
Hydrocephalus Following Experimental Subarachnoid Hemorrhage in Rats with Different Aerobic Capacity

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
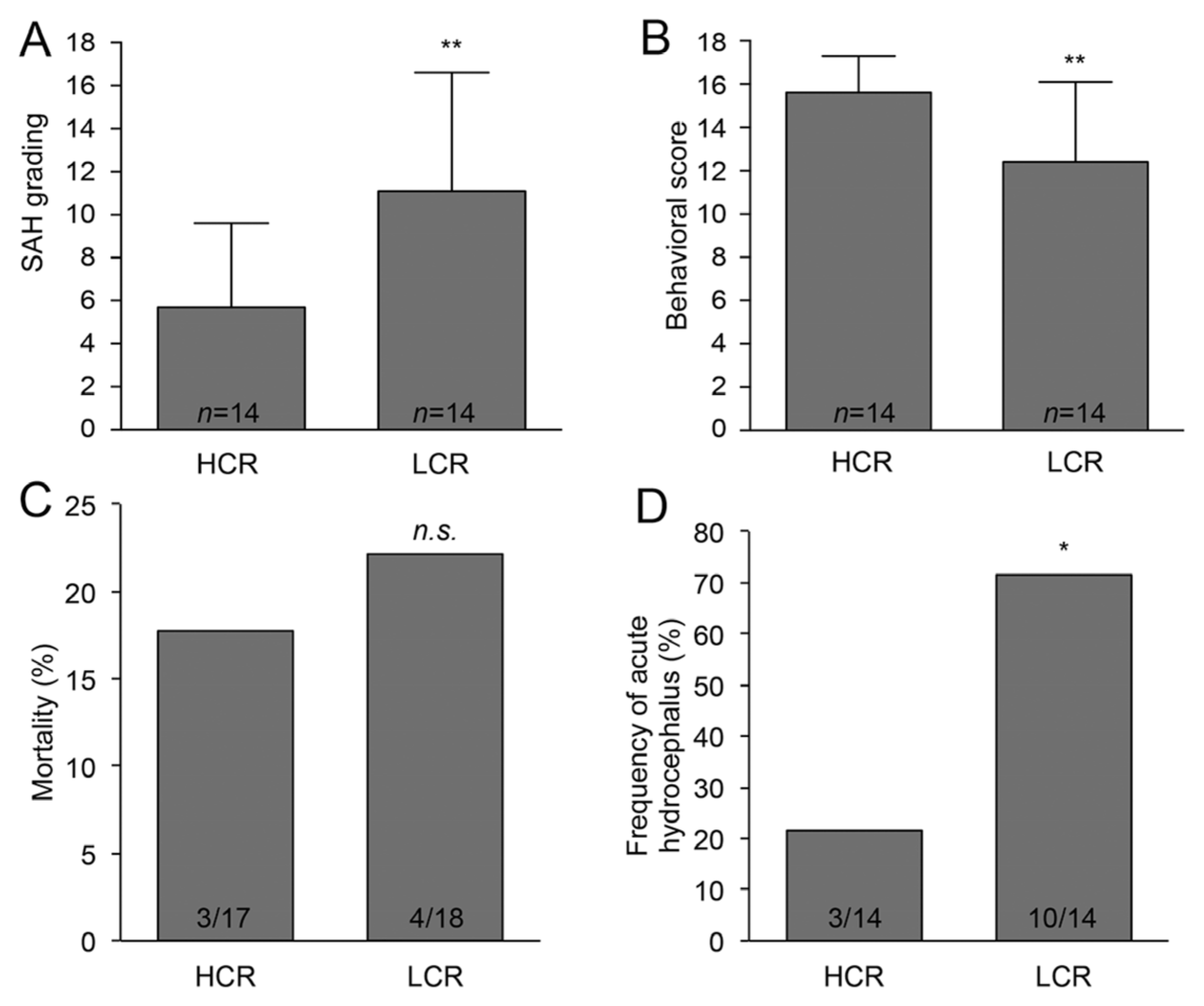
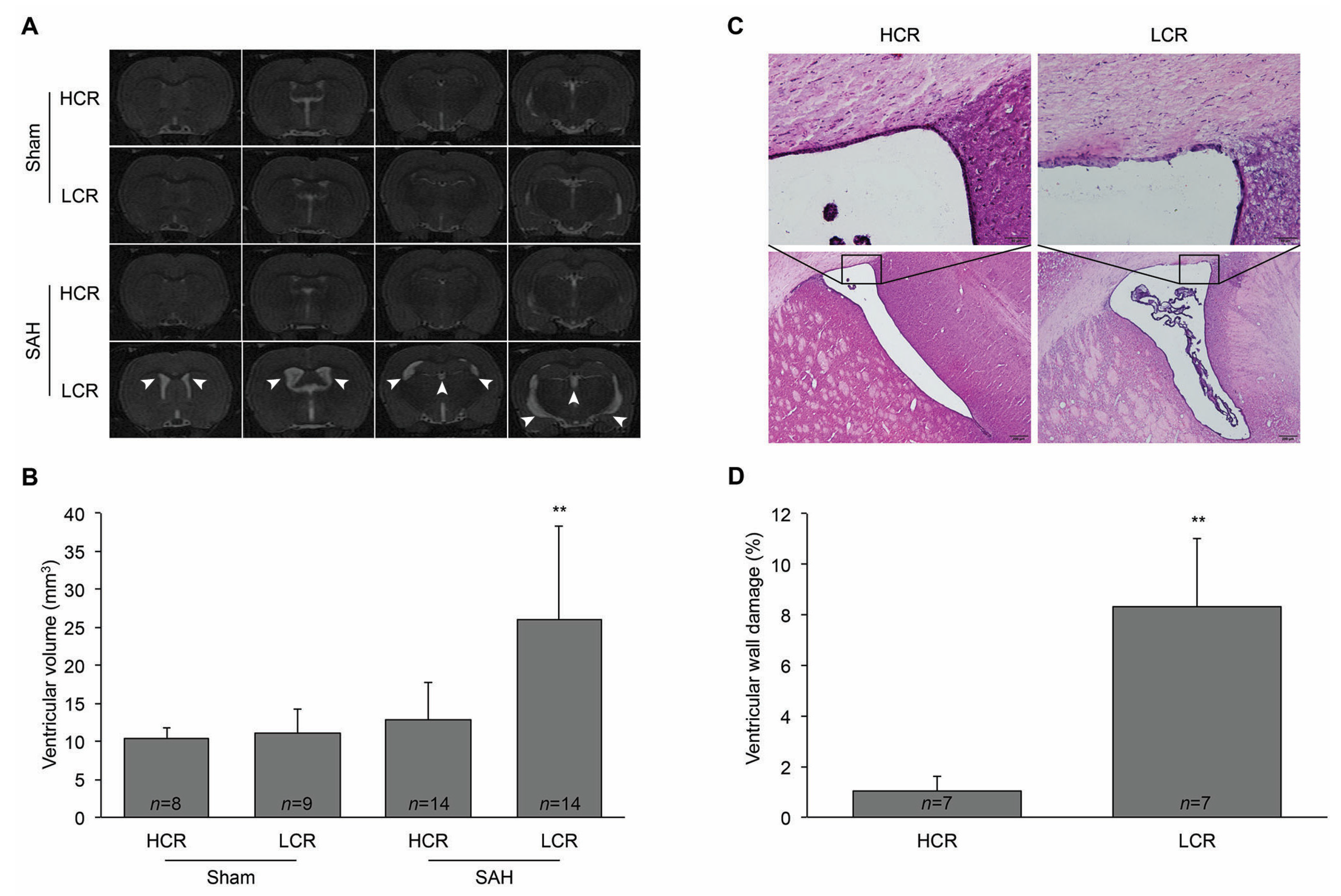
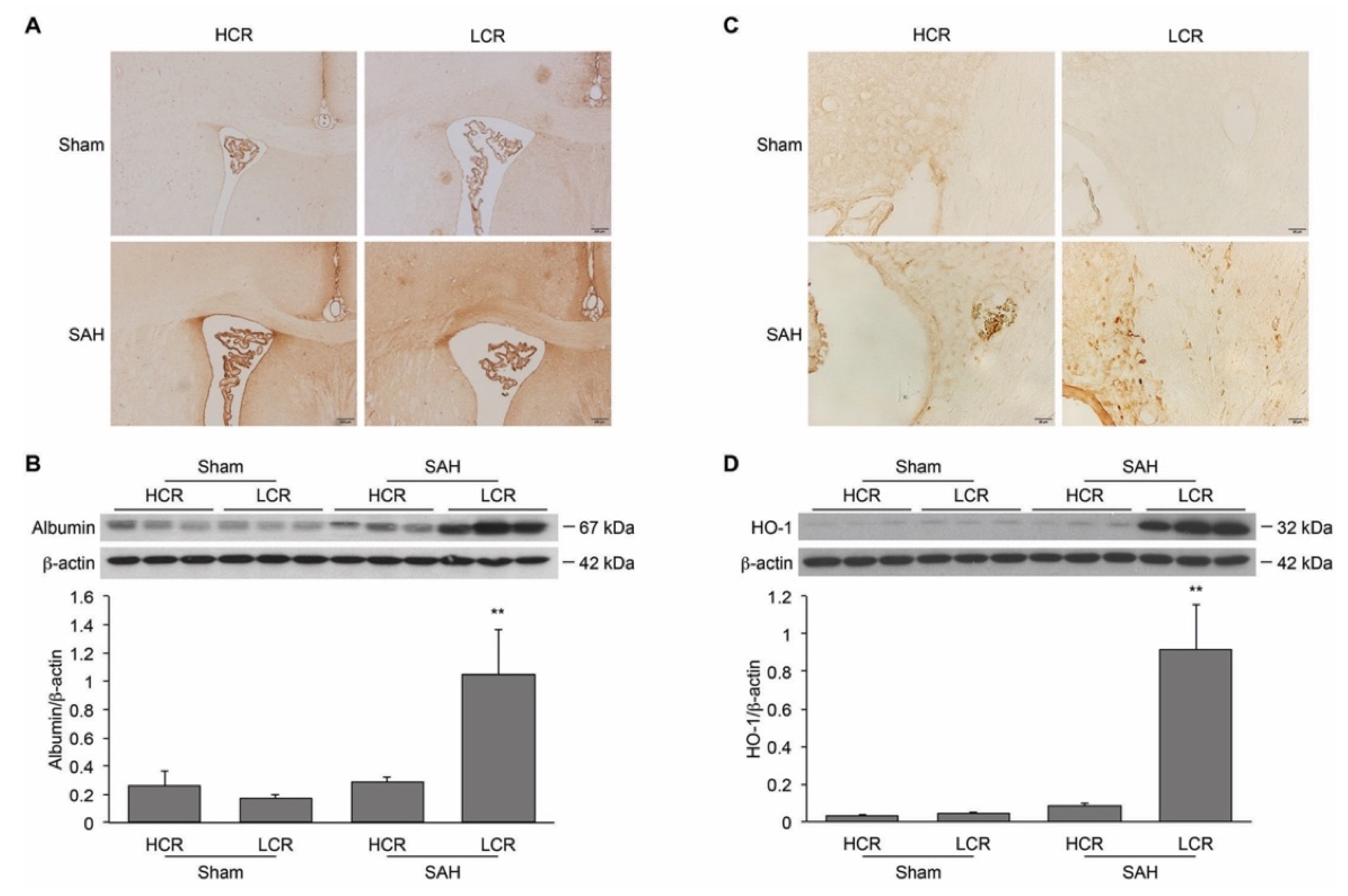
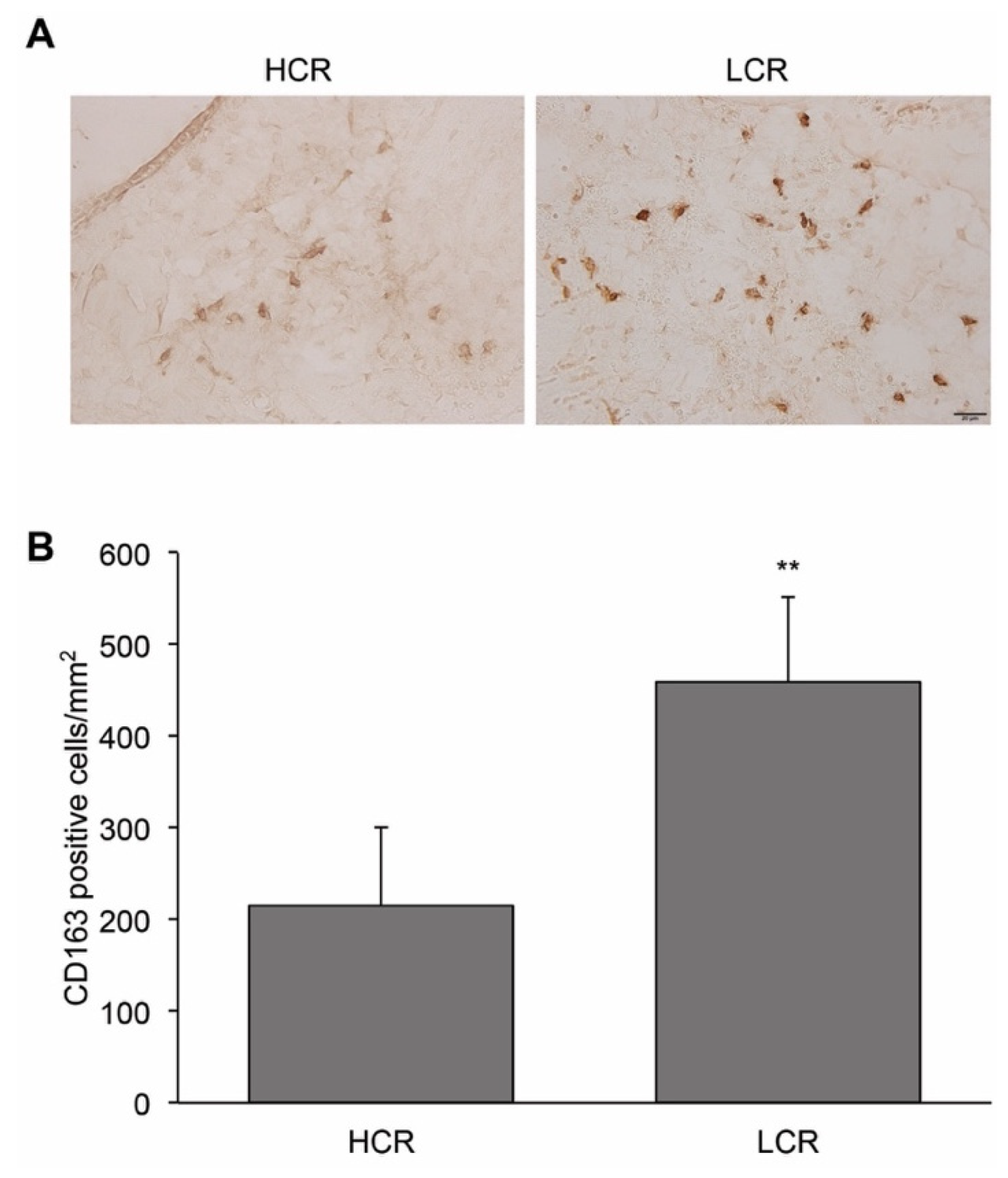
2. Results
3. Discussion
4. Materials and Methods
4.1. The Animal Preparation and SAH Induction
4.2. MRI Investigation
4.3. SAH Grading
4.4. Behavioral Tests
4.5. Ventricular Wall Damage Evaluation
4.6. Immunohistochemistry
4.7. Western Blotting Analysis
4.8. Cell Counting
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SAH | subarachnoid hemorrhage |
| HCR | high-capacity runners |
| LCR | low-capacity runners |
| HO-1 | heme oxgenase-1 |
| EBI | early brain injury |
| ICP | intracranial pressure |
| BBB | blood–brain barrier |
| IVH | intraventricular hemorrhage |
| Hp | haptoglobin |
| Hb | hemoglobin |
| CSF | cerebrospinal fluid |
References
- Cahill, J.; Zhang, J.H. Subarachnoid hemorrhage: Is it time for a new direction? Stroke 2009, 40, S86–S87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arboix, A.; Marti-Vilalta, J.L. Predictive clinical factors of very early in-hospital mortality in subarachnoid hemorrhage. Clin. Neurol. Neurosurg. 1999, 101, 100–105. [Google Scholar] [CrossRef]
- Kavanagh, T.; Mertens, D.J.; Hamm, L.F.; Beyene, J.; Kenned, J.; Corey, P.; Shephard, R.J. Peak oxygen intake and cardiac mortality in women referred for cardiac rehabilitation. J. Am. Coll. Cardiol. 2003, 42, 2139–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurl, S.; Sivenius, J.; Makikallio, T.H.; Rauramaa, R.; Laukkanen, J.A. Exercise workload, cardiovascular risk factor evaluation and the risk of stroke in middle-aged men. J. Intern. Med. 2009, 265, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.; Prakash, M.; Froelicher, V.; Do, D.; Partington, S.; Atwood, J.E. Exercise capacity and mortality among men referred for exercise testing. N. Engl. J. Med. 2002, 346, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Al Rifai, M.; Blaha, M.J.; Ahmed, A.; Almasoudi, F.; Johansen, M.C.; Qureshi, W.; Sakr, S.; Virani, S.S.; Brawner, C.A.; Ehrman, J.K.; et al. Cardiorespiratory Fitness and Incident Stroke Types: The FIT (Henry Ford ExercIse Testing) Project. Mayo Clin. Proc. 2020, 95, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Lindbohm, J.V.; Rautalin, I.; Jousilahti, P.; Salomaa, V.; Kaprio, J.; Korja, M. Physical activity associates with subarachnoid hemorrhage risk- a population-based long-term cohort study. Sci. Rep. 2019, 9, 9219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Liu, W.; Koch, L.G.; Britton, S.L.; Keep, R.F.; Xi, G.; Hua, Y. Susceptibility to intracerebral hemorrhage-induced brain injury segregates with low aerobic capacity in rats. Neurobiol. Dis. 2013, 49, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Du, H.; Ni, W.; Koch, L.G.; Britton, S.L.; Keep, R.F.; Xi, G.; Hua, Y. Iron-induced necrotic brain cell death in rats with different aerobic capacity. Transl. Stroke Res. 2015, 6, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Gao, F.; Zheng, M.; Koch, L.G.; Britton, S.L.; Keep, R.F.; Xi, G.; Hua, Y. Effects of Aerobic Capacity on Thrombin-Induced Hydrocephalus and White Matter Injury. Acta Neurochir. Suppl. 2016, 121, 379–384. [Google Scholar]
- van Asch, C.J.; van der Schaaf, I.C.; Rinkel, G.J. Acute hydrocephalus and cerebral perfusion after aneurysmal subarachnoid hemorrhage. AJNR Am. J. Neuroradiol. 2010, 31, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Hasan, D.; Vermeulen, M.; Wijdicks, E.F.; Hijdra, A.; van Gijn, J. Management problems in acute hydrocephalus after subarachnoid hemorrhage. Stroke 1989, 20, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Lucke-Wold, B.P.; Logsdon, A.F.; Manoranjan, B.; Turner, R.C.; McConnell, E.; Vates, G.E.; Huber, J.D.; Rosen, C.L.; Simard, J.M. Aneurysmal Subarachnoid Hemorrhage and Neuroinflammation: A Comprehensive Review. Int. J. Mol. Sci. 2016, 17, 497. [Google Scholar] [CrossRef] [PubMed]
- Abraham, N.G.; Drummond, G. CD163-Mediated hemoglobin-heme uptake activates macrophage HO-1, providing an antiinflammatory function. Circ. Res. 2006, 99, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, J.H.; Etzerodt, A.; Svendsen, P.; Moestrup, S.K. The haptoglobin-CD163-heme oxygenase-1 pathway for hemoglobin scavenging. Oxid. Med. Cell Longev. 2013, 2013, 523652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Yamaguchi, M.; Zhou, C.; Calvert, J.W.; Tang, J.; Zhang, J.H. Neurovascular protection reduces early brain injury after subarachnoid hemorrhage. Stroke 2004, 35, 2412–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Keep, R.F.; He, Y.; Sagher, O.; Hua, Y.; Xi, G. Hemoglobin and iron handling in brain after subarachnoid hemorrhage and the effect of deferoxamine on early brain injury. J. Cereb. Blood Flow Metab. 2010, 30, 1793–1803. [Google Scholar] [CrossRef] [Green Version]
- Rosen, D.S.; Amidei, C.; Tolentino, J.; Reilly, C.; Macdonald, R.L. Subarachnoid clot volume correlates with age, neurological grade, and blood pressure. Neurosurgery 2007, 60, 259–266. [Google Scholar] [CrossRef]
- Wisloff, U.; Najjar, S.M.; Ellingsen, O.; Haram, P.M.; Swoap, S.; Al-Share, Q.; Fernstrom, M.; Rezaei, K.; Lee, S.J.; Koch, L.G.; et al. Cardiovascular risk factors emerge after artificial selection for low aerobic capacity. Science 2005, 307, 418–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockard, M.M.; Gopinathannair, R.; Paton, C.M.; Phares, D.A.; Hagberg, J.M. Exercise training-induced changes in coagulation factors in older adults. Med. Sci. Sports Exerc. 2007, 39, 587–592. [Google Scholar] [CrossRef]
- Wang, J.S.; Chang, Y.L.; Chen, Y.C.; Tsai, H.H.; Fu, T.C. Effects of normoxic and hypoxic exercise regimens on monocyte-mediated thrombin generation in sedentary men. Clin. Sci. 2015, 129, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Banizs, B.; Pike, M.M.; Millican, C.L.; Ferguson, W.B.; Komlosi, P.; Sheetz, J.; Bell, P.D.; Schwiebert, E.M.; Yoder, B.K. Dysfunctional cilia lead to altered ependyma and choroid plexus function, and result in the formation of hydrocephalus. Development 2005, 132, 5329–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egashira, Y.; Zhao, H.; Hua, Y.; Keep, R.F.; Xi, G. White Matter Injury After Subarachnoid Hemorrhage: Role of Blood-Brain Barrier Disruption and Matrix Metalloproteinase-9. Stroke 2015, 46, 2909–2915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, T.; Jadhav, V.; Ayer, R.; Chen, W.; Suzuki, H.; Zhang, J.H. Thrombin inhibition by argatroban ameliorates early brain injury and improves neurological outcomes after experimental subarachnoid hemorrhage in rats. Stroke 2009, 40, 1530–1532. [Google Scholar] [CrossRef] [Green Version]
- Keep, R.F.; Andjelkovic, A.V.; Xiang, J.; Stamatovic, S.M.; Antonetti, D.A.; Hua, Y.; Xi, G. Brain endothelial cell junctions after cerebral hemorrhage: Changes, mechanisms and therapeutic targets. J. Cereb. Blood Flow Metab. 2018, 38, 1255–1275. [Google Scholar] [CrossRef] [PubMed]
- Turgut, M.; Erdogan, S.; Ergin, K.; Serter, M. Melatonin ameliorates blood-brain barrier permeability, glutathione, and nitric oxide levels in the choroid plexus of the infantile rats with kaolin-induced hydrocephalus. Brain Res. 2007, 1175, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zheng, M.; Hua, Y.; Chen, G.; Keep, R.F.; Xi, G. Hematoma Changes During Clot Resolution After Experimental Intracerebral Hemorrhage. Stroke 2016, 47, 1626–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Leclerc, J.L.; Lampert, A.S.; Loyola Amador, C.; Schlakman, B.; Vasilopoulos, T.; Svendsen, P.; Moestrup, S.K.; Dore, S. The absence of the CD163 receptor has distinct temporal influences on intracerebral hemorrhage outcomes. J. Cereb. Blood Flow Metab. 2018, 38, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Chen-Roetling, J.; Regan, R.F. Haptoglobin increases the vulnerability of CD163-expressing neurons to hemoglobin. J. Neurochem. 2016, 139, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Cao, S.; Hua, Y.; Keep, R.F.; Huang, Y.; Xi, G. CD163 Expression in Neurons After Experimental Intracerebral Hemorrhage. Stroke 2017, 48, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Okubo, S.; Strahle, J.; Keep, R.F.; Hua, Y.; Xi, G. Subarachnoid hemorrhage-induced hydrocephalus in rats. Stroke 2013, 44, 547–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, G.; Strahle, J.; Hua, Y.; Keep, R.F. Progress in translational research on intracerebral hemorrhage: Is there an end in sight? Prog. Neurobiol. 2014, 115, 45–63. [Google Scholar] [CrossRef] [Green Version]
- Schallner, N.; Pandit, R.; LeBlanc, R., 3rd; Thomas, A.J.; Ogilvy, C.S.; Zuckerbraun, B.S.; Gallo, D.; Otterbein, L.E.; Hanafy, K.A. Microglia regulate blood clearance in subarachnoid hemorrhage by heme oxygenase-1. J. Clin. Investig. 2015, 125, 2609–2625. [Google Scholar] [CrossRef] [Green Version]
- Sharp, F.R.; Zhan, X.; Liu, D.Z. Heat shock proteins in the brain: Role of Hsp70, Hsp 27, and HO-1 (Hsp32) and their therapeutic potential. Transl. Stroke Res. 2013, 4, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Olkowski, B.F.; Shah, S.O. Early Mobilization in the Neuro-ICU: How Far Can We Go? Neurocrit. Care 2017, 27, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, W.J.; Khajeh, L.; Ribbers, G.M.; Heijenbrok-Kal, M.H.; Sneekes, E.; van Kooten, F.; Neggers, S.; van den Berg-Emons, R.J. People With Aneurysmal Subarachnoid Hemorrhage Have Low Physical Fitness and Can Be Predisposed to Inactive and Sedentary Lifestyles. Phys. Ther. 2019, 99, 904–914. [Google Scholar] [CrossRef]
- Wu, J.; Hua, Y.; Keep, R.F.; Nakamura, T.; Hoff, J.T.; Xi, G. Iron and iron-handling proteins in the brain after intracerebral hemorrhage. Stroke 2003, 34, 2964–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyota, Y.; Shishido, H.; Ye, F.; Koch, L.G.; Britton, S.L.; Garton, H.J.L.; Keep, R.F.; Xi, G.; Hua, Y. Hydrocephalus Following Experimental Subarachnoid Hemorrhage in Rats with Different Aerobic Capacity. Int. J. Mol. Sci. 2021, 22, 4489. https://doi.org/10.3390/ijms22094489
Toyota Y, Shishido H, Ye F, Koch LG, Britton SL, Garton HJL, Keep RF, Xi G, Hua Y. Hydrocephalus Following Experimental Subarachnoid Hemorrhage in Rats with Different Aerobic Capacity. International Journal of Molecular Sciences. 2021; 22(9):4489. https://doi.org/10.3390/ijms22094489
Chicago/Turabian StyleToyota, Yasunori, Hajime Shishido, Fenghui Ye, Lauren G. Koch, Steven L. Britton, Hugh J. L. Garton, Richard F. Keep, Guohua Xi, and Ya Hua. 2021. "Hydrocephalus Following Experimental Subarachnoid Hemorrhage in Rats with Different Aerobic Capacity" International Journal of Molecular Sciences 22, no. 9: 4489. https://doi.org/10.3390/ijms22094489
APA StyleToyota, Y., Shishido, H., Ye, F., Koch, L. G., Britton, S. L., Garton, H. J. L., Keep, R. F., Xi, G., & Hua, Y. (2021). Hydrocephalus Following Experimental Subarachnoid Hemorrhage in Rats with Different Aerobic Capacity. International Journal of Molecular Sciences, 22(9), 4489. https://doi.org/10.3390/ijms22094489