
The Role of Eph Receptors and Ephrins in Corneal Physiology and Diseases


Abstract
:1. Introduction
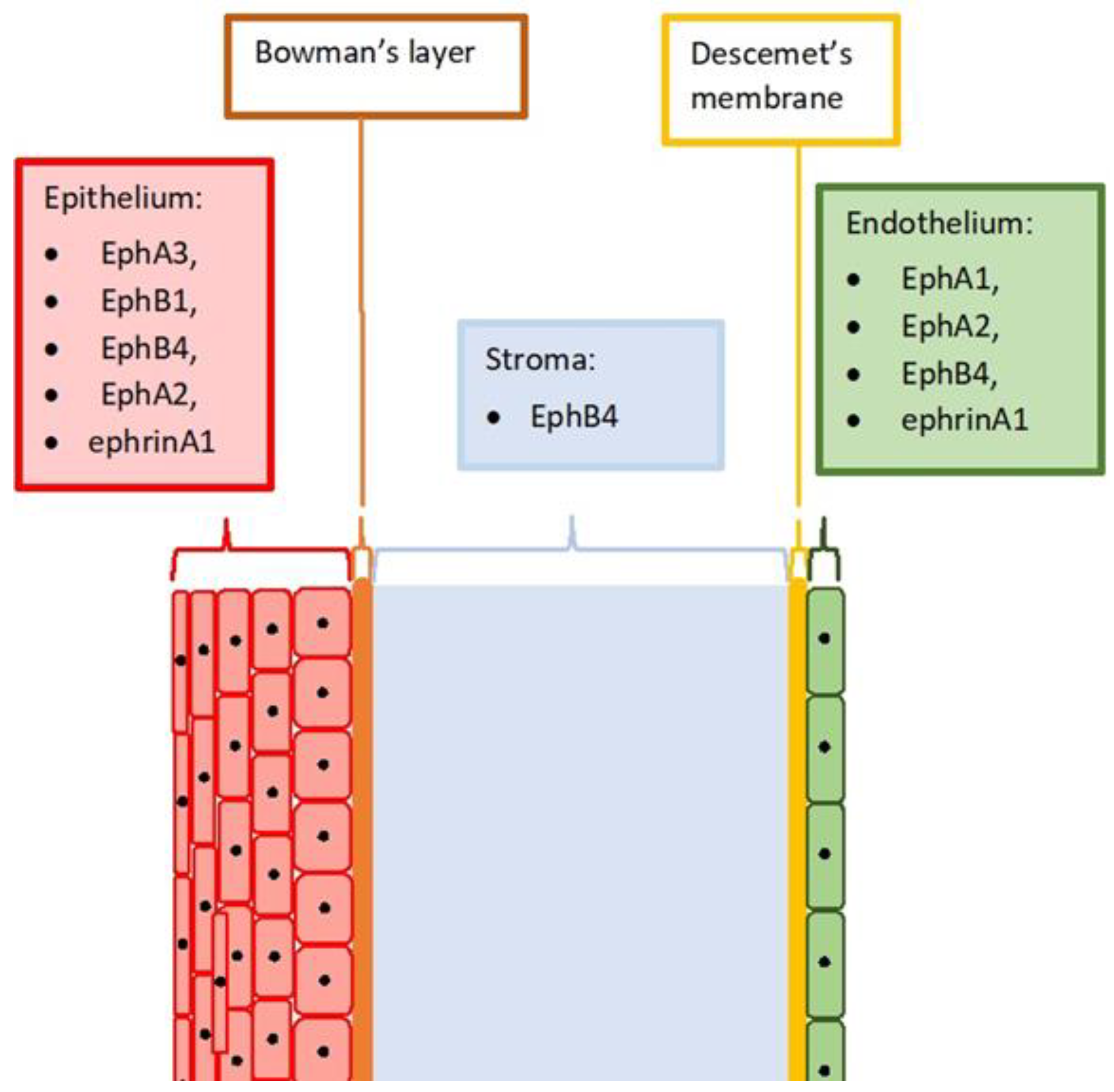
2. Eph and Ephrin Expression in the Healthy Cornea
3. Effect of Eph and Ephrins on the Corneal Epithelium
4. Effect of Eph Ad Ephrins on the Corneal Endothelium
5. Corneal Neovascularization
6. Eph and Ephrin Expression in Neovascularized Corneas
7. Eph and Ephrins in Corneal Neovascularization Models
8. Possible Applications of Eph and Ephrin Signaling System Modifying Drugs in Corneal Diseases
9. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Pasquale, E.B. Eph-Ephrin Bidirectional Signaling in Physiology and Disease. Cell 2008, 133, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Xi, H.-Q.; Wu, X.-S.; Wei, B.; Chen, L. Eph receptors and ephrins as targets for cancer therapy. J. Cell. Mol. Med. 2012, 16, 2894–2909. [Google Scholar] [CrossRef]
- Nievergall, E.; Lackmann, M.; Janes, P.W. Eph-dependent cell-cell adhesion and segregation in development and cancer. Cell. Mol. Life Sci. 2012, 69, 1813–1842. [Google Scholar] [CrossRef]
- Surawska, H.; Ma, P.C.; Salgia, R. The role of ephrins and Eph receptors in cancer. Cytokine Growth Factor Rev. 2004, 15, 419–433. [Google Scholar] [CrossRef]
- Rüegg, C.; Mariotti, A. Vascular integrins: Pleiotropic adhesion and signaling molecules in vascular homeostasis and angiogenesis. Cell. Mol. Life Sci. 2003, 60, 1135–1157. [Google Scholar] [CrossRef]
- Cramer, K.S.; Miko, I.J. Eph-ephrin signaling in nervous system development. F1000Research 2016, 5, 413. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Pasqualini, R.; Lindberg, R.A.; Kain, R.; Freeman, A.L.; Pasquale, E.B. The ephrin-A1 ligand and its receptor, EphA2, are expressed during tumor neovascularization. Oncogene 2000, 19, 6043–6052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez White, B.E.; Getsios, S. Eph receptor and ephrin function in breast, gut, and skin epithelia. Cell Adh. Migr. 2014, 8, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaff, D. Involvement of endothelial ephrin-B2 in adhesion and transmigration of EphB-receptor-expressing monocytes. J. Cell Sci. 2008, 121, 3842–3850. [Google Scholar] [CrossRef] [Green Version]
- Tognolini, M.; Hassan-Mohamed, I.; Giorgio, C.; Zanotti, I.; Lodola, A. Therapeutic perspectives of Eph–ephrin system modulation. Drug Discov. Today 2014, 19, 661–669. [Google Scholar] [CrossRef]
- Barquilla, A.; Pasquale, E.B. Eph receptors and ephrins: Therapeutic opportunities. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 465–487. [Google Scholar] [CrossRef] [Green Version]
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef]
- Patel, S.; Tutchenko, L. The refractive index of the human cornea: A review. Contact Lens Anterior Eye 2019, 42, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Boote, C.; Dennis, S.; Newton, R.H.; Puri, H.; Meek, K.M. Collagen fibrils appear more closely packed in the prepupillary cornea: Optical and biomechanical implications. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2941–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiemke, M.M.; Edelhauser, H.F.; Geroski, D.H. The developing corneal endothelium: Correlation of morphology, hydration and na/k ATPase pump site density. Curr. Eye Res. 1991, 10, 145–156. [Google Scholar] [CrossRef]
- Meek, K.M.; Knupp, C. Corneal structure and transparency. Prog. Retin. Eye Res. 2015, 49, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, T.; Chung, T.Y.; Chang, J.H.; Sayegh, R.; Casanova, F.H.; Azar, D.T. Comparison of EphA receptor tyrosine kinases and ephrinA ligand expression to EphB-ephrinB in vascularized corneas. Cornea 2007, 26, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, C.S.; Flood, W.D.; Koblar, S.A. High EphA3 expressing ophthalmic trigeminal sensory axons are sensitive to ephrin-A5-Fc: Implications for lobe specific axon guidance. Neuroscience 2005, 135, 97–109. [Google Scholar] [CrossRef]
- Walshe, J.; Richardson, N.A.; Al Abdulsalam, N.K.; Stephenson, S.A.; Harkin, D.G. A potential role for Eph receptor signalling during migration of corneal endothelial cells. Exp. Eye Res. 2018, 170, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Hogerheyde, T.A. Evaluation of Eph receptor and ephrin expression within the human cornea and limbus. Exp. Eye Res. 2013, 107, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, N.; Fatima, A.; Peng, H.; Bryar, P.J.; Lavker, R.M.; Getsios, S. EphA2/ephrin-A1 signaling complexes restrict corneal epithelial cell migration. Investig. Ophthalmol. Vis. Sci. 2012, 53, 936–945. [Google Scholar] [CrossRef]
- Lavker, R.M.; Kaplan, N.; Wang, J.; Peng, H. Corneal epithelial biology: Lessons stemming from old to new. Exp. Eye Res. 2020, 198, 108094. [Google Scholar] [CrossRef]
- Zhu, L.; Titone, R.; Robertson, D.M. The impact of hyperglycemia on the corneal epithelium: Molecular mechanisms and insight. Ocul. Surf. 2019, 17, 644–654. [Google Scholar] [CrossRef]
- Shih, K.C.; Lam, K.L.; Tong, L. A systematic review on the impact of diabetes mellitus on the ocular surface. Nutr. Diabetes 2017, 7, e251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Yu FS, X. Impaired epithelial wound healing and EGFR signaling pathways in the corneas of diabetic rats. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3301–3308. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, N. Epha2/ephrin-A1 mediate corneal epithelial cell compartmentalization via ADAM10 regulation of EGFR signaling. Investig. Ophthalmol. Vis. Sci. 2018, 59, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R.M.; Tseng, S.C.; Sun, T.T. Corneal epithelial stem cells at the limbus: Looking at some old problems from a new angle. Exp. Eye Res. 2004, 78, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, Y.; Xie, L.; He, X.; Bai, J. Progress in the pathogenesis of pterygium. Curr. Eye Res. 2013, 38, 1191–1197. [Google Scholar] [CrossRef]
- John-Aryankalayil, M. Microarray and protein analysis of human pterygium. Mol. Vis. 2006, 23, 55–64. [Google Scholar]
- Livezeanu, C.; Crǎiţoiu, M.M.C.; Mǎnescu, R.; Mocanu, C.; Crǎiţoiu, Ş. Angiogenesis in the pathogenesis of pterygium. Rom. J. Morphol. Embryol. 2011, 52, 837–844. [Google Scholar] [PubMed]
- Xue, C.; Chen, Y.; Huang, Z.; Ge, Y.; Wang, H.; Wang, J. EphB4 expression in pterygium is associated with microvessel density. Int. J. Clin. Exp. Med. 2014, 7, 4008–4015. [Google Scholar]
- Xue, C.; Huang, Z.; Wang, J.; Dong, Y.; Zhou, X. EphrinB2 and EphB4 expression in pterygia: New insights and preliminary results. Can. J. Ophthalmol. 2009, 44, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Pleyer, U. The human corneal endothelium: New insights into electrophysiology and ion channels. Prog. Retin. Eye Res. 2007, 26, 359–378. [Google Scholar] [CrossRef]
- Ojima, T. EphrinA1 inhibits vascular endothelial growth factor-induced intracellular signaling and suppresses retinal neovascularization and blood-retinal barrier breakdown. Am. J. Pathol. 2006, 168, 331–339. [Google Scholar] [CrossRef] [Green Version]
- White, A.I.; Sabater, A.L. Current strategies for human corneal endothelial regeneration. Regen. Med. 2019, 14, 257–261. [Google Scholar] [CrossRef]
- Giorgio, C.; Mohamed, I.H.; Flammini, L.; Barocelli, E.; Incerti, M.; Lodola, A.; Tognolini, M. Lithocholic Acid Is an Eph-ephrin Ligand Interfering with Eph-kinase Activation. PLoS ONE 2011, 6, e18128. [Google Scholar] [CrossRef]
- Azar, D.T. Corneal angiogenic privilege: Angiogenic and antiangiogenic factors in corneal avascularity, vasculogenesis, and wound healing (An American Ophthalmological Society Thesis). Trans. Am. Ophthalmol. Soc. 2006, 104, 264–302. [Google Scholar] [PubMed]
- Sharif, Z.; Sharif, W. Corneal neovascularization: Updates on pathophysiology, investigations & management. Rom. J. Ophthalmol. 2019, 63, 15–22. [Google Scholar] [PubMed]
- Abdelfattah, N.S. Clinical correlates of common corneal neovascular diseases: A literature review. Int. J. Ophthalmol. 2015, 8, 182–193. [Google Scholar]
- Lee, P.; Wang, C.C.; Adamis, A.P. Ocular neovascularization: An epidemiologic review. Surv. Ophthalmol. 1998, 43, 245–269. [Google Scholar] [CrossRef]
- Alipour, F.; Khaheshi, S.; Soleimanzadeh, M.; Heidarzadeh, S. Contact lens-related complications: A review. J. Ophthalmic Vis. Res. 2017, 12, 193–204. [Google Scholar]
- Papas, E. Corneal vascularisation and contact lenses. Arch. Soc. Esp. Oftalmol. 2006, 81, 309–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.; Illingworth, C. Treatments for corneal neovascularization: A review. Cornea 2011, 30, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Ellenberg, D. Novel aspects of corneal angiogenic and lymphangiogenic privilege. Prog. Retin. Eye Res. 2010, 29, 208–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brantley-Sieders, D.; Chen, J. Eph receptor tyrosine kinases in angiogenesis: From development to disease. Angiogenesis 2004, 7, 17–28. [Google Scholar] [CrossRef]
- Cheng, N. Blockade of EphA receptor tyrosine kinase activation inhibits vascular endothelial cell growth factor-induced angiogenesis. Mol. Cancer Res. 2002, 1, 2–11. [Google Scholar]
- Kuijper, S.; Turner, C.J.; Adams, R.H. Regulation of Angiogenesis by Eph–Ephrin Interactions. Trends Cardiovasc. Med. 2007, 17, 145–151. [Google Scholar] [CrossRef]
- Kenyon, B.M.; Voest, E.E.; Chen, C.C.; Flynn, E.; Folkman, J.; D’Amato, R.J. A model of Angiogenesis in the Mouse Cornea. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1625–1632. [Google Scholar]
- Maekawa, H. Ephrin-B2 Induces Migration of Endothelial Cells through the Phosphatidylinositol-3 Kinase Pathway and Promotes Angiogenesis in Adult Vasculature. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 2008–2014. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.I.; Asahara, T.; Masuda, H.; Isner, J.M.; Losordo, D.W. Functional ephrin-B2 expression for promotive interaction between arterial and venous vessels in postnatal neovascularization. Circulation 2005, 111, 2210–2218. [Google Scholar] [CrossRef] [Green Version]
- Uyen Huynh-Do TO, D.; Vindis, C.; Liu, H.; Cerretti, D.; McGrew, J.; Enriquez, M.; Chen, J. Ephrin-B1 transduces signals to activate integrin-mediated migration, attachment and angiogenesis. J. Cell Sci. 2002, 115, 3073–3081. [Google Scholar] [CrossRef]
- Pandey, A.; Shao, H.; Marks, R.M.; Polverini, P.J.; Dixit, V.M. Role of B61, the ligand for the Eck receptor tyrosine kinase, in TNF-α-induced angiogenesis. Science 1995, 268, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Lindauer, M.; Hochhaus, A. Dasatinib. Recent Results Cancer Res. 2018, 212, 29–68. [Google Scholar]
- He, S. Soluble EphB4 regulates choroidal endothelial cell function and inhibits laser-induced choroidal neovascularization. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4772–4779. [Google Scholar] [CrossRef]
- Davies, M.H.; Zamora, D.O.; Smith, J.R.; Powers, M.R. Soluble ephrin-B2 mediates apoptosis in retinal neovascularization and in endothelial cells. Microvasc. Res. 2009, 77, 382–386. [Google Scholar] [CrossRef] [Green Version]
- Zamora, D.O.; Davies, M.H.; Planck, S.R.; Rosenbaum, J.T.; Powers, M.R. Soluble forms of EphrinB2 and EphB4 reduce retinal neovascularization in a model of proliferative retinopathy. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2175–2182. [Google Scholar] [CrossRef]
- Ehlken, C. Therapeutic interference with EphrinB2 signalling inhibits oxygen-induced angioproliferative retinopathy. Acta Ophthalmol. 2011, 89, 82–90. [Google Scholar] [CrossRef]
- Brar, M.; Cheng, L.; Yuson, R.; Mojana, F.; Freeman, W.R.; Gill, P.S. Ocular safety profile and intraocular pharmacokinetics of an antagonist of EphB4/EphrinB2 signalling. Br. J. Ophthalmol. 2010, 94, 1668–1673. [Google Scholar]
- Tsang, S.H.; Sharma, T. Drug-induced retinal toxicity. Adv. Exp. Med. Biol. 2018, 1085, 227–232. [Google Scholar]
- Horita, S. Species differences in ocular pharmacokinetics and pharmacological activities of regorafenib and pazopanib eye-drops among rats, rabbits and monkeys. Pharmacol. Res. Perspect. 2019, 7, e00545. [Google Scholar] [CrossRef]
- Kim, Y.C. Gelling hypotonic polymer solution for extended topical drug delivery to the eye. Nat. Biomed. Eng. 2020, 4, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, T. Topical administration of regorafenib eye drops: Phase I dose-escalation study in healthy volunteers. Br. J. Clin. Pharmacol. 2018, 84, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, M.; Zhang, C. Inhibition of angiogenesis by leflunomide via targeting the soluble ephrin-A1/EphA2 system in bladder cancer. Sci. Rep. 2018, 8, 1539. [Google Scholar] [CrossRef] [Green Version]
- Dobrzanski, P. Antiangiogenic and Antitumor Efficacy of EphA2 Receptor Antagonist. Cancer Res. 2004, 64, 910–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janes, P.W.; Vail, M.E.; Gan, H.K.; Scott, A.M. Antibody targeting of eph receptors in cancer. Pharmaceuticals 2020, 13, 88. [Google Scholar] [CrossRef]
- Tang, F.H.F.; Davis, D.; Arap, W.; Pasqualini, R.; Staquicini, F.I. Eph receptors as cancer targets for antibody-based therapy. Adv. Cancer Res. 2020, 147, 303–317. [Google Scholar]



{kind=link}
{kind=link}
| Healthy Cornea | NV Cornea | |
|---|---|---|
| EphA1 | Endothelium | Epithelium |
| EphA2 | Epithelium, endothelium | Epithelium |
| EphA3 | Epithelium | Epithelium, stroma |
| EphB1 | Epithelium | Epithelium, stroma |
| EphB4 | Epithelium, stroma, endothelium | Epithelium, stroma |
| ephrinA1 | Epithelium, endothelium | Epithelium, stroma |
| ephrinA2 | Not shown | Epithelium, stroma |
| ephrinB1 | Not shown | Epithelium, stroma |
| ephrinB2 | Not shown | Epithelium, stroma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczmarek, R.; Zimmer, K.; Gajdzis, P.; Gajdzis, M. The Role of Eph Receptors and Ephrins in Corneal Physiology and Diseases. Int. J. Mol. Sci. 2021, 22, 4567. https://doi.org/10.3390/ijms22094567
Kaczmarek R, Zimmer K, Gajdzis P, Gajdzis M. The Role of Eph Receptors and Ephrins in Corneal Physiology and Diseases. International Journal of Molecular Sciences. 2021; 22(9):4567. https://doi.org/10.3390/ijms22094567
Chicago/Turabian StyleKaczmarek, Radoslaw, Katarzyna Zimmer, Pawel Gajdzis, and Malgorzata Gajdzis. 2021. "The Role of Eph Receptors and Ephrins in Corneal Physiology and Diseases" International Journal of Molecular Sciences 22, no. 9: 4567. https://doi.org/10.3390/ijms22094567
APA StyleKaczmarek, R., Zimmer, K., Gajdzis, P., & Gajdzis, M. (2021). The Role of Eph Receptors and Ephrins in Corneal Physiology and Diseases. International Journal of Molecular Sciences, 22(9), 4567. https://doi.org/10.3390/ijms22094567