Liguori, M.; Gioia, M.C.; Valentino, P.; Nisticò, R.; Quattrone, A.; Fera, F. Neurobiological mechanisms underlying emotional processing in relapsing-remitting multiple sclerosis. Brain 2009, 132, 3380–3391. [Google Scholar] [CrossRef] [Green Version]
- Rocca, M.A.; Pravatà, E.; Valsasina, P.; Radaelli, M.; Colombo, B.; Vacchi, L.; Gobbi, C.; Comi, G.; Falini, A.; Filippi, M. Hippocampal-DMN disconnectivity in MS is related to WM lesions and depression. Hum. Brain Mapp. 2015, 36, 5051–5063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinstein, A.; Magalhaes, S.; Richard, J.F.; Audet, B.; Moore, C. The link between multiple sclerosis and depression. Nat. Rev. Neurol. 2014, 10, 507–517. [Google Scholar] [CrossRef]
- Feinstein, A. Neuropsychiatric syndromes associated with multiple sclerosis. J. Neurol. 2007, 254, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, A.; Roy, P.; Lobaugh, N.; Feinstein, K.; O’Connor, P.; Black, S. Structural brain abnormalities in multiple sclerosis patients with major depression. Neurology 2004, 62, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Haider, L.; Zrzavy, T.; Hametner, S.; Höftberger, R.; Bagnato, F.; Grabner, G.; Trattnig, S.; Pfeifenbring, S.; Brück, W.; Lassmann, H. The topograpy of demyelination and neurodegeneration in the multiple sclerosis brain. Brain 2016, 139, 807–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakshi, R.; Czarnecki, D.; Shaikh, Z.A.; Priore, R.L.; Janardhan, V.; Kaliszky, Z.; Kinkel, P.R. Brain MRI lesions and atrophy are related to depression in multiple sclerosis. Neuroreport 2000, 11, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Lazzarotto, A.; Margoni, M.; Franciotta, S.; Zywicki, S.; Riccardi, A.; Poggiali, D.; Anglani, M.; Gallo, P. Selective Cerebellar Atrophy Associates with Depression and Fatigue in the Early Phases of Relapse-Onset Multiple Sclerosis. Cerebellum 2020, 19, 192–200. [Google Scholar] [CrossRef]
- Yaldizli, Ö.; Penner, I.K.; Yonekawa, T.; Naegelin, Y.; Kuhle, J.; Pardini, M.; Chard, D.T.; Stippich, C.; Kira, J.I.; Bendfeldt, K.; et al. The association between olfactory bulb volume, cognitive dysfunction, physical disability and depression in multiple sclerosis. Eur. J. Neurol. 2016, 23, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Asok, A.; Leroy, F.; Rayman, J.B.; Kandel, E.R. Molecular Mechanisms of the Memory Trace. Trends Neurosci. 2019, 42, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Hsu, K.S. Progress in understanding the factors regulating reversibility of long-term potentiation. Rev. Neurosci. 2001, 12, 51–68. [Google Scholar] [CrossRef]
- Milnerwood, A.J.; Raymond, L.A. Early synaptic pathophysiology in neurodegeneration: Insights from Huntington’s disease. Trends Neurosci. 2010, 33, 513–523. [Google Scholar] [CrossRef]
- Calabresi, P.; Pisani, A.; Rothwell, J.; Ghiglieri, V.; Obeso, J.A.; Picconi, B. Hyperkinetic disorders and loss of synaptic downscaling. Nat. Neurosci. 2016, 19, 868–875. [Google Scholar] [CrossRef]
- Spires-Jones, T.L.; Hyman, B.T. The Intersection of Amyloid Beta and Tau at Synapses in Alzheimer’s Disease. Neuron 2014, 82, 756–771. [Google Scholar] [CrossRef] [Green Version]
- Pisani, A.; Centonze, D.; Bernardi, G.; Calabresi, P. Striatal synaptic plasticity: Implications for motor learning and Parkinson’s disease. Mov. Disord. 2005, 20, 395–402. [Google Scholar] [CrossRef]
- Scullion, S.E.; Barker, G.R.I.; Warburton, E.C.; Randall, A.D.; Brown, J.T. Muscarinic Receptor-Dependent Long Term Depression in the Perirhinal Cortex and Recognition Memory are Impaired in the rTg4510 Mouse Model of Tauopathy. Neurochem. Res. 2019, 44, 617–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, C.A.; Witton, J.; Nowacki, J.; Tsaneva-Atanasova, K.; Jones, M.W.; Randall, A.D.; Brown, J.T. Altered intrinsic pyramidal neuron properties and pathway-specific synaptic dysfunction underlie aberrant hippocampal network function in a mouse model of tauopathy. J. Neurosci. 2016, 36, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Hoover, B.R.; Reed, M.N.; Su, J.; Penrod, R.D.; Kotilinek, L.A.; Grant, M.K.; Pitstick, R.; Carlson, G.A.; Lanier, L.M.; Yuan, L.L.; et al. Tau Mislocalization to Dendritic Spines Mediates Synaptic Dysfunction Independently of Neurodegeneration. Neuron 2010, 68, 1067–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozzi, A.; de Iure, A.; Bagetta, V.; Tantucci, M.; Durante, V.; Quiroga-Varela, A.; Costa, C.; Di Filippo, M.; Ghiglieri, V.; Latagliata, E.C.; et al. Alpha-Synuclein Produces Early Behavioral Alterations via Striatal Cholinergic Synaptic Dysfunction by Interacting with GluN2D N -Methyl-D-Aspartate Receptor Subunit. Biol. Psychiatry 2016, 79, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Durante, V.; de Iure, A.; Loffredo, V.; Vaikath, N.; De Risi, M.; Paciotti, S.; Quiroga-Varela, A.; Chiasserini, D.; Mellone, M.; Mazzocchetti, P.; et al. Alpha-synuclein targets GluN2A NMDA receptor subunit causing striatal synaptic dysfunction and visuospatial memory alteration. Brain 2019, 142, 1365–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, N.; Iemolo, A.; Mancini, M.; Cacace, F.; De Risi, M.; Latagliata, E.C.; Ghiglieri, V.; Bellenchi, G.C.; Puglisi-Allegra, S.; Calabresi, P.; et al. Motor learning and metaplasticity in striatal neurons: Relevance for Parkinson’s disease. Brain 2018, 141, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Schirinzi, T.; Madeo, G.; Martella, G.; Maltese, M.; Picconi, B.; Calabresi, P.; Pisani, A. Early synaptic dysfunction in Parkinson’s disease: Insights from animal models. Mov. Disord. 2016, 31, 802–813. [Google Scholar] [CrossRef]
- Sepers, M.D.; Smith-Dijak, A.; LeDue, J.; Kolodziejczyk, K.; Mackie, K.; Raymond, L.A. Endocannabinoid-specific impairment in synaptic plasticity in striatum of huntington’s disease mouse model. J. Neurosci. 2018, 38, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Picconi, B.; Centonze, D.; Håkansson, K.; Bernardi, G.; Greengard, P.; Fisone, G.; Cenci, M.A.; Calabresi, P. Loss of bidirectional striatal synaptic plasticity in L-DOPA-induced dyskinesia. Nat. Neurosci. 2003, 6, 501–506. [Google Scholar] [CrossRef]
- Ghiglieri, V.; Campanelli, F.; Marino, G.; Natale, G.; Picconi, B.; Calabresi, P. Corticostriatal synaptic plasticity alterations in the R6/1 transgenic mouse model of Huntington’s disease. J. Neurosci. Res. 2019, 97, 1655–1664. [Google Scholar] [CrossRef]
- Di Filippo, M.; de Iure, A.; Giampà, C.; Chiasserini, D.; Tozzi, A.; Orvietani, P.L.; Ghiglieri, V.; Tantucci, M.; Durante, V.; Quiroga-Varela, A.; et al. Persistent activation of microglia and NADPH oxidase [corrected] drive hippocampal dysfunction in experimental multiple sclerosis. Sci. Rep. 2016, 6, 20926. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Hao, J.; Liu, R.; Turner, G.; Shi, F.D.; Rho, J.M. Inflammation-mediated memory dysfunction and effects of a ketogenic diet in a murine model of multiple sclerosis. PLoS ONE 2012, 7, e35476. [Google Scholar] [CrossRef] [Green Version]
- Planche, V.; Panatier, A.; Hiba, B.; Ducourneau, E.G.; Raffard, G.; Dubourdieu, N.; Maitre, M.; Lesté-Lasserre, T.; Brochet, B.; Dousset, V.; et al. Selective dentate gyrus disruption causes memory impairment at the early stage of experimental multiple sclerosis. Brain. Behav. Immun. 2017, 60, 240–254. [Google Scholar] [CrossRef]
- Di Filippo, M.; Chiasserini, D.; Gardoni, F.; Viviani, B.; Tozzi, A.; Giampà, C.; Costa, C.; Tantucci, M.; Zianni, E.; Boraso, M.; et al. Effects of central and peripheral inflammation on hippocampal synaptic plasticity. Neurobiol. Dis. 2013, 52, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Nisticò, R.; Mango, D.; Mandolesi, G.; Piccinin, S.; Berretta, N.; Pignatelli, M.; Feligioni, M.; Musella, A.; Gentile, A.; Mori, F.; et al. Inflammation Subverts Hippocampal Synaptic Plasticity in Experimental Multiple Sclerosis. PLoS ONE 2013, 8, e54666. [Google Scholar] [CrossRef] [Green Version]
- Picconi, B.; Bagetta, V.; Ghiglieri, V.; Paill, V.; Di Filippo, M.; Pendolino, V.; Tozzi, A.; Giampá, C.; Fusco, F.R.; Sgobio, C.; et al. Inhibition of phosphodiesterases rescues striatal long-term depression and reduces levodopa-induced dyskinesia. Brain 2011, 134, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Thiele, S.L.; Chen, B.; Lo, C.; Gertler, T.S.; Warre, R.; Surmeier, J.D.; Brotchie, J.M.; Nash, J.E. Selective loss of bi-directional synaptic plasticity in the direct and indirect striatal output pathways accompanies generation of parkinsonism and l-DOPA induced dyskinesia in mouse models. Neurobiol. Dis. 2014, 71, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belujon, P.; Lodge, D.J.; Grace, A.A. Aberrant striatal plasticity is specifically associated with dyskinesia following levodopa treatment. Mov. Disord. 2010, 25, 1568–1576. [Google Scholar] [CrossRef] [Green Version]
- Prescott, I.A.; Liu, L.D.; Dostrovsky, J.O.; Hodaie, M.; Lozano, A.M.; Hutchison, W.D. Lack of depotentiation at basal ganglia output neurons in PD patients with levodopa-induced dyskinesia. Neurobiol. Dis. 2014, 71, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, F.; Ponzo, V.; Bonnì, S.; Motta, C.; Negrão Serra, P.C.; Bozzali, M.; Caltagirone, C.; Martorana, A.; Koch, G. Long-term potentiation–like cortical plasticity is disrupted in Alzheimer’s disease patients independently from age of onset. Ann. Neurol. 2016, 80, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Motta, C.; Di Lorenzo, F.; Ponzo, V.; Pellicciari, M.C.; Bonnì, S.; Picazio, S.; Mercuri, N.B.; Caltagirone, C.; Martorana, A.; Koch, G. Transcranial magnetic stimulation predicts cognitive decline in patients with Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2018, 89, 1237–1242. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Motta, C.; Bonnì, S.; Mercuri, N.B.; Caltagirone, C.; Martorana, A.; Koch, G. LTP-like cortical plasticity is associated with verbal memory impairment in Alzheimer’s disease patients. Brain Stimul. 2019, 12, 148–151. [Google Scholar] [CrossRef]
- Battaglia, F.; Wang, H.Y.; Ghilardi, M.F.; Gashi, E.; Quartarone, A.; Friedman, E.; Nixon, R.A. Cortical Plasticity in Alzheimer’s Disease in Humans and Rodents. Biol. Psychiatry 2007, 62, 1405–1412. [Google Scholar] [CrossRef]
- Benussi, A.; Cosseddu, M.; Filareto, I.; Dell’Era, V.; Archetti, S.; Sofia Cotelli, M.; Micheli, A.; Padovani, A.; Borroni, B. Impaired long-term potentiation–like cortical plasticity in presymptomatic genetic frontotemporal dementia. Ann. Neurol. 2016, 80, 472–476. [Google Scholar] [CrossRef]
- Benussi, A.; Dell’Era, V.; Cantoni, V.; Cotelli, M.S.; Cosseddu, M.; Spallazzi, M.; Alberici, A.; Padovani, A.; Borroni, B. Neurophysiological correlates of positive and negative symptoms in frontotemporal dementia. J. Alzheimer’s Dis. 2020, 73, 1133–1142. [Google Scholar] [CrossRef]
- Ueki, Y.; Mima, T.; Kotb, M.A.; Sawada, H.; Saiki, H.; Ikeda, A.; Begum, T.; Reza, F.; Nagamine, T.; Fukuyama, H. Altered plasticity of the human motor cortex in Parkinson’s disease. Ann. Neurol. 2006, 59, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Z.; Rothwell, J.C.; Lu, C.S.; Chuang, W.L.; Chen, R.S. Abnormal bidirectional plasticity-like effects in Parkinson’s disease. Brain 2011, 134, 2312–2320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishore, A.; Joseph, T.; Velayudhan, B.; Popa, T.; Meunier, S. Early, severe and bilateral loss of LTP and LTD-like plasticity in motor cortex (M1) in de novo Parkinson’s disease. Clin. Neurophysiol. 2012, 123, 822–828. [Google Scholar] [CrossRef]
- Orth, M.; Schippling, S.; Schneider, S.A.; Bhatia, K.P.; Talelli, P.; Tabrizi, S.J.; Rothwell, J.C. Abnormal motor cortex plasticity in premanifest and very early manifest Huntington disease. J. Neurol. Neurosurg. Psychiatry 2010, 81, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Schippling, S.; Schneider, S.A.; Bhatia, K.P.; Münchau, A.; Rothwell, J.C.; Tabrizi, S.J.; Orth, M. Abnormal Motor Cortex Excitability in Preclinical and Very Early Huntington’s Disease. Biol. Psychiatry 2009, 65, 959–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzano, C.; Dinapoli, L.; Gilio, F.; Suppa, A.; Bagnato, S.; Currà, A.; Inghilleri, M.; Berardelli, A. Motor cortical excitability studied with repetitive transcranial magnetic stimulation in patients with Huntington’s disease. Clin. Neurophysiol. 2006, 117, 1677–1681. [Google Scholar] [CrossRef]
- Mori, F.; Rossi, S.; Sancesario, G.; Codecá, C.; Mataluni, G.; Monteleone, F.; Buttari, F.; Kusayanagi, H.; Castelli, M.; Motta, C.; et al. Cognitive and cortical plasticity deficits correlate with altered amyloid-Β CSF levels in multiple sclerosis. Neuropsychopharmacology 2011, 36, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Mori, F.; Kusayanagi, H.; Buttari, F.; Centini, B.; Monteleone, F.; Nicoletti, C.G.; Bernardi, G.; Di Cantogno, E.V.; Marciani, M.G.; Centonze, D. Early treatment with high-dose interferon beta-1a reverses cognitive and cortical plasticity deficits in multiple sclerosis. Funct. Neurol. 2012, 27, 163–168. [Google Scholar] [PubMed]
- Wirsching, I.; Buttmann, M.; Odorfer, T.; Volkmann, J.; Classen, J.; Zeller, D. Altered motor plasticity in an acute relapse of multiple sclerosis. Eur. J. Neurosci. 2018, 47, 251–257. [Google Scholar] [CrossRef]
- Li, S.; Jin, M.; Koeglsperger, T.; Shepardson, N.E.; Shankar, G.M.; Selkoe, D.J. Soluble a β oligomers inhibit long-term potentiation through a mechanism involving excessive activation of extrasynaptic NR2B-containing NMDA receptors. J. Neurosci. 2011, 31, 6627–6638. [Google Scholar] [CrossRef]
- Shankar, G.M.; Li, S.; Mehta, T.H.; Garcia-Munoz, A.; Shepardson, N.E.; Smith, I.; Brett, F.M.; Farrell, M.A.; Rowan, M.J.; Lemere, C.A.; et al. Amyloid-β protein dimers isolated directly from Alzheimer’s brains impair synaptic plasticity and memory. Nat. Med. 2008, 14, 837–842. [Google Scholar] [CrossRef] [Green Version]
- Gulisano, W.; Melone, M.; Li Puma, D.D.; Tropea, M.R.; Palmeri, A.; Arancio, O.; Grassi, C.; Conti, F.; Puzzo, D. The effect of amyloid-β peptide on synaptic plasticity and memory is influenced by different isoforms, concentrations, and aggregation status. Neurobiol. Aging 2018, 71, 51–60. [Google Scholar] [CrossRef]
- Mandolesi, G.; Gentile, A.; Musella, A.; Fresegna, D.; De Vito, F.; Bullitta, S.; Sepman, H.; Marfia, G.A.; Centonze, D. Synaptopathy connects inflammation and neurodegeneration in multiple sclerosis. Nat. Rev. Neurol. 2015, 11, 721–724. [Google Scholar] [CrossRef]
- Centonze, D.; Siracusano, A.; Calabresi, P.; Bernardi, G. Long-term potentiation and memory processes in the psychological works of Sigmund Freud and in the formation of neuropsychiatric symptoms. Neuroscience 2005, 130, 559–565. [Google Scholar] [CrossRef]
- Kuhn, M.; Mainberger, F.; Feige, B.; Maier, J.G.; Mall, V.; Jung, N.H.; Reis, J.; Klöppel, S.; Normann, C.; Nissen, C. State-dependent partial occlusion of cortical LTP-like plasticity in major depression. Neuropsychopharmacology 2016, 41, 2794. [Google Scholar] [CrossRef] [Green Version]
- Player, M.J.; Taylor, J.L.; Weickert, C.S.; Alonzo, A.; Sachdev, P.; Martin, D.; Mitchell, P.B.; Loo, C.K. Neuroplasticity in depressed individuals compared with healthy controls. Neuropsychopharmacology 2013, 38, 2101–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zak, N.; Moberget, T.; Bøen, E.; Boye, B.; Waage, T.R.; Dietrichs, E.; Harkestad, N.; Malt, U.F.; Westlye, L.T.; Andreassen, O.A.; et al. Longitudinal and cross-sectional investigations of long-term potentiation-like cortical plasticity in bipolar disorder type II and healthy individuals. Transl. Psychiatry 2018, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Elvsshagen, T.; Moberget, T.; Bøen, E.; Boye, B.; Englin, N.O.A.; Pedersen, P.; Andreassen, O.A.; Dietrichs, E.; Malt, U.F.; Andersson, S. Evidence for impaired neocortical synaptic plasticity in bipolar II disorder. Biol. Psychiatry 2012, 71, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Pang, F.; Liu, S.; Shen, Y.; Liu, L.; Fang, Z.; Wang, C.; Hu, Z.; Yuan, T.F. Altered Motor-Striatal Plasticity and Cortical Functioning in Patients with Schizophrenia. Neurosci. Bull. 2017, 33, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Kantrowitz, J.T.; Epstein, M.L.; Beggel, O.; Rohrig, S.; Lehrfeld, J.M.; Revheim, N.; Lehrfeld, N.P.; Reep, J.; Parker, E.; Silipo, G.; et al. Neurophysiological mechanisms of cortical plasticity impairments in schizophrenia and modulation by the NMDA receptor agonist D-serine. Brain 2016, 139, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, D.M.; Wohleb, E.S.; Duman, R.S. Emerging treatment mechanisms for depression: Focus on glutamate and synaptic plasticity. Drug Discov. Today 2016, 21, 454–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Sanacora, G.; Krystal, J.H. Altered Connectivity in Depression: GABA and Glutamate Neurotransmitter Deficits and Reversal by Novel Treatments. Neuron 2019, 102, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Lener, M.S.; Niciu, M.J.; Ballard, E.D.; Park, M.; Park, L.T.; Nugent, A.C.; Zarate, C.A. Glutamate and Gamma-Aminobutyric Acid Systems in the Pathophysiology of Major Depression and Antidepressant Response to Ketamine. Biol. Psychiatry 2017, 81, 886–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, W.K.; Storch, E.A.; Sheth, S.A. Harmonizing the neurobiology and treatment of obsessive-compulsive disorder. Am. J. Psychiatry 2021, 178, 17–29. [Google Scholar] [CrossRef]
- Mineo, D.; Cacace, F.; Mancini, M.; Vannelli, A.; Campanelli, F.; Natale, G.; Marino, G.; Cardinale, A.; Calabresi, P.; Picconi, B.; et al. Dopamine drives binge-like consumption of a palatable food in experimental Parkinsonism. Mov. Disord. 2019, 34, 821–831. [Google Scholar] [CrossRef]
- Yu, X.; Taylor, A.M.W.; Nagai, J.; Golshani, P.; Evans, C.J.; Coppola, G.; Khakh, B.S. Reducing Astrocyte Calcium Signaling In Vivo Alters Striatal Microcircuits and Causes Repetitive Behavior. Neuron 2018, 99, 1170–1187. [Google Scholar] [CrossRef] [Green Version]
- Gubert, C.; Renoir, T.; Hannan, A.J. Why Woody got the blues: The neurobiology of depression in Huntington’s disease. Neurobiol. Dis. 2020, 142, 104958. [Google Scholar] [CrossRef]
- Wright, D.J.; Gray, L.J.; Finkelstein, D.I.; Crouch, P.J.; Pow, D.; Pang, T.Y.; Li, S.; Smith, Z.M.; Francis, P.S.; Renoir, T.; et al. N-acetylcysteine modulates glutamatergic dysfunction and depressive behavior in Huntington’s disease. Hum. Mol. Genet. 2016, 25, 2923–2933. [Google Scholar] [CrossRef] [Green Version]
- Gentile, A.; Fresegna, D.; Federici, M.; Musella, A.; Rizzo, F.R.; Sepman, H.; Bullitta, S.; De Vito, F.; Haji, N.; Rossi, S.; et al. Dopaminergic dysfunction is associated with IL-1β-dependent mood alterations in experimental autoimmune encephalomyelitis. Neurobiol. Dis. 2014, 74, 347–358. [Google Scholar] [CrossRef]
- Gentile, A.; Fresegna, D.; Musella, A.; Sepman, H.; Bullitta, S.; De Vito, F.; Fantozzi, R.; Usiello, A.; Maccarrone, M.; Mercuri, N.B.; et al. Interaction between interleukin-1β and type-1 cannabinoid receptor is involved in anxiety-like behavior in experimental autoimmune encephalomyelitis. J. Neuroinflamm. 2016, 13, 231. [Google Scholar] [CrossRef] [Green Version]
- Haji, N.; Mandolesi, G.; Gentile, A.; Sacchetti, L.; Fresegna, D.; Rossi, S.; Musella, A.; Sepman, H.; Motta, C.; Studer, V.; et al. TNF-α-mediated anxiety in a mouse model of multiple sclerosis. Exp. Neurol. 2012, 237, 296–303. [Google Scholar] [CrossRef]
- Rossi, S.; Studer, V.; Motta, C.; Polidoro, S.; Perugini, J.; Macchiarulo, G.; Giovannetti, A.M.; Pareja-Gutierrez, L.; Calò, A.; Colonna, I.; et al. Neuroinflammation drives anxiety and depression in relapsing-remitting multiple sclerosis. Neurology 2017, 89, 1338–1347. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Eisenberger, N.I.; Inagaki, T.K.; Mashal, N.M.; Irwin, M.R. Inflammation and social experience: An inflammatory challenge induces feelings of social disconnection in addition to depressed mood. Brain. Behav. Immun. 2010, 24, 558–563. [Google Scholar] [CrossRef] [Green Version]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimer’s Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Chao, Y.X.; West, A.; Chan, L.L.; Poewe, W.; Jankovic, J. Parkinson disease and the immune system—Associations, mechanisms and therapeutics. Nat. Rev. Neurol. 2020, 16, 303–318. [Google Scholar] [CrossRef]
- Ledo, J.H.; Azevedo, E.P.; Beckman, D.; Ribeiro, F.C.; Santos, L.E.; Razolli, D.S.; Kincheski, G.C.; Melo, H.M.; Bellio, M.; Teixeira, A.L.; et al. Cross talk between brain innate immunity and serotonin signaling underlies depressive-like behavior induced by Alzheimer’s amyloid-β oligomers in mice. J. Neurosci. 2016, 36, 12106–12116. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Spampinato, S.F.; Morgese, M.G.; Tascedda, F.; Salluzzo, M.G.; Giambirtone, M.C.; Caruso, G.; Munafò, A.; Torrisi, S.A.; Leggio, G.M.; et al. Neurobiological links between depression and AD: The role of TGF-β1 signaling as a new pharmacological target. Pharmacol. Res. 2018, 130, 374–384. [Google Scholar] [CrossRef]
- Jack, C.R.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Paquet, C.; Magnin, E.; Wallon, D.; Troussière, A.C.; Dumurgier, J.; Jager, A.; Bellivier, F.; Bouaziz-Amar, E.; Blanc, F.; Beaufils, E.; et al. Utility of CSF biomarkers in psychiatric disorders: A national multicentre prospective study. Alzheimer’s Res. Ther. 2016, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Steffens, D.C.; Au, R.; Folstein, M.; Summergrad, P.; Yee, J.; Rosenberg, I.; Mwamburi, D.M.; Wei, Q.Q. Amyloid-associated depression: A prodromal depression of Alzheimer disease? Arch. Gen. Psychiatry 2008, 65, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Direk, N.; Schrijvers, E.M.C.; de Bruijn, R.F.A.G.; Mirza, S.; Hofman, A.; Ikram, M.A.; Tiemeier, H. Plasma amyloid β, depression, and dementia in community-dwelling elderly. J. Psychiatr. Res. 2013, 47, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Karikari, T.K.; Pascoal, T.A.; Ashton, N.J.; Janelidze, S.; Benedet, A.L.; Rodriguez, J.L.; Chamoun, M.; Savard, M.; Kang, M.S.; Therriault, J.; et al. Blood phosphorylated tau 181 as a biomarker for Alzheimer’s disease: A diagnostic performance and prediction modelling study using data from four prospective cohorts. Lancet Neurol. 2020, 19, 422–433. [Google Scholar] [CrossRef]
- Neumann, M.; Sampathu, D.M.; Kwong, L.K.; Truax, A.C.; Micsenyi, M.C.; Chou, T.T.; Bruce, J.; Schuck, T.; Grossman, M.; Clark, C.M.; et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 2006, 314, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Foulds, P.; McAuley, E.; Gibbons, L.; Davidson, Y.; Pickering-Brown, S.M.; Neary, D.; Snowden, J.S.; Allsop, D.; Mann, D.M.A. TDP-43 protein in plasma may index TDP-43 brain pathology in Alzheimer’s disease and frontotemporal lobar degeneration. Acta Neuropathol. 2008, 116, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Bourbouli, M.; Rentzos, M.; Bougea, A.; Zouvelou, V.; Constantinides, V.C.; Zaganas, I.; Evdokimidis, I.; Kapaki, E.; Paraskevas, G.P. Cerebrospinal Fluid TAR DNA-Binding Protein 43 Combined with Tau Proteins as a Candidate Biomarker for Amyotrophic Lateral Sclerosis and Frontotemporal Dementia Spectrum Disorders. Dement. Geriatr. Cogn. Disord. 2017, 44, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Foulds, P.G.; Davidson, Y.; Mishra, M.; Hobson, D.J.; Humphreys, K.M.; Taylor, M.; Johnson, N.; Weintraub, S.; Akiyama, H.; Arai, T.; et al. Plasma phosphorylated-TDP-43 protein levels correlate with brain pathology in frontotemporal lobar degeneration. Acta Neuropathol. 2009, 118, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, T.; Baba, H.; Maeshima, H.; Shimano, T.; Inoue, M.; Ishiguro, M.; Yasuda, S.; Shukuzawa, H.; Suzuki, T.; Arai, H. Serum levels of TDP-43 in late-life patients with depressive episode. J. Affect. Disord. 2019, 250, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Foulds, P.G.; Diggle, P.; Mitchell, J.D.; Parker, A.; Hasegawa, M.; Masuda-Suzukake, M.; Mann, D.M.A.; Allsop, D. A longitudinal study on a-synuclein in blood plasma as a biomarker for Parkinson’s disease. Sci. Rep. 2013, 3, 2540. [Google Scholar] [CrossRef]
- Ishii, R.; Tokuda, T.; Tatebe, H.; Ohmichi, T.; Kasai, T.; Nakagawa, M.; Mizuno, T.; El-Agnaf, O.M.A. Decrease in plasma levels of α-synuclein is evident in patients with Parkinson’s disease after elimination of heterophilic antibody interference. PLoS ONE 2015, 10, e0123162. [Google Scholar] [CrossRef]
- Malec-Litwinowicz, M.; Plewka, A.; Plewka, D.; Bogunia, E.; Morek, M.; Szczudlik, A.; Szubiga, M.; Rudzińska-Bar, M. The relation between plasma α-synuclein level and clinical symptoms or signs of Parkinson’s disease. Neurol. Neurochir. Pol. 2018, 52, 243–251. [Google Scholar] [CrossRef]
- Barbour, R.; Kling, K.; Anderson, J.P.; Banducci, K.; Cole, T.; Diep, L.; Fox, M.; Goldstein, J.M.; Soriano, F.; Seubert, P.; et al. Red blood cells are the major source of alpha-synuclein in blood. Neurodegener. Dis. 2008, 5, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Parnetti, L.; Gaetani, L.; Eusebi, P.; Paciotti, S.; Hansson, O.; El-Agnaf, O.; Mollenhauer, B.; Blennow, K.; Calabresi, P. CSF and blood biomarkers for Parkinson’s disease. Lancet Neurol. 2019, 18, 573–586. [Google Scholar] [CrossRef]
- Rotter, A.; Lenz, B.; Pitsch, R.; Richter-Schmidinger, T.; Kornhuber, J.; Rhein, C. Alpha-synuclein RNA expression is increased in major depression. Int. J. Mol. Sci. 2019, 20, 2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiguro, M.; Baba, H.; Maeshima, H.; Shimano, T.; Inoue, M.; Ichikawa, T.; Yasuda, S.; Shukuzawa, H.; Suzuki, T.; Arai, H. Increased Serum Levels of α-Synuclein in Patients with Major Depressive Disorder. Am. J. Geriatr. Psychiatry 2019, 27, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.M.; Egbufoama, S.; Vawter, M.P. SNAP-25 reduction in the hippocampus of patients with schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2003, 27, 411–417. [Google Scholar] [CrossRef]
- Fairfoul, G.; McGuire, L.I.; Pal, S.; Ironside, J.W.; Neumann, J.; Christie, S.; Joachim, C.; Esiri, M.; Evetts, S.G.; Rolinski, M.; et al. Alpha-synuclein RT-QuIC in the CSF of patients with alpha-synucleinopathies. Ann. Clin. Transl. Neurol. 2016, 3, 812–818. [Google Scholar] [CrossRef]
- Imitola, J.; Chitnis, T.; Khoury, S.J. Cytokines in multiple sclerosis: From bench to bedside. Pharmacol. Ther. 2005, 106, 163–177. [Google Scholar] [CrossRef]
- Bruno, A.; Dolcetti, E.; Rizzo, F.R.; Fresegna, D.; Musella, A.; Gentile, A.; De Vito, F.; Caioli, S.; Guadalupi, L.; Bullitta, S.; et al. Inflammation-Associated Synaptic Alterations as Shared Threads in Depression and Multiple Sclerosis. Front. Cell. Neurosci. 2020, 14, 169. [Google Scholar] [CrossRef]
- Orlovska-Waast, S.; Köhler-Forsberg, O.; Brix, S.W.; Nordentoft, M.; Kondziella, D.; Krogh, J.; Benros, M.E. Cerebrospinal fluid markers of inflammation and infections in schizophrenia and affective disorders: A systematic review and meta-analysis. Mol. Psychiatry 2019, 24, 869–887. [Google Scholar] [CrossRef] [Green Version]
- Gallego, J.A.; Blanco, E.A.; Husain-Krautter, S.; Madeline Fagen, E.; Moreno-Merino, P.; del Ojo-Jiménez, J.A.; Ahmed, A.; Rothstein, T.L.; Lencz, T.; Malhotra, A.K. Cytokines in cerebrospinal fluid of patients with schizophrenia spectrum disorders: New data and an updated meta-analysis. Schizophr. Res. 2018, 202, 64–71. [Google Scholar] [CrossRef]
- Dahl, J.; Ormstad, H.; Aass, H.C.D.; Malt, U.F.; Bendz, L.T.; Sandvik, L.; Brundin, L.; Andreassen, O.A. The plasma levels of various cytokines are increased during ongoing depression and are reduced to normal levels after recovery. Psychoneuroendocrinology 2014, 45, 77–86. [Google Scholar] [CrossRef]
- Tong, Q.; Zhang, L.; Yuan, Y.; Jiang, S.; Zhang, R.; Xu, Q.; Ding, J.; Li, D.; Zhou, X.; Zhang, K. Reduced plasma serotonin and 5-hydroxyindoleacetic acid levels in Parkinson’s disease are associated with nonmotor symptoms. Park. Relat. Disord. 2015, 21, 882–887. [Google Scholar] [CrossRef]
- Aeinehband, S.; Brenner, P.; Ståhl, S.; Bhat, M.; Fidock, M.D.; Khademi, M.; Olsson, T.; Engberg, G.; Jokinen, J.; Erhardt, S.; et al. Cerebrospinal fluid kynurenines in multiple sclerosis; relation to disease course and neurocognitive symptoms. Brain. Behav. Immun. 2016, 51, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Knorr, U.; Simonsen, A.H.; Zetterberg, H.; Blennow, K.; Hasselbalch, S.G.; Kessing, L.V. Biomarkers in cerebrospinal fluid of patients with bipolar disorder versus healthy individuals: A systematic review. Eur. Neuropsychopharmacol. 2018, 28, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Gaetani, L.; Blennow, K.; Calabresi, P.; Di Filippo, M.; Parnetti, L.; Zetterberg, H. Neurofilament light chain as a biomarker in neurological disorders. J. Neurol. Neurosurg. Psychiatry 2019, 90, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, N.; Andreasson, U.; Zetterberg, H.; Blennow, K.; Weiner, M.W.; Aisen, P.; Toga, A.W.; Petersen, R.; Jack, C.R.; Jagust, W.; et al. Association of plasma neurofilament light with neurodegeneration in patients with Alzheimer disease. JAMA Neurol. 2017, 74, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Lista, S.; Toschi, N.; Baldacci, F.; Zetterberg, H.; Blennow, K.; Kilimann, I.; Teipel, S.J.; Cavedo, E.; dos Santos, A.M.; Epelbaum, S.; et al. Diagnostic accuracy of CSF neurofilament light chain protein in the biomarker-guided classification system for Alzheimer’s disease. Neurochem. Int. 2017, 108, 355–360. [Google Scholar] [CrossRef] [Green Version]
- De Jong, D.; Jansen, R.W.M.M.; Pijnenburg, Y.A.L.; Van Geel, W.J.A.; Borm, G.F.; Kremer, H.P.H.; Verbeek, M.M. CSF neurofilament proteins in the differential diagnosis of dementia. J. Neurol. Neurosurg. Psychiatry 2007, 78, 936–938. [Google Scholar] [CrossRef] [Green Version]
- Kuhle, J.; Plattner, K.; Bestwick, J.P.; Lindberg, R.L.; Ramagopalan, S.V.; Norgren, N.; Nissim, A.; Malaspina, A.; Leppert, D.; Giovannoni, G.; et al. A comparative study of CSF neurofilament light and heavy chain protein in MS. Mult. Scler. J. 2013, 19, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Portelius, E.; Zetterberg, H.; Skillbäck, T.; Törnqvist, U.; Andreasson, U.; Trojanowski, J.Q.; Weiner, M.W.; Shaw, L.M.; Mattsson, N.; Blennow, K. Cerebrospinal fluid neurogranin: Relation to cognition and neurodegeneration in Alzheimer’s disease. Brain 2015, 138, 3373–3385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorens, F.; Thüne, K.; Tahir, W.; Kanata, E.; Diaz-Lucena, D.; Xanthopoulos, K.; Kovatsi, E.; Pleschka, C.; Garcia-Esparcia, P.; Schmitz, M.; et al. YKL-40 in the brain and cerebrospinal fluid of neurodegenerative dementias. Mol. Neurodegener. 2017, 12, 83. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.N.; Swarovski, M.S.; Linortner, P.; Shahid, M.; Zuckerman, A.J.; Wang, Q.; Channappa, D.; Minhas, P.S.; Mhatre, S.D.; Plowey, E.D.; et al. Soluble TREM2 is elevated in parkinson’s disease subgroups with increased csf tau. Brain 2020, 143, 932–943. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menculini, G.; Chipi, E.; Paolini Paoletti, F.; Gaetani, L.; Nigro, P.; Simoni, S.; Mancini, A.; Tambasco, N.; Di Filippo, M.; Tortorella, A.; et al. Insights into the Pathophysiology of Psychiatric Symptoms in Central Nervous System Disorders: Implications for Early and Differential Diagnosis. Int. J. Mol. Sci. 2021, 22, 4440. https://doi.org/10.3390/ijms22094440
Menculini G, Chipi E, Paolini Paoletti F, Gaetani L, Nigro P, Simoni S, Mancini A, Tambasco N, Di Filippo M, Tortorella A, et al. Insights into the Pathophysiology of Psychiatric Symptoms in Central Nervous System Disorders: Implications for Early and Differential Diagnosis. International Journal of Molecular Sciences. 2021; 22(9):4440. https://doi.org/10.3390/ijms22094440
Chicago/Turabian StyleMenculini, Giulia, Elena Chipi, Federico Paolini Paoletti, Lorenzo Gaetani, Pasquale Nigro, Simone Simoni, Andrea Mancini, Nicola Tambasco, Massimiliano Di Filippo, Alfonso Tortorella, and et al. 2021. "Insights into the Pathophysiology of Psychiatric Symptoms in Central Nervous System Disorders: Implications for Early and Differential Diagnosis" International Journal of Molecular Sciences 22, no. 9: 4440. https://doi.org/10.3390/ijms22094440
APA StyleMenculini, G., Chipi, E., Paolini Paoletti, F., Gaetani, L., Nigro, P., Simoni, S., Mancini, A., Tambasco, N., Di Filippo, M., Tortorella, A., & Parnetti, L. (2021). Insights into the Pathophysiology of Psychiatric Symptoms in Central Nervous System Disorders: Implications for Early and Differential Diagnosis. International Journal of Molecular Sciences, 22(9), 4440. https://doi.org/10.3390/ijms22094440