
Thermal Analysis of Stomatal Response under Salinity and High Light

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
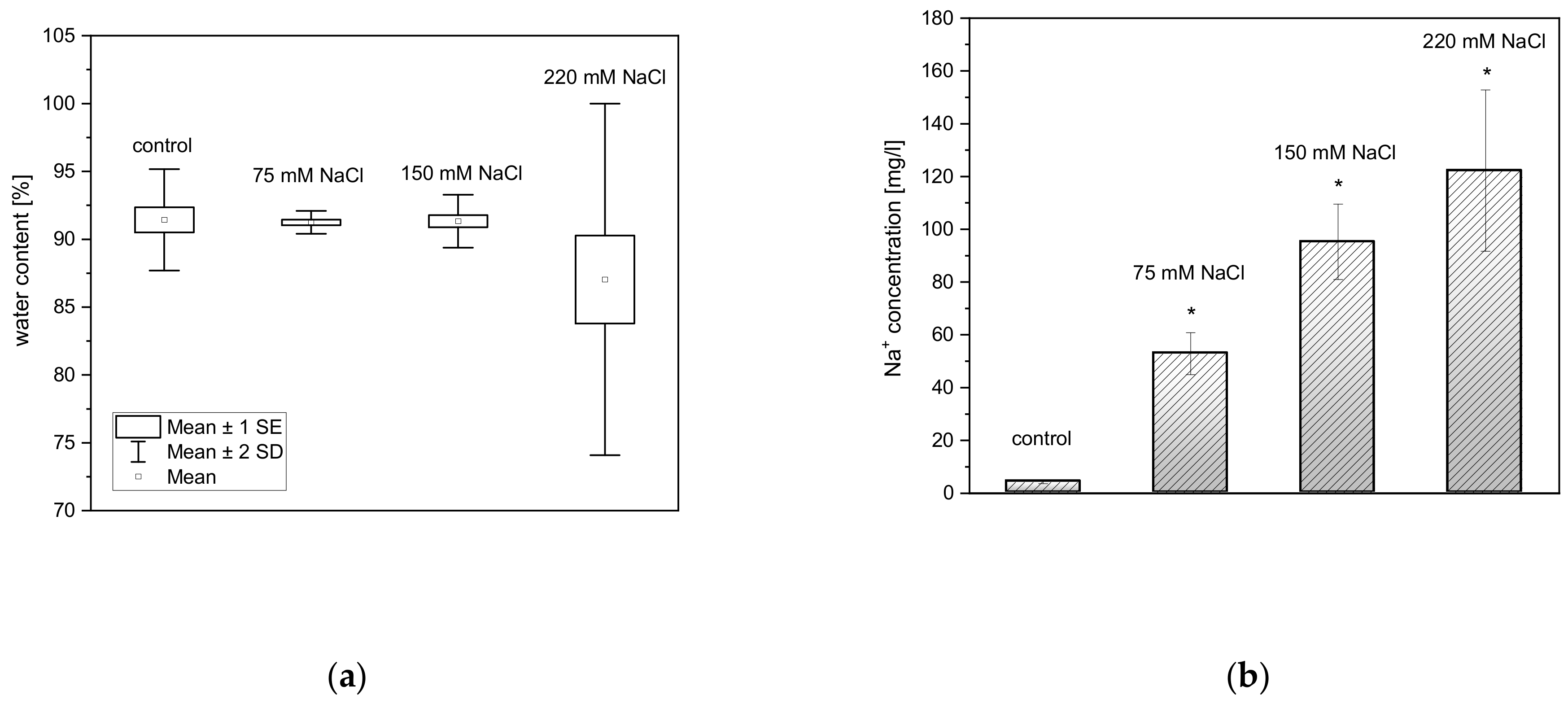
2.1. Effect of Salinity on Dry Mass, Water Content, and Na+ Uptake by Plants
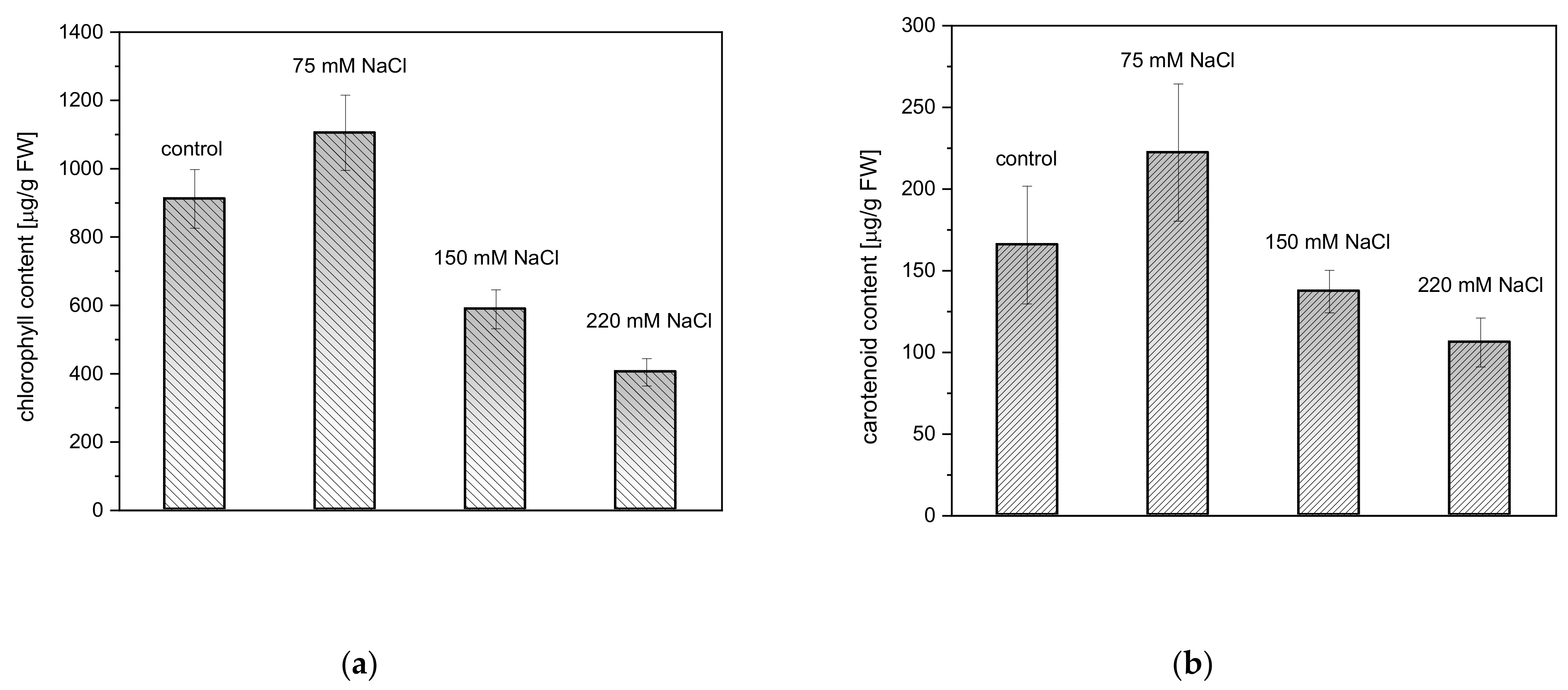
2.2. Effect of Salinity on Photosynthetic Pigment Content
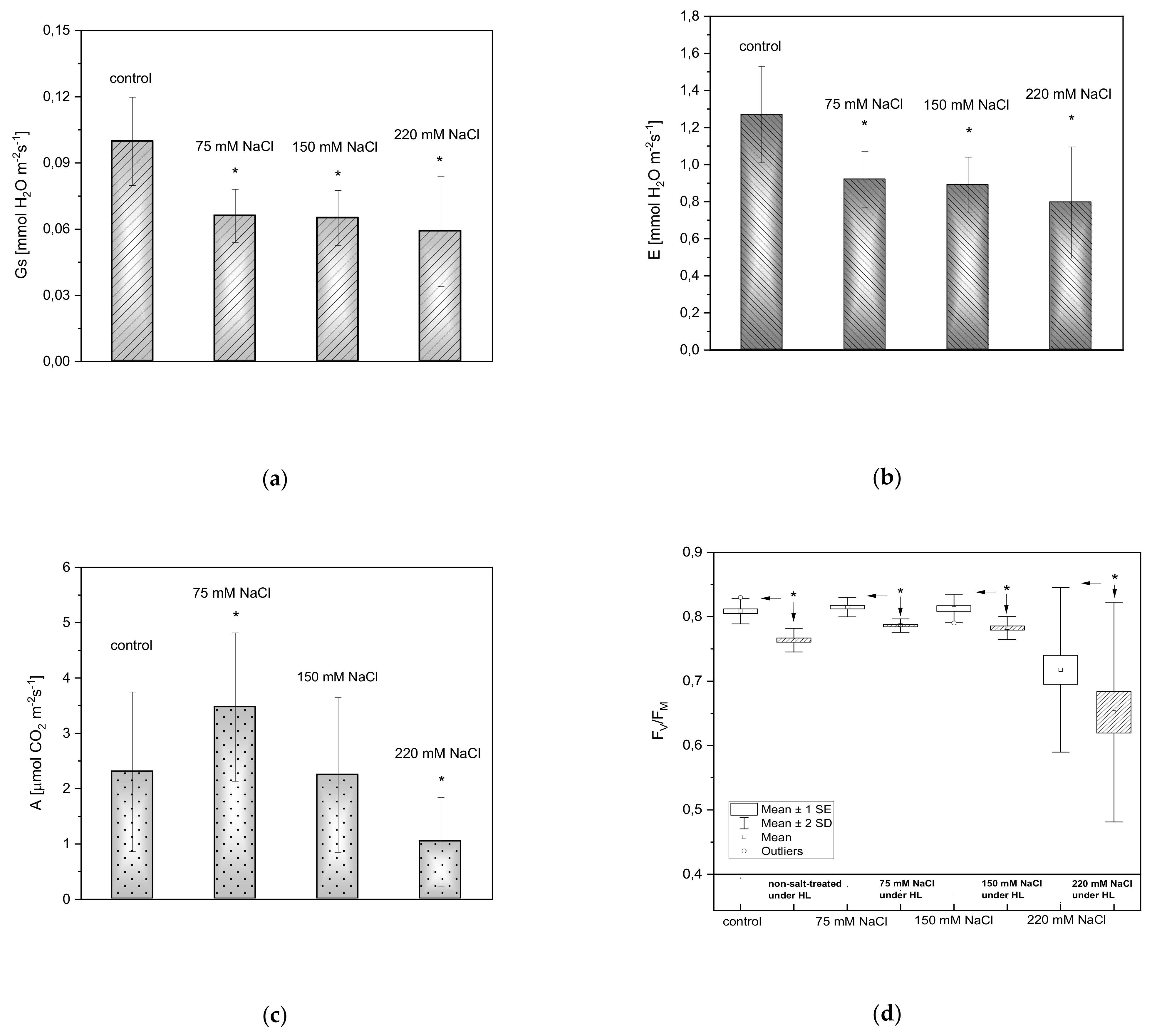
2.3. Gas Exchange and Fluorescence Measurements
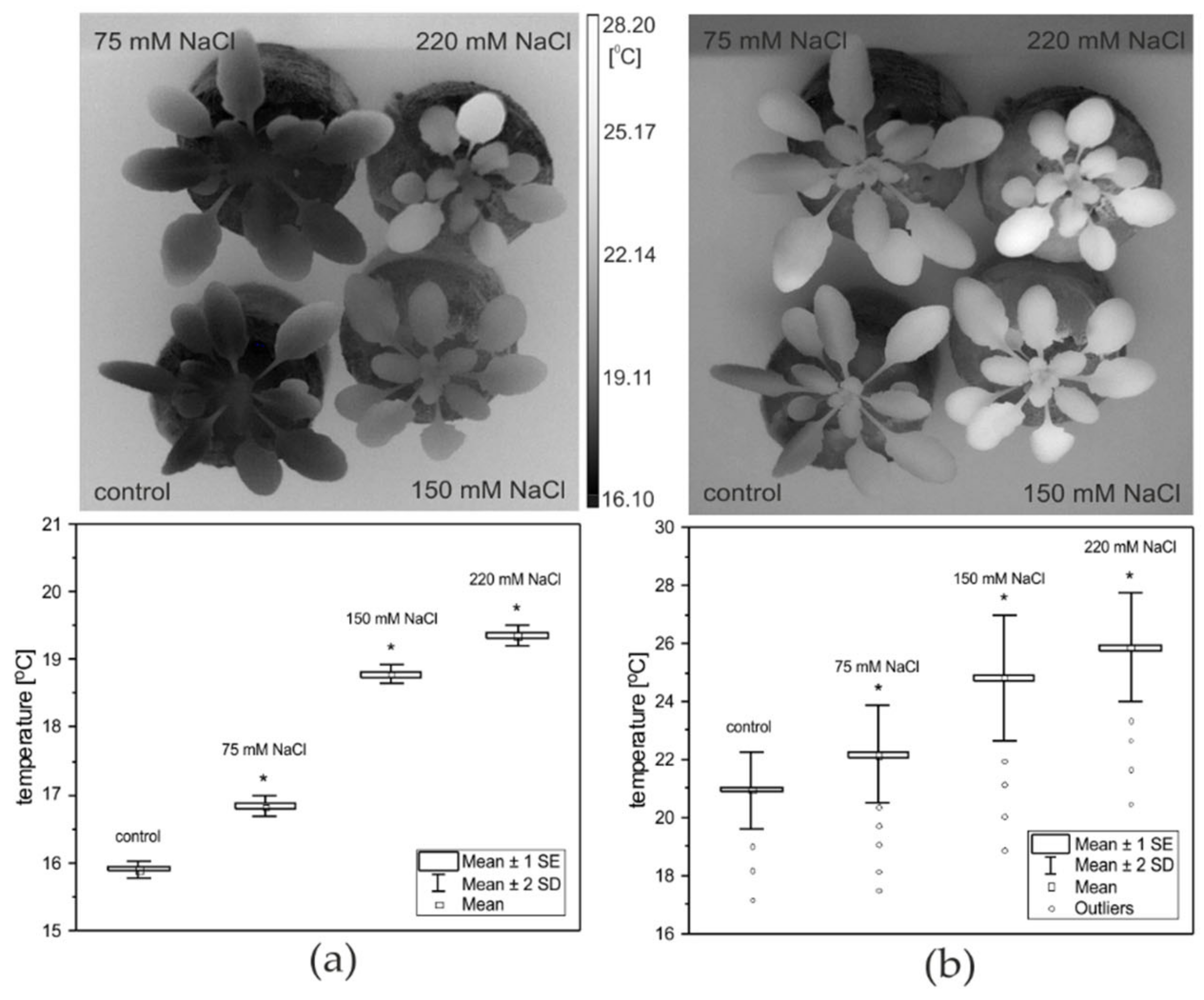
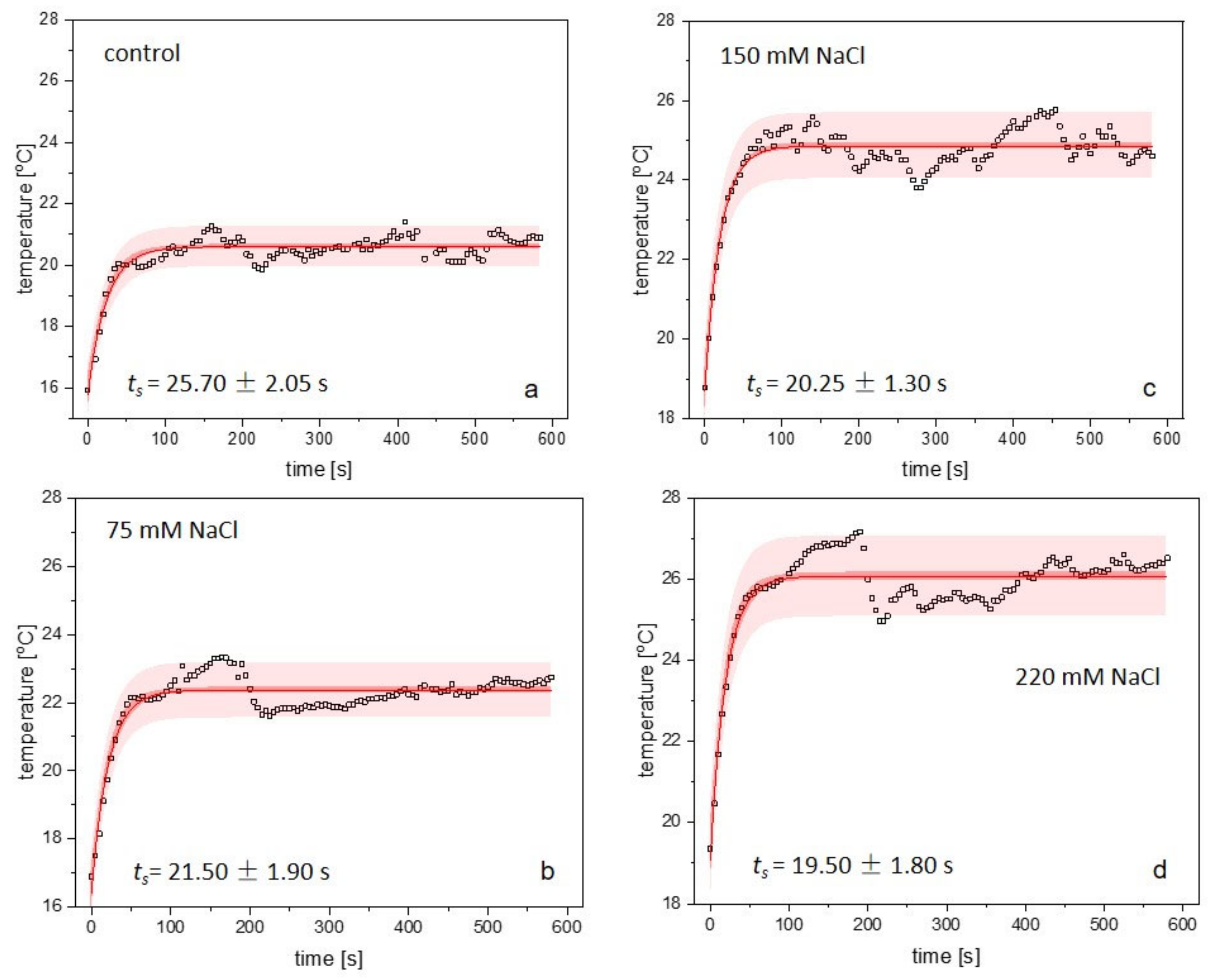
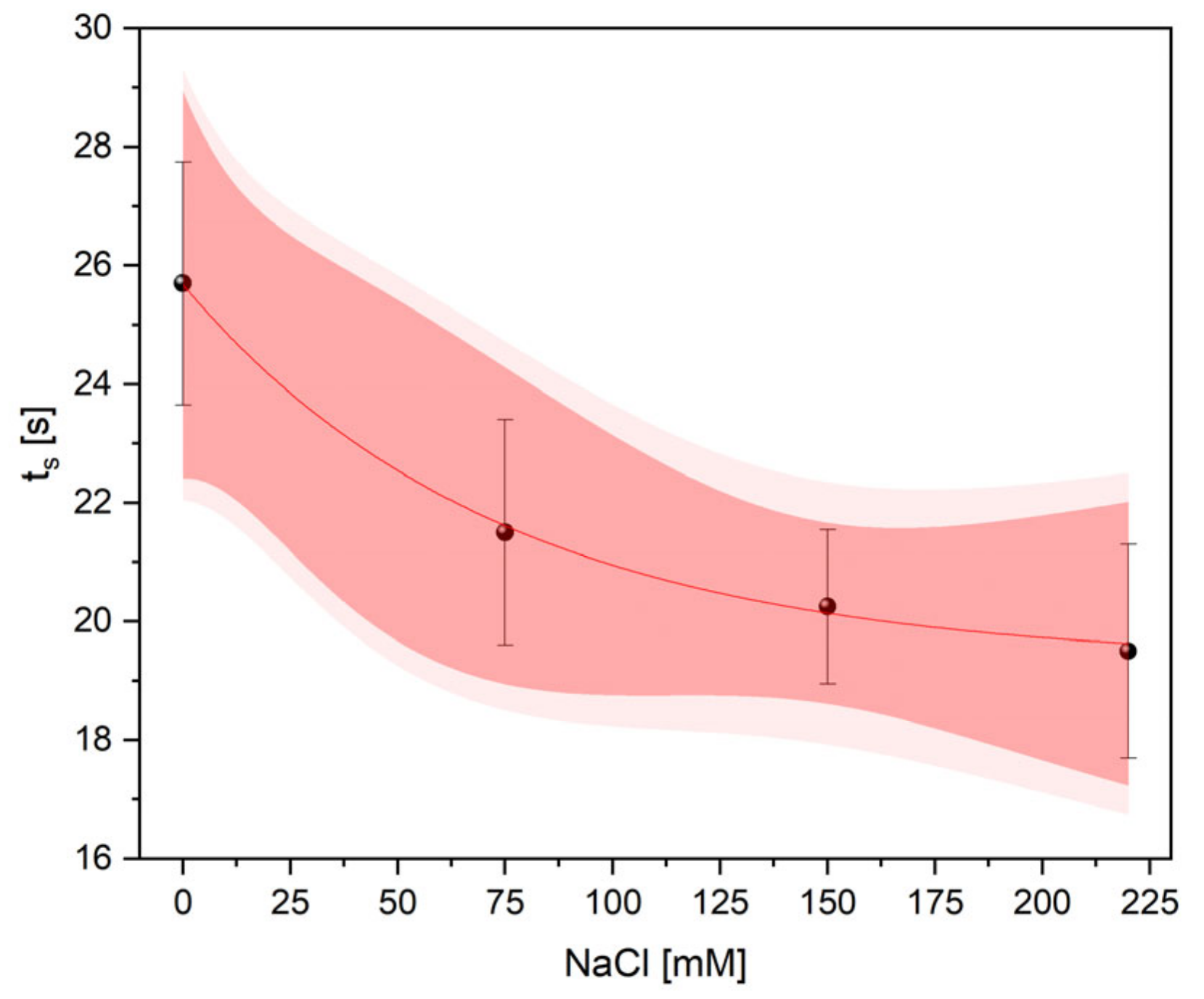
2.4. Thermometric Measurements
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Measurement of Gas Exchange
4.3. Measurement of Infrared Imaging and HL Treatment
4.4. Sodium Content Analysis
4.5. Leaf Water Content
4.6. Measurements of Photosynthetic Pigments
4.7. Measurements of Maximum Quantum Yield of Photosystem II (Maximal Efficiency of PSII Photochemistry)
4.8. Statistical Analysis and Data Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| A. thaliana | Arabidopsis thaliana |
| Chl a | chlorophyll a |
| Car | Carotenoid |
| F0(FM) | the minimum (maximum) chlorophyll a fluorescence in the dark-adapted state |
| HL | high light |
| ICP | inductively coupled plasma emission spectrometry |
| IR | Infrared |
| LED | light-emitting diode |
| LL | low light |
| PAR | photosynthetically active radiation |
| PSII | photosystem II |
| ROS | reactive oxygen species |
| TG | Thermogram |
References
- Jones, H.G.; Archer, N.; Rotenberg, E.; Casa, R. Radiation measurement for plant ecophysiology. J. Exp. Bot. 2003, 54, 879–889. [Google Scholar] [CrossRef]
- Bison, P.; Grinzato, E. Fast estimate of solid materials thermal conductivity by IR thermography. Qirt J. 2010, 7, 17–34. [Google Scholar] [CrossRef]
- Jafari, M.; Minaei, S.; Safaie, N. Detection of pre-symptomatic rose powdery-mildew and gray-mold diseases based on thermal vision. Infrared Phys. Technol. 2017, 85, 170–183. [Google Scholar] [CrossRef]
- Sagan, V.; Maimaitijiang, M.; Sidike, P.; Eblimit, K.; Peterson, K.; Hartling, S.; Esposito, F.; Khanal, K.; Newcomb, M.; Pauli, D.; et al. UAV-based high resolution thermal imaging for vegetation monitoring, and plant phenotyping using ICI 8640 P, FLIR Vue Pro R 640, and thermoMap cameras. Remote Sens. 2019, 11, 330. [Google Scholar] [CrossRef] [Green Version]
- Sagakami, T.; Kubo, S. Application of pulse heating thermography and lock-in thermography to quantitative nondestructive evaluations. Infrared Phys. Technol. 2002, 43, 211–218. [Google Scholar] [CrossRef]
- Tanner, C.B. Plant temperatures. Agron. J. 1963, 55, 201–211. [Google Scholar] [CrossRef]
- Kestin, J.; Sengers, J.V.; Parsi, B.K.; Levelt Sengers, J.M.H. Thermophysical properties of fluid H2O. J. Phys. Chem. Ref. Data 1984, 13. [Google Scholar] [CrossRef]
- Zábranský, M.; Kolská, Z.; Růžička, N.J.; Domalski, E.S. Heat capacity of liquids: Critical review and recommended values. Supplement II. J. Phys. Chem. Ref. Data 2010, 39, 033108. [Google Scholar] [CrossRef]
- Kana, R.; Vass, I. Thermoimaging as a tool for studying light-induced heating of leaves. Correlation of heat dissipation with the efficiency of photosystem II photochemistry and non-photochemical quenching. Environ. Exp. Bot. 2008, 64, 90–96. [Google Scholar] [CrossRef]
- Jones, H.G.; Stoll, M.; Santos, T.; de Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 54, 879–889. [Google Scholar] [CrossRef]
- Leinonen, I.; Grant, O.M.; Tagliavia, C.P.P.; Chaves, M.M.; Jones, H.G. Estimating stomata conductance with thermal imaginery. Plant Cell Environ. 2006, 29, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Alchanatis, V.; Meron, M.; Saranga, Y.; Tsipris, J. Estimation of leaf water potential by thermal imagery and spatial analysis. J. Exp. Bot. 2005, 56, 1843–1852. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G. Application of thermal imaging and infrared sensing in plant physiology and ecophysiology. Adv. Bot. Res. 2004, 41, 107–163. [Google Scholar] [CrossRef]
- Raza, S.A.; Prince, G.; Clarkson, J.P.; Rajpoot, N.M. Automatic detection of diseased tomato plants using thermal and stereo visible light images. PLoS ONE 2015, 10, e0123262. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Chen, H.; Ciechanowska, I.; Spaner, D. Application of infrared thermal imaging for the rapid diagnosis of crop disease. IFAC-PapersOnLine 2018, 51, 424–430. [Google Scholar] [CrossRef]
- Nilssona, H.E. Hand-held radiometry and IR- thermography of plant diseases in field plot experiments. Int. J. Remote Sens. 1991, 12, 545–557. [Google Scholar] [CrossRef]
- Wisniewski, M.; Neuner, G.; Gusta, L. The use of high-resolution infrared thermography (HRIT) for the study of ice nucleation and ice propagation in plants. J. Vis. Exp. 2015, 99, 1–11. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Zhang, R.X.; Peng, K.; Tagliavia, C.; Li, S.; Xue, S.; Liu, A.; Hu, H.; Zhang, J.; Hubbard, K.E.; et al. The BIG protein distinguishes the process of CO2—Induced stomatal closure from the inhibition of stomatal opening by CO2. New Phytol. 2018, 218, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Merlot, S.; Mustilli, A.C.; Genty, B.; North, H.; Lefebvre, V.; Sotta, B.; Vavasseur, A.; Giraudat, J. Use of infrared thermal imaging to isolate Arabidopsis mutants defective in stomatal regulation. Plant J. 2002, 30, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Lamprecht, I. Flower ovens: Thermal investigations on heat producing plants. Thermochim. Acta 2002, 391, 107–118. [Google Scholar] [CrossRef]
- Hairmansis, A.; Berger, B.; Tester, M.; Roy, S.J. Image-based phenotyping for non-destructive screening of different salinity tolerance traits in rice. Rice 2014, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Munns, R.; James, R.A.; Sirault, X.R.R.; Furbank, R.T.; Jones, H.G. New phenotyping methods for screening wheat and barley for beneficial responses to water deficit. J. Exp. Bot. 2010, 61, 3499–3507. [Google Scholar] [CrossRef] [Green Version]
- James, R.A.; Sirault, X.R.R. Infrared thermography in plant phenotyping for salinity tolerance. Methods Mol. Biol. 2012, 913, 173–189. [Google Scholar] [CrossRef]
- Urrestarazu, M. Infrared thermography used to diagnose the effects of salinity in a soilless culture. Quant. InfraRed Therm. J. 2013, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sirault, X.R.R.; James, R.A.; Furbank, R.T. A new screening method for osmotic component of salinity tolerance in cereals using infrared thermography. Funct. Plant Biol. 2009, 36, 970–997. [Google Scholar] [CrossRef]
- Furbank, R.T.; Tester, M. Phenomics- technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Isayenkov, S.V. Physiological and molecular aspects of salt stress in plants. Cytol. Genet. 2012, 46, 302–318. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Ali, Q. Relative membrane permeability and activities of some antioxidant enzymes as the key determinants of salt tolerance in canola (Brassica napus L.). Environ. Exp. Bot. 2008, 63, 266–273. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanism of salinity tolerance. Ann. Rev. Plant Physiol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugnoli, E.; Lauteri, M. Effects of salinity on stomatal conductance, photosynthetic capacity and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3Non-Halophytes. Plant Physiol. 1991, 95, 628–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.; Gupta, N.K.; Gupta, S.; Hasegawa, H. Effect of NaCl salinity on photosynthetic rate, transpiration rate, and oxidative stress tolerance in contrasting wheat genotypes. Photosynthetica 2005, 43, 609–613. [Google Scholar] [CrossRef]
- Suárez, N. Effects of short-and long-term salinity on leaf water relations, gas exchange, and growth in Ipomoea pes-caprae. Flora 2011, 206, 267–275. [Google Scholar] [CrossRef]
- Chen, T.W.; Stutzel, H.; Kahlen, K. High light aggravates functional limitations of cucumber canopy photosynthesis under salinity. Ann. Bot. 2018, 121, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Juneau, P.; Barnett, A.; Meleder, V.; Dupuy, C.; Lavaud, J. Combined effect of high light and high salinity on the regulation of photosynthesis in three diatom species belonging to the main growth forms of intertidal flat inhabiting microphytobenthos. J. Exp. Mar. Biol. Ecol. 2015, 463, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.M.; Zhang, J.H. Effects of salt stress on PSII function and photoinhibition in the cyanobacterium Spirulina platensis. J. Plant Physiol. 1999, 155, 740–745. [Google Scholar] [CrossRef]
- Murata, N.; Takahshi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta 2007, 1767, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, T.; Krieger-Liszkay, A. Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Nishiyama, Y.; Miyairi, S.; Yamamoto, H.; Inagaki, N.; Kanesaki, Y.; Murata, N. Salt stress inhibits the repair of photodamaged photosystem II by suppressing the transcription and translation of psbA genes in Synechocystis. Plant Physiol. 2002, 130, 1443–1453. [Google Scholar] [CrossRef] [Green Version]
- Kummerlen, B.; Dauwe, S.; Schmundt, D.; Schurr, U. Thermography to measure water relations of plant leaves. In Handbook of Computer Vision and Applications.; Academic Press: London, UK, 1999; Volume 3, pp. 763–781. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Yepes-Molina, L.; Bárzana, G.; Carvajal, M. Controversial regulation of gene expression and protein transduction of aquaporins under drought and salinity stress. Plants 2020, 9, 1662. [Google Scholar] [CrossRef]
- Karpinska, B.; Wingsle, G.; Karpinski, S. Antagonistic effects of hydrogen peroxide and glutathione on acclimation to excess excitation energy in Arabidopsis. IUBMB Life 2000, 50, 21–26. [Google Scholar] [CrossRef]
- Kostaki, K.I.; Coupel-Ledru, A.; Bonnell, V.C.; Gustavsson, M.; Sun, P.; McLaughlin, F.J.; Fraser, D.P.; McLachlan, D.H.; Hetherington, A.M.; Dodd, A.N.; et al. Guard cells integrate light and temperature signals to control stomatal aperture. Plant Physiol. 2020, 182, 1404–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant. Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- James, R.A.; von Caemmerer, S.; Condon, A.G.; Zwart, A.B.; Munns, R. Genetic variation in tolerance to the osmotic stress component of salinity stress in durum wheat. Funct. Plant Biol. 2008, 35, 111–123. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Campillo, A.; Jimenez, A.; Alarcon, J.J.; Sevilla, F. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol. 1999, 141, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Liu, Z.J. Effect of NaCl on growth, morphology, and camptothecin accumulation in Camptotheca acuminata seedlings. Can. J. Plant Sci. 2003, 83, 931–938. [Google Scholar] [CrossRef]
- Abdol-Qados, A.M.S. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Davy, A.J.; Fernández-Muñoz, F.; Castellanos, E.M.; Luque, T.; Figueroa, M.E. Growth and photosynthetic responses to salinity of the salt-marsh shrub Atriplex portulacoides. Ann. Bot. 2007, 100, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepelkova, H.; Yocum, C.F. Current status of the role of Cl− ion in the oxygen-evolving complex. Photosynth. Res. 2007, 93, 111–121. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Mattson, M.P. How does hormesis impact biology, toxicology, and medicine? NPJ Aging Mech. Dis. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Öquist, G.; Chow, W.S.; Anderson, J.M. Photoinhibition of photosynthesis represents a mechanism for the long-term regulation of photosystem II. Planta 1992, 186, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Pliego-Cortésa, H.; Bedouxb, G.; Boulhob, R.; Taupinb, L.; Freile-Pelegrína, Y.; Bourgougnonb, N.; Robledoa, D. Stress tolerance and photoadaptation to solar radiation in Rhodymenia pseudopalmata (Rhodophyta) through mycosporine-like amino acids, phenolic compounds, and pigments in an Integrated Multi-Trophic Aquaculture system. Algal Res. 2019, 41, 1–11. [Google Scholar] [CrossRef]
- Gao, Y.; Zheng, W.; Zhang, C.; Zhang, L.; Xu, K. High temperature and high light intensity induced photoinhibition of bayberry (Myrica rubra Sieb. et Zucc.) by disruption of D1 turnover in photosystem II. Sci. Hortic. 2019, 248, 132–137. [Google Scholar] [CrossRef]
- Gerganova, M.T.; Faik, A.K.; Velitchkova, M.Y. Acquired tolerance of the photosynthetic apparatus to photoinhibition as a result of growing Solanum lycopersicum at moderately higher temperature and light intensity. Funct. Plant. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Somersalo, S.; Zumbusch, E.; Weyers, B.; Laasch, H. On the mechanism of photoinhibition in chloroplasts. Relationship between changes in fluorescence and activity of Photosystem II. J. Plant Physiol. 1990, 136, 472–479. [Google Scholar] [CrossRef]
- Critchley, C. D1 protein turnover: Response to photodamage or regulatory mechanism? In Photoinhibition of Photosynthesis; Baker, N.R., Jr., Bowyer, J.R., Eds.; Bios Scientific: Oxford, UK, 1994; pp. 195–203. [Google Scholar]
- Gerard, J.; Kluitenberg, J.; Biggar, W. Canopy temperature as a measure of salinity stress on sorghum. Irrig. Sci. 1992, 13, 115–121. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.; Lawson, T. Dynamic leaf energy balance: Deriving stomatal conductance from thermal imaging in a dynamic environment. J. Exp. Bot. 2019, 70, 2839–2855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orzechowska, A.; Trtílek, M.; Niewiadomska, E. A thermographic study of plant response to excessive light. Acta Phys. Pol. A 2021, 139, 257–260. [Google Scholar] [CrossRef]
- Orzechowska, A.; Trtílek, M.; Tokarz, K.; Rozpądek, P. A study of light-induced stomatal response in Arabidopsis using thermal imaging. Biochem. Biophys. Res. Commun. 2020, 533, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Bajons, P.; Klinger, G.; Schlosser, V. Determination of stomatal conductance by means of infrared termography. Infrared Phys. Technol. 2005, 46, 429–439. [Google Scholar] [CrossRef]
- Weyers, J.; Lawson, T. Heterogenity in stomatal characteristics. Adv. Bot. Res. 1997, 26, 317–352. [Google Scholar] [CrossRef]
- Lawson, T.; Weyers, J. Spatial and temporal variation in gas exchange over the lower surface of Phaselous vulgaris L. primary leaves. J. Exp. Bot. 1999, 50, 1381–1391. [Google Scholar] [CrossRef]
- Buckley, T.N.; Mott, K.A. Stomatal responses to non—Local changes in PFD: Evidence for long—Distance hydraulic interactions. Plant Cell Environ. 2001, 23, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.-I.; Lee, B.-R.; Chae, M.-J.; Lee, E.-J.; Lee, T.-G.; Jung, G.-B.; Kim, M.-S.; Lee, J. Ascorbate-Mediated Modulation of Cadmium Stress Responses: Reactive Oxygen Species and Redox Status in Brassica napus. Front. Plant Sci. 2020, 11, 1823. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzym. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Cruz, A.J.; Savage, L.J.; Zegarac, R.; Hall, C.C.; Satoh-Cruz, M.; Davis, G.A.; Kovac, W.K.; Chen, J.; Kramer, D.M. Dynamic environmental photosynthetic imaging reveals emergent phenotypes. Cell Syst. 2016, 2, 365–377. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orzechowska, A.; Trtílek, M.; Tokarz, K.M.; Szymańska, R.; Niewiadomska, E.; Rozpądek, P.; Wątor, K. Thermal Analysis of Stomatal Response under Salinity and High Light. Int. J. Mol. Sci. 2021, 22, 4663. https://doi.org/10.3390/ijms22094663
Orzechowska A, Trtílek M, Tokarz KM, Szymańska R, Niewiadomska E, Rozpądek P, Wątor K. Thermal Analysis of Stomatal Response under Salinity and High Light. International Journal of Molecular Sciences. 2021; 22(9):4663. https://doi.org/10.3390/ijms22094663
Chicago/Turabian StyleOrzechowska, Aleksandra, Martin Trtílek, Krzysztof Michał Tokarz, Renata Szymańska, Ewa Niewiadomska, Piotr Rozpądek, and Katarzyna Wątor. 2021. "Thermal Analysis of Stomatal Response under Salinity and High Light" International Journal of Molecular Sciences 22, no. 9: 4663. https://doi.org/10.3390/ijms22094663
APA StyleOrzechowska, A., Trtílek, M., Tokarz, K. M., Szymańska, R., Niewiadomska, E., Rozpądek, P., & Wątor, K. (2021). Thermal Analysis of Stomatal Response under Salinity and High Light. International Journal of Molecular Sciences, 22(9), 4663. https://doi.org/10.3390/ijms22094663