
Skeletal Muscle Proteomic Profile Revealed Gender-Related Metabolic Responses in a Diet-Induced Obesity Animal Model

 , , , , , , , ,
, , , , , , , ,  and
and
Abstract
:1. Introduction
2. Results
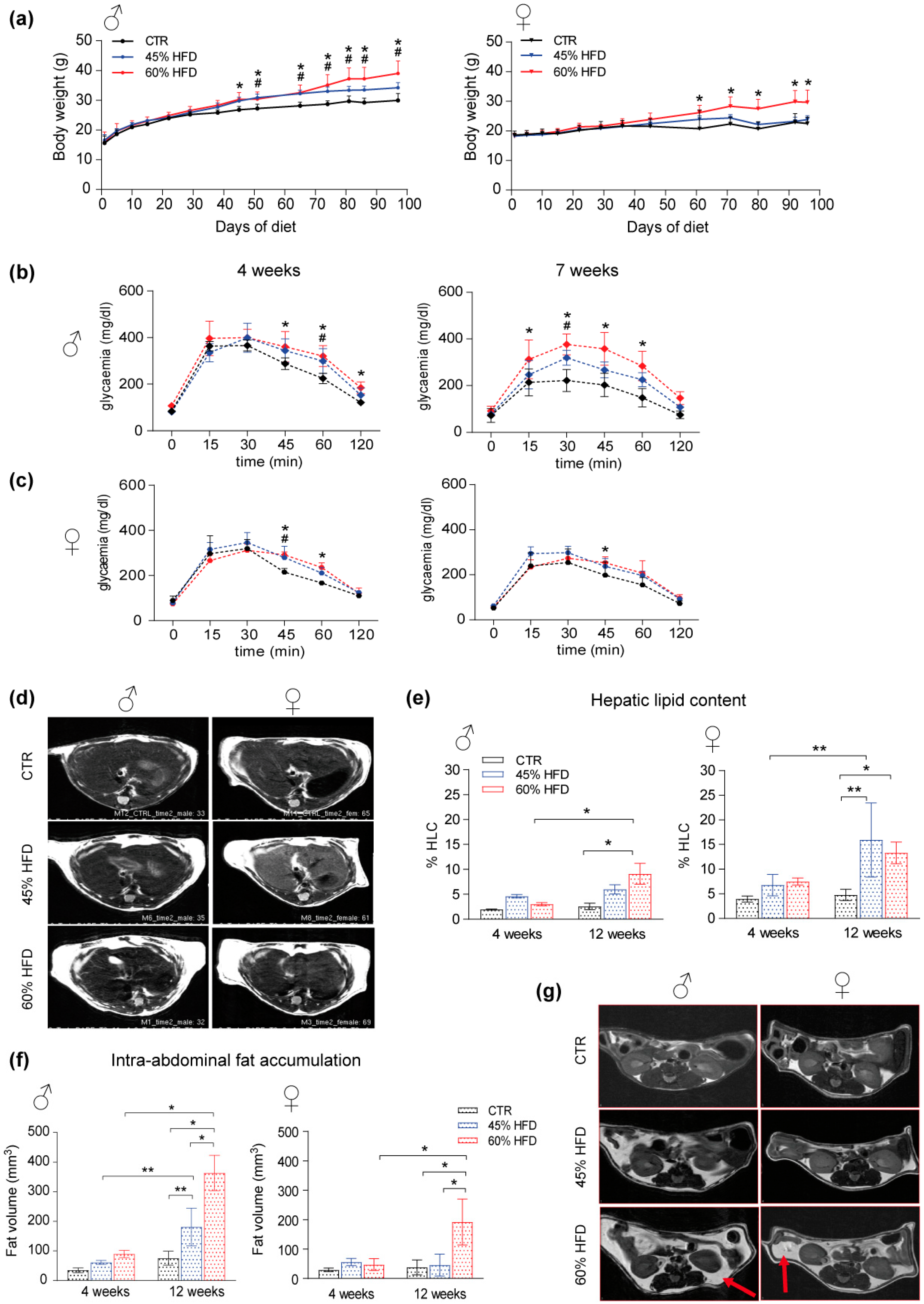
2.1. High-Fat Regimen Effects and Gender Response
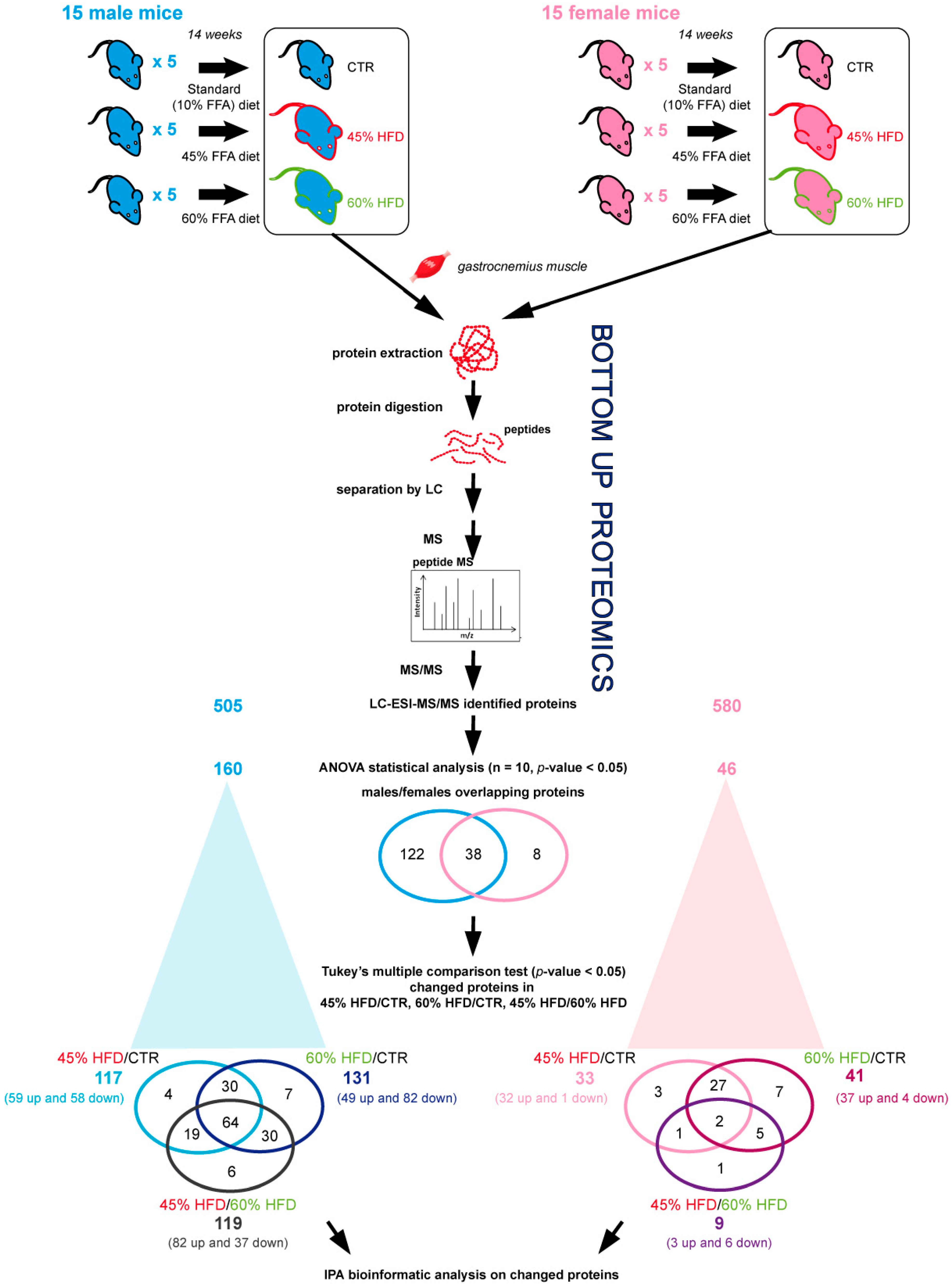
2.2. Proteomic Profiles of Skeletal Muscle in High-Fat Diet
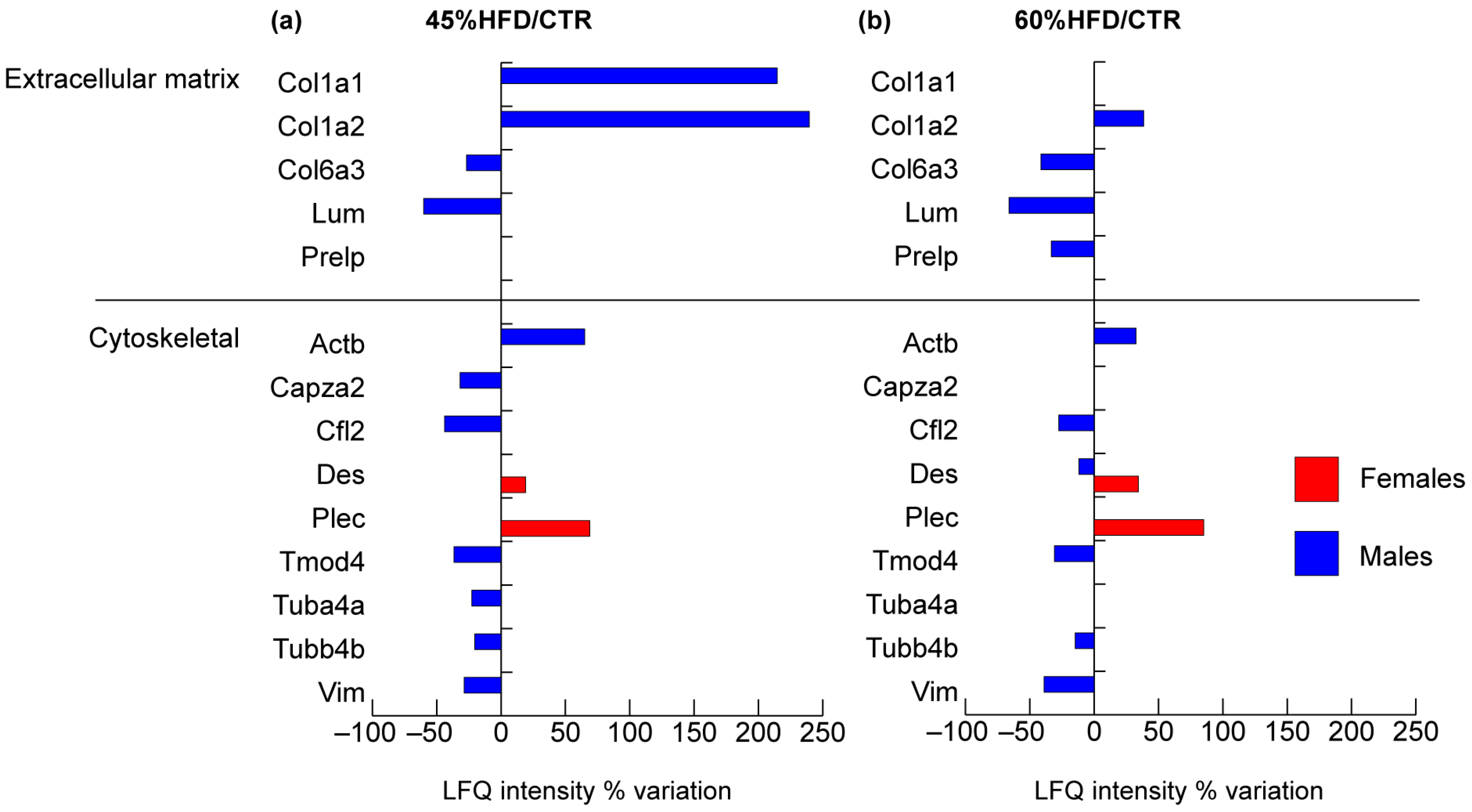
2.2.1. Structural Proteins
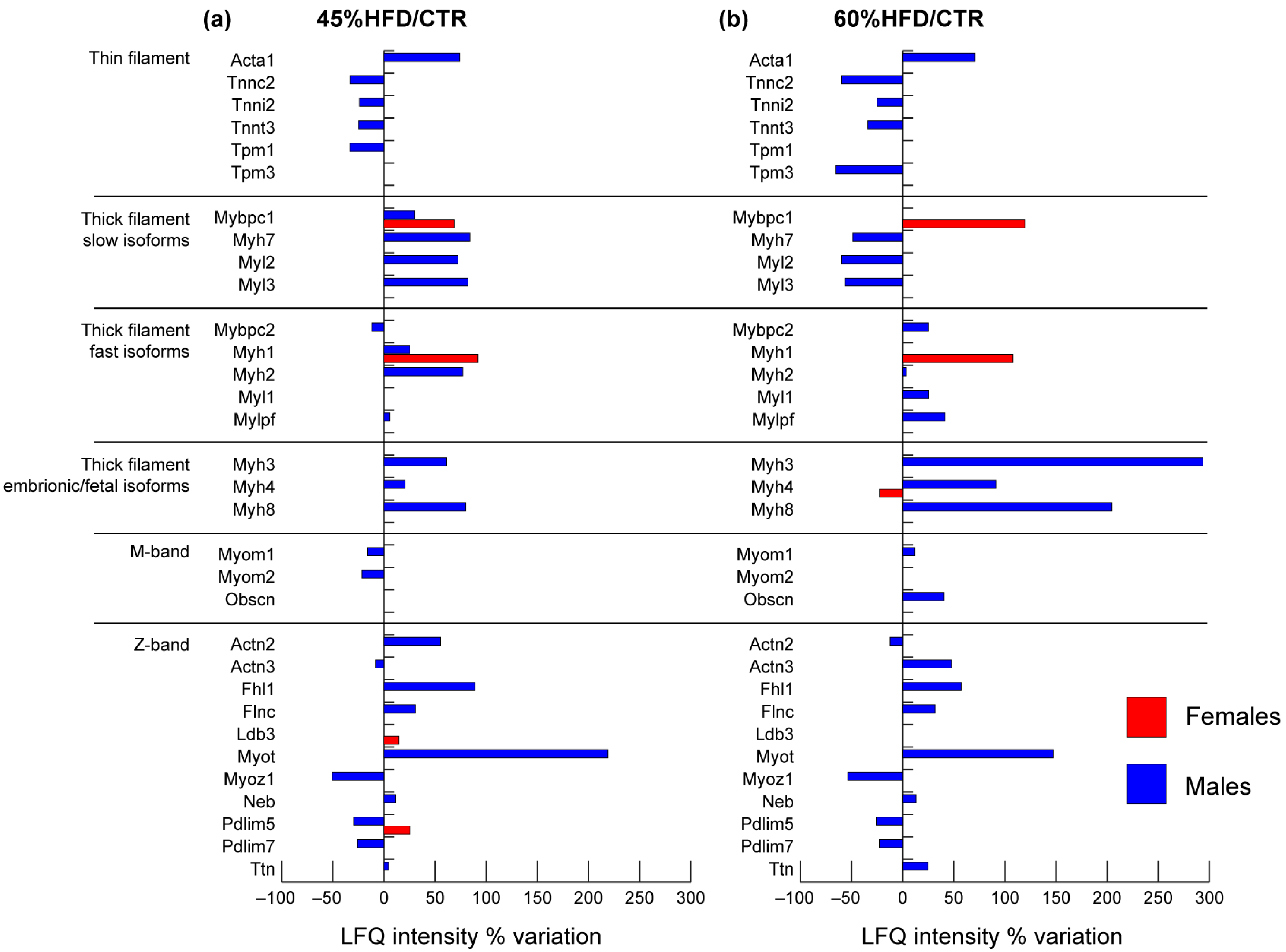
2.2.2. Contractile Proteins
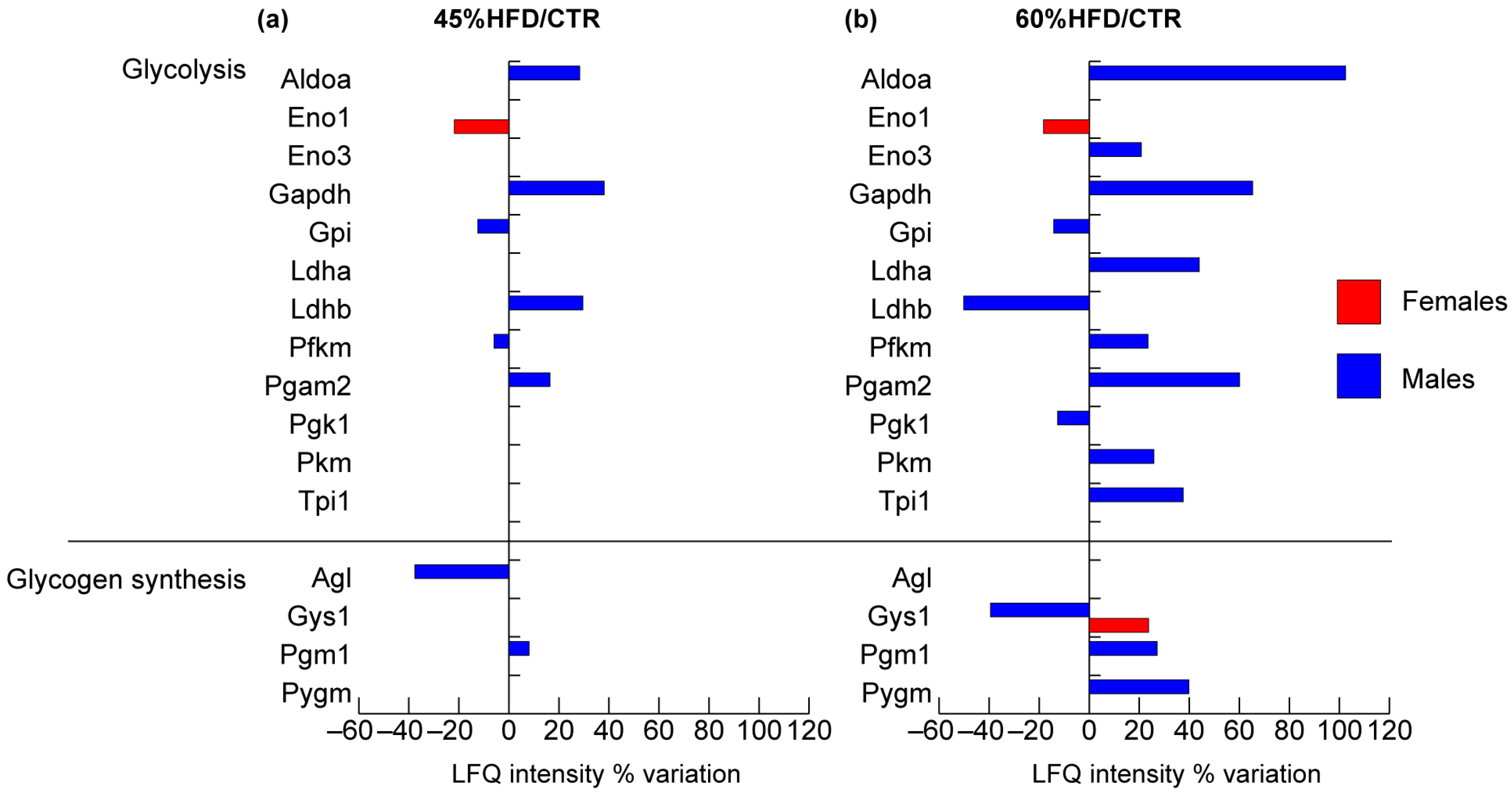
2.2.3. Glucose and Glycogen Metabolism
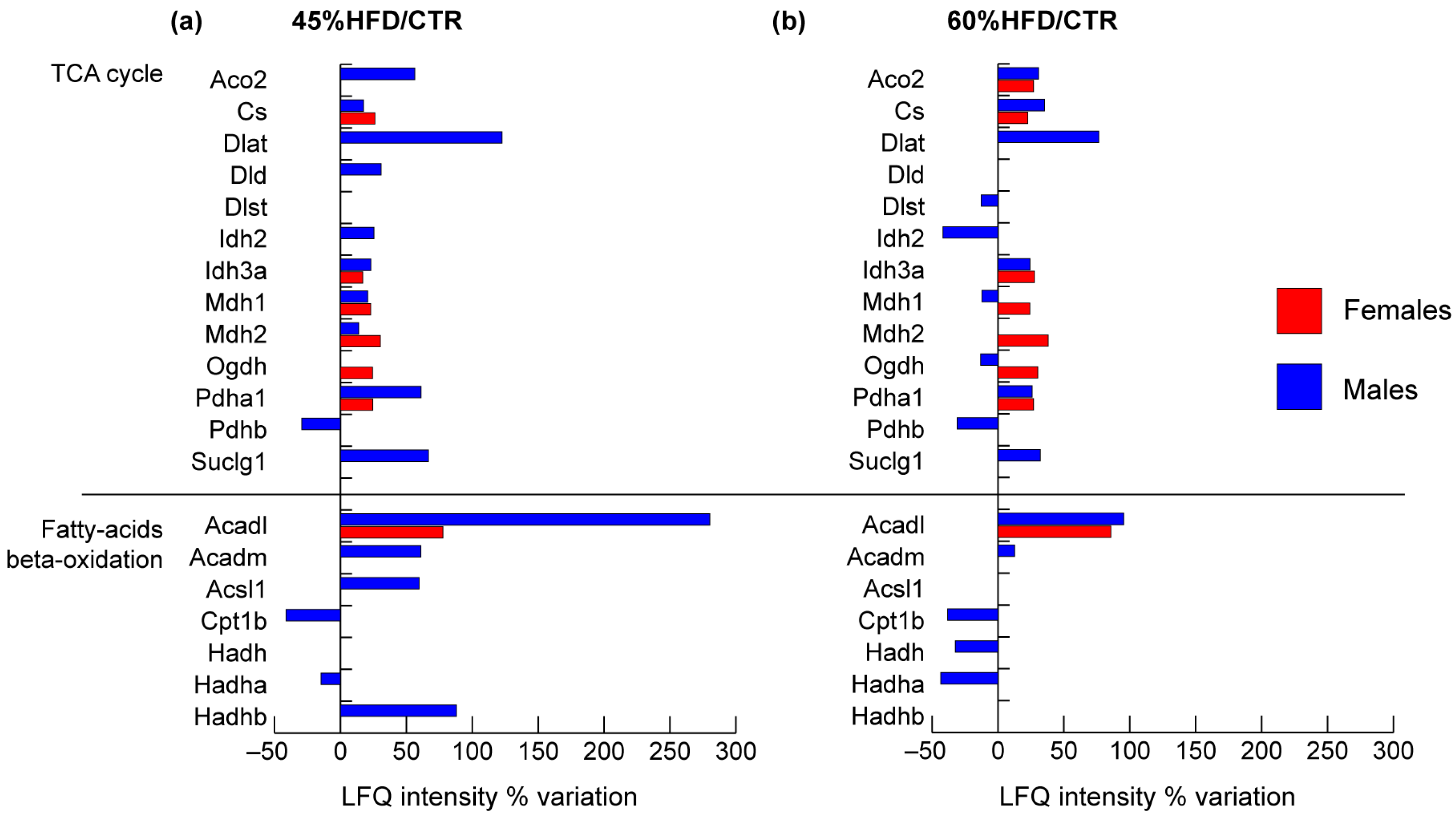
2.2.4. TCA Cycle and Fatty Acids Metabolism
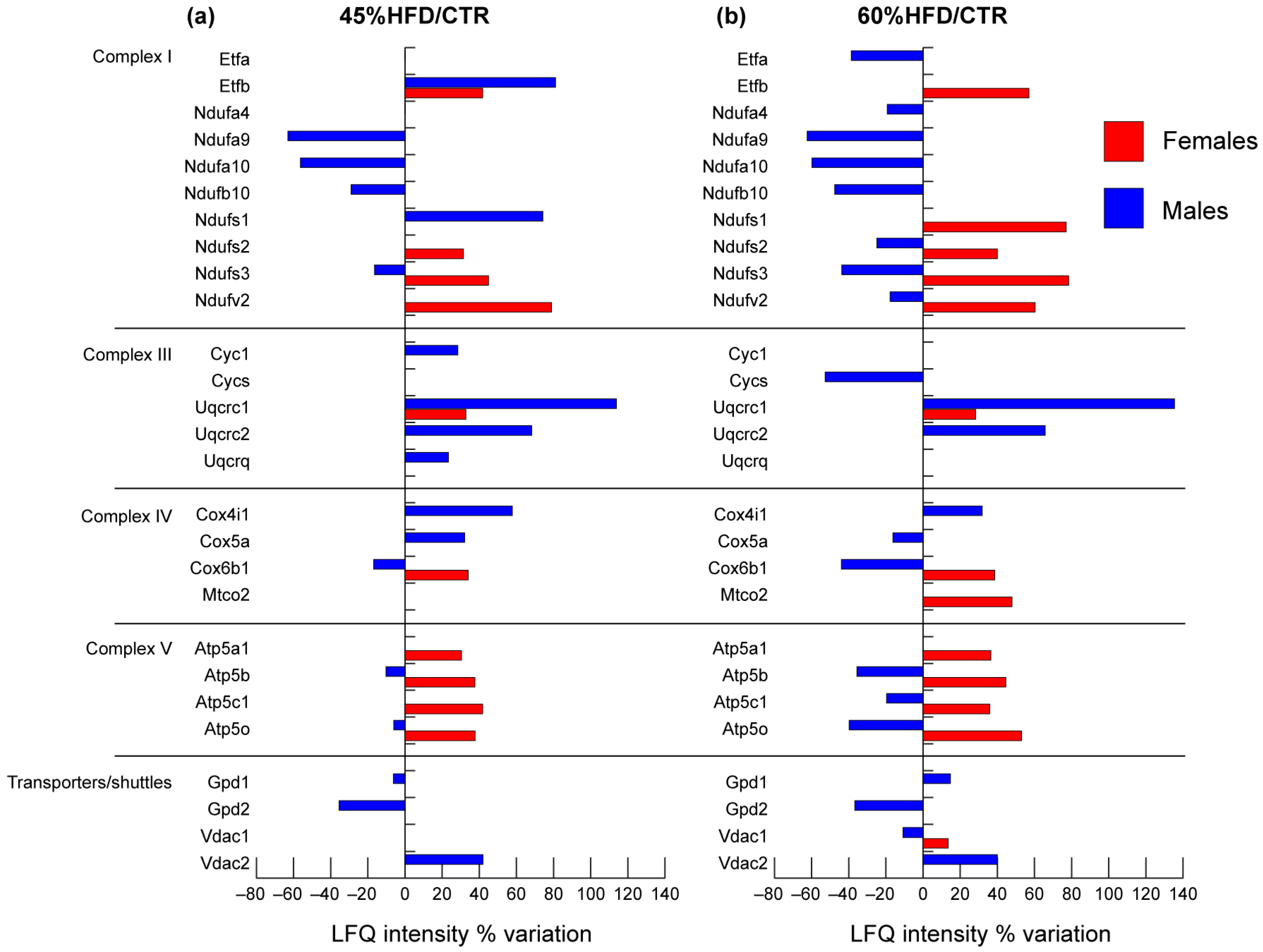
2.2.5. Mitochondrial Respiratory Chain
2.3. Bioinformatics Analysis of Proteomics Results
2.3.1. Canonical Pathways
2.3.2. Diseases and Biofunctions
2.3.3. Upstream Regulators
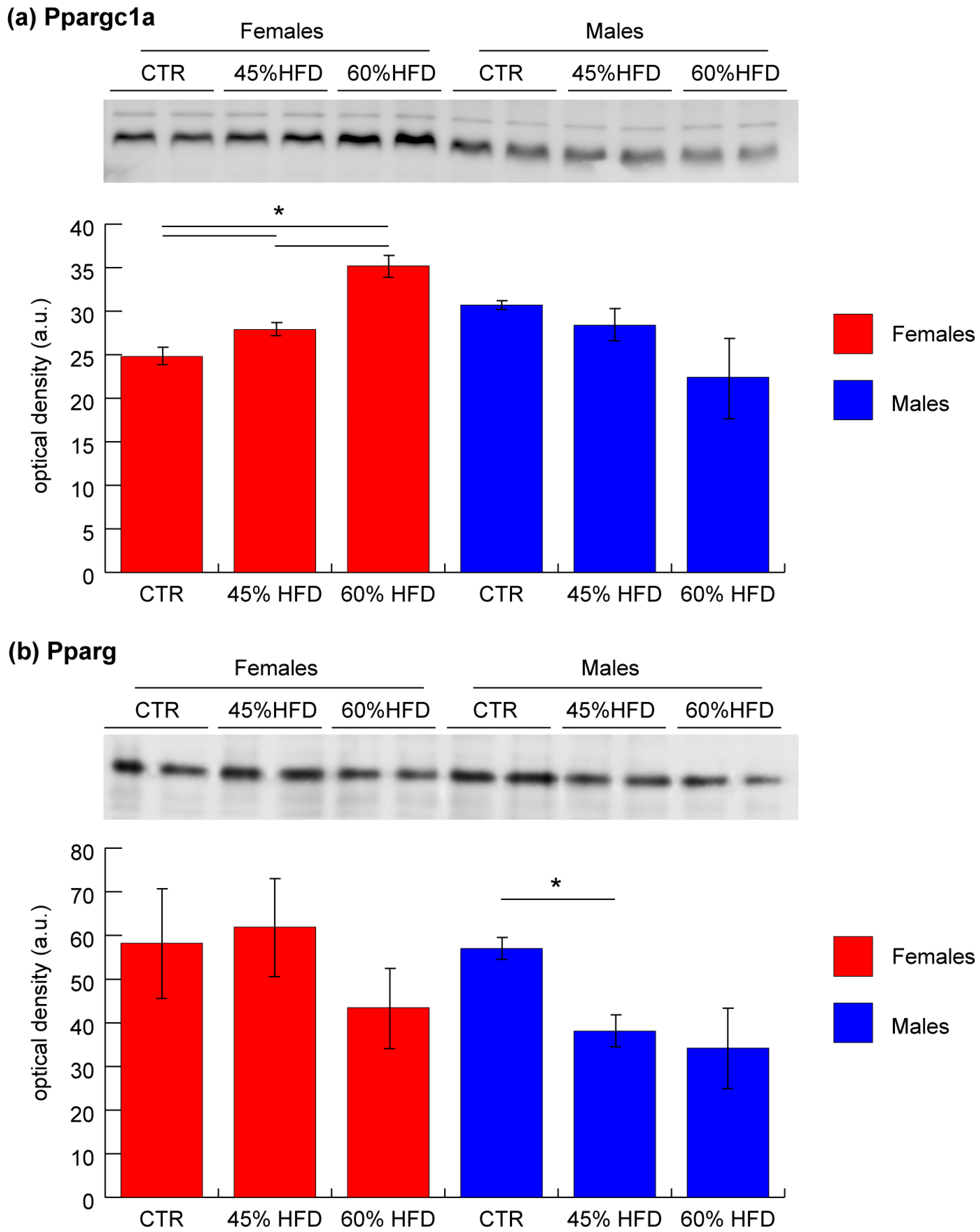
2.4. Validation of Ppargc1a and Pparg by Immunoblotting
3. Discussion
4. Materials and Methods
4.1. Animals, Diet Administration and Glucose Tolerance Test
4.2. MRI Study
4.3. Statistical Analysis
4.4. Muscle Biopsies, Protein Extraction and Quantification
4.5. Label-Free Liquid Chromatography with Tandem Mass Spectrometry (LC–MS/MS)
4.6. Ingenuity Pathway Analysis
4.7. Immunoblotting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECD. Obesity Update 2017. Available online: http://www.oecd.org/health/obesity-update.htm (accessed on 15 December 2020).
- Bluher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- Goossens, G.H. The role of adipose tissue dysfunction in the pathogenesis of obesity-related insulin resistance. Physiol. Behav. 2008, 94, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Fruh, S.M. Obesity: Risk factors, complications, and strategies for sustainable long-term weight management. J. Am. Assoc. Nurse Pract. 2017, 29, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Abate, M. How obesity modifies tendons (implications for athletic activities). Muscles Ligaments Tendons J. 2014, 4, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Gaida, J.E.; Alfredson, H.; Kiss, Z.S.; Bass, S.L.; Cook, J.L. Asymptomatic Achilles tendon pathology is associated with a central fat distribution in men and a peripheral fat distribution in women: A cross sectional study of 298 individuals. BMC Musculoskelet. Disord. 2010, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.L.; Messer, L.C.; Rappazzo, K.M.; Jagai, J.S.; Grabich, S.C.; Lobdell, D.T. The association between physical inactivity and obesity is modified by five domains of environmental quality in U.S. adults: A cross-sectional study. PLoS ONE 2018, 13, e0203301. [Google Scholar] [CrossRef]
- Moellering, D.R.; Smith, D.L., Jr. Ambient temperature and obesity. Curr. Obes. Rep. 2012, 1, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soubry, A.; Guo, L.; Huang, Z.; Hoyo, C.; Romanus, S.; Price, T.; Murphy, S.K. Obesity-Related DNA methylation at imprinted genes in human sperm: Results from the TIEGER study. Clin. Epigenet. 2016, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Link, J.C.; Reue, K. Genetic basis for sex differences in obesity and lipid metabolism. Annu. Rev. Nutr. 2017, 37, 225–245. [Google Scholar] [CrossRef]
- Villareal, D.T.; Apovian, C.M.; Kushner, R.F.; Klein, S. Obesity in older adults: Technical review and position statement of the American Society for Nutrition and NAASO, The Obesity Society. Obes. Res. 2005, 13, 1849–1863. [Google Scholar] [CrossRef]
- Turnbaugh, P.J. Microbes and diet-induced obesity: Fast, cheap, and out of control. Cell Host Microbe 2017, 21, 278–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popkin, B.M. The nutrition transition and obesity in the developing world. J. Nutr. 2001, 131, 871S–873S. [Google Scholar] [CrossRef] [PubMed]
- Drewnowski, A. Nutrition transition and global dietary trends. Nutrition 2000, 16, 486–487. [Google Scholar] [CrossRef]
- Freeman, A.M.; Pennings, N. Insulin Resistance; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Daghri, N.M.; Torretta, E.; Barbacini, P.; Asare, H.; Ricci, C.; Capitanio, D.; Guerini, F.R.; Sabico, S.B.; Alokail, M.S.; Clerici, M.; et al. Sphingolipid serum profiling in vitamin D deficient and dyslipidemic obese dimorphic adults. Sci. Rep. 2019, 9, 16664. [Google Scholar] [CrossRef] [PubMed]
- Torretta, E.; Barbacini, P.; Al-Daghri, N.M.; Gelfi, C. Sphingolipids in obesity and correlated co-morbidities: The contribution of gender, age and environment. Int. J. Mol. Sci. 2019, 20, 5901. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ballantyne, C.M. Skeletal muscle inflammation and insulin resistance in obesity. J. Clin. Investig. 2017, 127, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.N.; Choi, K.M. The implications of sarcopenia and sarcopenic obesity on cardiometabolic disease. J. Cell. Biochem. 2015, 116, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Xu, Y. Association of sarcopenic obesity with the risk of all-cause mortality: A meta-analysis of prospective cohort studies. Geriatr. Gerontol. Int. 2016, 16, 155–166. [Google Scholar] [CrossRef]
- Herrenbruck, A.R.; Bollinger, L.M. Role of skeletal muscle autophagy in high-fat-diet-induced obesity and exercise. Nutr. Rev. 2020, 78, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Tallis, J.; James, R.S.; Seebacher, F. The effects of obesity on skeletal muscle contractile function. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinkovich, A.; Livshits, G. Sarcopenic obesity or obese sarcopenia: A cross talk between age-associated adipose tissue and skeletal muscle inflammation as a main mechanism of the pathogenesis. Ageing Res. Rev. 2017, 35, 200–221. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Freire, M.; Semba, R.D.; Ubaida-Mohien, C.; Fabbri, E.; Scalzo, P.; Hojlund, K.; Dufresne, C.; Lyashkov, A.; Ferrucci, L. The human skeletal muscle proteome project: A reappraisal of the current literature. J. Cachexia Sarcopenia Muscle 2017, 8, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Capitanio, D.; Moriggi, M.; Gelfi, C. Mapping the human skeletal muscle proteome: Progress and potential. Expert Rev. Proteom. 2017, 14, 825–839. [Google Scholar] [CrossRef]
- Ubaida-Mohien, C.; Lyashkov, A.; Gonzalez-Freire, M.; Tharakan, R.; Shardell, M.; Moaddel, R.; Semba, R.D.; Chia, C.W.; Gorospe, M.; Sen, R.; et al. Discovery proteomics in aging human skeletal muscle finds change in spliceosome, immunity, proteostasis and mitochondria. eLife 2019, 8. [Google Scholar] [CrossRef]
- Deshmukh, A.S.; Murgia, M.; Nagaraj, N.; Treebak, J.T.; Cox, J.; Mann, M. Deep proteomics of mouse skeletal muscle enables quantitation of protein isoforms, metabolic pathways, and transcription factors. Mol. Cell. Proteom. MCP 2015, 14, 841–853. [Google Scholar] [CrossRef] [Green Version]
- La, G.; Zhou, M.; Lim, J.Y.; Oh, S.; Xing, H.; Liu, N.; Yang, Y.; Liu, X.; Zhong, L. Proteomics and transcriptomics analysis reveals clues into the mechanism of the beneficial effect of electrical stimulation on rat denervated gastrocnemius muscle. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2019, 52, 769–786. [Google Scholar] [CrossRef]
- Pedditzi, E.; Peters, R.; Beckett, N. The risk of overweight/obesity in mid-life and late life for the development of dementia: A systematic review and meta-analysis of longitudinal studies. Age Ageing 2016, 45, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Sharabi, K.; Tavares, C.D.; Rines, A.K.; Puigserver, P. Molecular pathophysiology of hepatic glucose production. Mol. Asp. Med. 2015, 46, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef] [Green Version]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef]
- Geer, E.B.; Shen, W. Gender differences in insulin resistance, body composition, and energy balance. Gend. Med. 2009, 6 (Suppl. 1), 60–75. [Google Scholar] [CrossRef] [Green Version]
- Palmer, B.F.; Clegg, D.J. The sexual dimorphism of obesity. Mol. Cell. Endocrinol. 2015, 402, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, P. Differences in lipolysis between human subcutaneous and omental adipose tissues. Ann. Med. 1995, 27, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Varghese, M.; Singer, K. Gender and sex differences in adipose tissue. Curr. Diabetes Rep. 2018, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, P.S.; Thornhill, A.R.; Boudrean, S.K.; Darling, S.M.; Bishop, C.E.; Evans, E.P. The genetic basis of XX-XY differences present before gonadal sex differentiation in the mouse. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1995, 350, 253–260. [Google Scholar] [CrossRef]
- Wijchers, P.J.; Yandim, C.; Panousopoulou, E.; Ahmad, M.; Harker, N.; Saveliev, A.; Burgoyne, P.S.; Festenstein, R. Sexual dimorphism in mammalian autosomal gene regulation is determined not only by Sry but by sex chromosome complement as well. Dev. Cell 2010, 19, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewing, P.; Chiang, C.W.; Sinchak, K.; Sim, H.; Fernagut, P.O.; Kelly, S.; Chesselet, M.F.; Micevych, P.E.; Albrecht, K.H.; Harley, V.R.; et al. Direct regulation of adult brain function by the male-specific factor SRY. Curr. Biol. CB 2006, 16, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokop, J.W.; Deschepper, C.F. Chromosome Y genetic variants: Impact in animal models and on human disease. Physiol. Genom. 2015, 47, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Ongusaha, P.P.; Miles, P.D.; Havstad, J.C.; Zhang, F.; So, W.V.; Kudlow, J.E.; Michell, R.H.; Olefsky, J.M.; Field, S.J.; et al. Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance. Nature 2008, 451, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.P.; McGreal, S.R.; Graw, S.; Tessman, R.; Koppel, S.J.; Dhakal, P.; Zhang, Z.; Machacek, M.; Zachara, N.E.; Koestler, D.C.; et al. Sustained O-GlcNAcylation reprograms mitochondrial function to regulate energy metabolism. J. Biol. Chem. 2017, 292, 14940–14962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.P.; Bale, T.L. Sex differences in microRNA regulation of gene expression: No smoke, just miRs. Biol. Sex Differ. 2012, 3, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berletch, J.B.; Yang, F.; Disteche, C.M. Escape from X inactivation in mice and humans. Genome Biol. 2010, 11, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victoria, B.; Nunez Lopez, Y.O.; Masternak, M.M. MicroRNAs and the metabolic hallmarks of aging. Mol. Cell. Endocrinol. 2017, 455, 131–147. [Google Scholar] [CrossRef]
- Xie, H.; Lim, B.; Lodish, H.F. MicroRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009, 58, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Lundsgaard, A.M.; Kiens, B. Gender differences in skeletal muscle substrate metabolism—Molecular mechanisms and insulin sensitivity. Front. Endocrinol. 2014, 5, 195. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Huenchullan, S.; McLennan, S.V.; Verhoeven, A.; Twigg, S.M.; Tam, C.S. The emerging role of skeletal muscle extracellular matrix remodelling in obesity and exercise. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2017, 18, 776–790. [Google Scholar] [CrossRef]
- Collins, K.H.; Herzog, W.; MacDonald, G.Z.; Reimer, R.A.; Rios, J.L.; Smith, I.C.; Zernicke, R.F.; Hart, D.A. Obesity, metabolic syndrome, and musculoskeletal disease: Common inflammatory pathways suggest a central role for loss of muscle integrity. Front. Physiol. 2018, 9, 112. [Google Scholar] [CrossRef]
- Duan, Y.; Li, F.; Tan, B.; Yao, K.; Yin, Y. Metabolic control of myofibers: Promising therapeutic target for obesity and type 2 diabetes. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2017, 18, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Stuart, C.A.; McCurry, M.P.; Marino, A.; South, M.A.; Howell, M.E.; Layne, A.S.; Ramsey, M.W.; Stone, M.H. Slow-Twitch fiber proportion in skeletal muscle correlates with insulin responsiveness. J. Clin. Endocrinol. Metab. 2013, 98, 2027–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, C.A.; South, M.A.; Lee, M.L.; McCurry, M.P.; Howell, M.E.; Ramsey, M.W.; Stone, M.H. Insulin responsiveness in metabolic syndrome after eight weeks of cycle training. Med. Sci. Sports Exerc. 2013, 45, 2021–2029. [Google Scholar] [CrossRef] [Green Version]
- Gaster, M.; Staehr, P.; Beck-Nielsen, H.; Schroder, H.D.; Handberg, A. GLUT4 is reduced in slow muscle fibers of type 2 diabetic patients: Is insulin resistance in type 2 diabetes a slow, type 1 fiber disease? Diabetes 2001, 50, 1324–1329. [Google Scholar] [CrossRef] [Green Version]
- Tanner, C.J.; Barakat, H.A.; Dohm, G.L.; Pories, W.J.; MacDonald, K.G.; Cunningham, P.R.; Swanson, M.S.; Houmard, J.A. Muscle fiber type is associated with obesity and weight loss. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E1191–E1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshima, H.; Tamura, Y.; Kakehi, S.; Kurebayashi, N.; Murayama, T.; Nakamura, K.; Kakigi, R.; Okada, T.; Sakurai, T.; Kawamori, R.; et al. Long-Term, but not short-term high-fat diet induces fiber composition changes and impaired contractile force in mouse fast-twitch skeletal muscle. Physiol. Rep. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, J.; Newby, F.D.; Gebhart, S.S.; DiGirolamo, M. Insulin resistance in obesity is associated with elevated basal lactate levels and diminished lactate appearance following intravenous glucose and insulin. Metab. Clin. Exp. 1992, 41, 22–27. [Google Scholar] [CrossRef]
- Friedman, J.E.; Caro, J.F.; Pories, W.J.; Azevedo, J.L., Jr.; Dohm, G.L. Glucose metabolism in incubated human muscle: Effect of obesity and non-insulin-dependent diabetes mellitus. Metab. Clin. Exp. 1994, 43, 1047–1054. [Google Scholar] [CrossRef]
- Hulver, M.W.; Berggren, J.R.; Cortright, R.N.; Dudek, R.W.; Thompson, R.P.; Pories, W.J.; MacDonald, K.G.; Cline, G.W.; Shulman, G.I.; Dohm, G.L.; et al. Skeletal muscle lipid metabolism with obesity. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E741–E747. [Google Scholar] [CrossRef]
- Kim, J.Y.; Hickner, R.C.; Cortright, R.L.; Dohm, G.L.; Houmard, J.A. Lipid oxidation is reduced in obese human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E1039–E1044. [Google Scholar] [CrossRef] [Green Version]
- Heilbronn, L.K.; Gregersen, S.; Shirkhedkar, D.; Hu, D.; Campbell, L.V. Impaired fat oxidation after a single high-fat meal in insulin-sensitive nondiabetic individuals with a family history of type 2 diabetes. Diabetes 2007, 56, 2046–2053. [Google Scholar] [CrossRef] [Green Version]
- Sparks, L.M.; Xie, H.; Koza, R.A.; Mynatt, R.; Hulver, M.W.; Bray, G.A.; Smith, S.R. A high-fat diet coordinately downregulates genes required for mitochondrial oxidative phosphorylation in skeletal muscle. Diabetes 2005, 54, 1926–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleier, L.; Drose, S. Superoxide generation by complex III: From mechanistic rationales to functional consequences. Biochim. Biophys. Acta 2013, 1827, 1320–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Sanchez, A.; Madrigal-Santillan, E.; Bautista, M.; Esquivel-Soto, J.; Morales-Gonzalez, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sanchez-Rivera, G.; Valadez-Vega, C.; Morales-Gonzalez, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [Green Version]
- Catalan, V.F.; Frühbeck, G.; Gomez-Ambrosi, J. Inflammatory and oxidative stress markers in skeletal muscle of obese subjects. In Obesity; Amelia Marti del Moral, C.M.A.G., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 163–189. [Google Scholar] [CrossRef]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated regulation of skeletal muscle hypertrophy and atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
- Hevener, A.L.; Zhou, Z.; Moore, T.M.; Drew, B.G.; Ribas, V. The impact of ERalpha action on muscle metabolism and insulin sensitivity—Strong enough for a man, made for a woman. Mol. Metab. 2018, 15, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Knutti, D.; Kralli, A. PGC-1, a versatile coactivator. Trends Endocrinol. Metab. TEM 2001, 12, 360–365. [Google Scholar] [CrossRef]
- Russell, A.P. PGC-1alpha and exercise: Important partners in combating insulin resistance. Curr. Diabetes Rev. 2005, 1, 175–181. [Google Scholar] [CrossRef]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef]
- Michael, L.F.; Wu, Z.; Cheatham, R.B.; Puigserver, P.; Adelmant, G.; Lehman, J.J.; Kelly, D.P.; Spiegelman, B.M. Restoration of insulin-sensitive glucose transporter (GLUT4) gene expression in muscle cells by the transcriptional coactivator PGC-1. Proc. Natl. Acad. Sci. USA 2001, 98, 3820–3825. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, S.N.; Emter, R.; Hock, M.B.; Knutti, D.; Cardenas, J.; Podvinec, M.; Oakeley, E.J.; Kralli, A. The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 6472–6477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstrale, M.; Laurila, E.; et al. PGC-1alpha-Responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, S.; Pharaoh, G.; Ranjit, R.; Murphy, A.; Matsuzaki, S.; Nair, B.C.; Forbes, B.; Gispert, S.; Auburger, G.; Humphries, K.M.; et al. Loss of mitochondrial protease ClpP protects mice from diet-induced obesity and insulin resistance. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Kukat, A.; Szczepanowska, K.; Hermans, S.; Senft, K.; Brandscheid, C.P.; Maiti, P.; Trifunovic, A. CLPP deficiency protects against metabolic syndrome but hinders adaptive thermogenesis. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; He, L.; Johnson, M.S.; Li, Y.; Zeng, L.; Ding, Y.; Long, Q.; Moore, J.F.; Sharer, J.D.; Nagy, T.R.; et al. Carnitine palmitoyltransferase 1b deficiency protects mice from diet-induced insulin resistance. J. Diabetes Metab. 2014, 5, 361. [Google Scholar] [CrossRef] [Green Version]
- Puzio-Kuter, A.M. The Role of p53 in Metabolic regulation. Genes Cancer 2011, 2, 385–391. [Google Scholar] [CrossRef]
- Qi, Z.; He, J.; Zhang, Y.; Shao, Y.; Ding, S. Exercise training attenuates oxidative stress and decreases p53 protein content in skeletal muscle of type 2 diabetic Goto-Kakizaki rats. Free Radic. Biol. Med. 2011, 50, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Kung, C.P.; Murphy, M.E. The role of the p53 tumor suppressor in metabolism and diabetes. J. Endocrinol. 2016, 231, R61–R75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varaljai, R.; Islam, A.B.; Beshiri, M.L.; Rehman, J.; Lopez-Bigas, N.; Benevolenskaya, E.V. Increased mitochondrial function downstream from KDM5A histone demethylase rescues differentiation in pRB-deficient cells. Genes Dev. 2015, 29, 1817–1834. [Google Scholar] [CrossRef] [Green Version]
- Hevener, A.L.; He, W.; Barak, Y.; Le, J.; Bandyopadhyay, G.; Olson, P.; Wilkes, J.; Evans, R.M.; Olefsky, J. Muscle-Specific Pparg deletion causes insulin resistance. Nat. Med. 2003, 9, 1491–1497. [Google Scholar] [CrossRef]
- Fu, Y.; Luo, L.; Luo, N.; Zhu, X.; Garvey, W.T. NR4A orphan nuclear receptors modulate insulin action and the glucose transport system: Potential role in insulin resistance. J. Biol. Chem. 2007, 282, 31525–31533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khamzina, L.; Veilleux, A.; Bergeron, S.; Marette, A. Increased activation of the mammalian target of rapamycin pathway in liver and skeletal muscle of obese rats: Possible involvement in obesity-linked insulin resistance. Endocrinology 2005, 146, 1473–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, J.J.; Ruthenborg, R.J.; Cho, K.W.; Kim, J.W. Regulation of obesity and insulin resistance by hypoxia-inducible factors. Hypoxia 2014, 2, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, E.M.; Meers, G.M.E.; Ruegsegger, G.N.; Wankhade, U.D.; Robinson, T.; Koch, L.G.; Britton, S.L.; Rector, R.S.; Shankar, K.; Thyfault, J.P. Intrinsic high aerobic capacity in male rats protects against diet-induced insulin resistance. Endocrinology 2019, 160, 1179–1192. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.F.; Duarte, J.M.N.; Gruetter, R. Increased hepatic fatty acid polyunsaturation precedes ectopic lipid deposition in the liver in adaptation to high-fat diets in mice. MAGMA 2018, 31, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canonical Pathways 45% HFD/CTR | Females | Males | Canonical Pathways 60% HFD/CTR | Females | Males |
|---|---|---|---|---|---|
| Oxidative phosphorylation | 3 | 0 | Oxidative phosphorylation | 3.162 | −2.5 |
| TCA cycle II (eukaryotic) | 2.236 | 2.646 | TCA cycle II (eukaryotic) | 2.449 | 0.378 |
| Actin cytoskeleton-signaling | N/A | 3.207 | Glycolysis I | N/A | 2.121 |
| Estrogen-receptor-signaling | N/A | 2.449 | |||
| Regulation of actin-based motility by Rho | N/A | 2.236 | |||
| Gluconeogenesis I | N/A | 2.236 | |||
| ILK-signaling | N/A | 2.138 | |||
| Protein kinase A-signaling | N/A | −2.236 | |||
| RhoGDI-signaling | N/A | −2.449 |
| Diseases and Bio Functions 45% HFD/CTR | Females | Males | Diseases and Bio Functions 60% HFD/CTR | Females | Males |
|---|---|---|---|---|---|
| Cell death of muscle cells | −2 | 0.302 | Concentration of lipid | N/A | 2.779 |
| Necrosis of muscle | −2.236 | N/A | Concentration of fatty acid | N/A | 1.99 |
| Necrosis | −2.927 | 0.154 | Transmembrane potential of mitochondria | N/A | −1.964 |
| Damage of muscle | N/A | 1.982 | Consumption of oxygen | N/A | −1.982 |
| Quantity of glycogen | N/A | 1.977 | |||
| Quantity of carbohydrate | N/A | 1.821 | |||
| Concentration of fatty acid | N/A | 1.65 | |||
| Cellular homeostasis | N/A | −1.644 | |||
| Transmembrane potential | N/A | −2.2 | |||
| Transmembrane potential of mitochondria | N/A | −2.2 |
| Upstream Regulators 45% HFD/CTR | Females | Males | Upstream Regulators 60% HFD/CTR | Females | Males |
|---|---|---|---|---|---|
| Insr | 3.561 | 2.02 | Insr | 3.38 | 0.191 |
| Ppargc1a | 3.124 | 0.904 | Ppargc1a | 3.279 | −0.299 |
| Rb1 | 2.449 | 0.335 | Rb1 | 2.333 | −0.709 |
| Igf1r | 2.236 | 0.632 | Igf1r | 2.236 | −0.905 |
| Esrra | 2.219 | 0.154 | Pten | 2.182 | 0.688 |
| Ppargc1b | 2 | 1.026 | Stk11 | 2.121 | 0.471 |
| Myc | 0.831 | 2.213 | Ppargc1b | 2 | −0.396 |
| Nr4a1 | −1 | −3.174 | Map4k4 | −2.236 | 0.535 |
| Tp53 | −2.111 | −1.035 | Kdm5a | −2.333 | 0.816 |
| Kdm5a | −2.449 | −0.626 | Tp53 | −2.887 | −0.671 |
| Rictor | −3 | −0.258 | Rictor | −3.162 | 2 |
| Clpp | −3.051 | −2.449 | Clpp | −3.207 | 0.209 |
| Cpt1b | −3.138 | −0.249 | Cpt1b | −3.293 | 2.467 |
| Pparg | N/A | −2.18 | Hif1a | N/A | 2.211 |
| Rptor | N/A | 2.425 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriggi, M.; Belloli, S.; Barbacini, P.; Murtaj, V.; Torretta, E.; Chaabane, L.; Canu, T.; Penati, S.; Malosio, M.L.; Esposito, A.; et al. Skeletal Muscle Proteomic Profile Revealed Gender-Related Metabolic Responses in a Diet-Induced Obesity Animal Model. Int. J. Mol. Sci. 2021, 22, 4680. https://doi.org/10.3390/ijms22094680
Moriggi M, Belloli S, Barbacini P, Murtaj V, Torretta E, Chaabane L, Canu T, Penati S, Malosio ML, Esposito A, et al. Skeletal Muscle Proteomic Profile Revealed Gender-Related Metabolic Responses in a Diet-Induced Obesity Animal Model. International Journal of Molecular Sciences. 2021; 22(9):4680. https://doi.org/10.3390/ijms22094680
Chicago/Turabian StyleMoriggi, Manuela, Sara Belloli, Pietro Barbacini, Valentina Murtaj, Enrica Torretta, Linda Chaabane, Tamara Canu, Silvia Penati, Maria Luisa Malosio, Antonio Esposito, and et al. 2021. "Skeletal Muscle Proteomic Profile Revealed Gender-Related Metabolic Responses in a Diet-Induced Obesity Animal Model" International Journal of Molecular Sciences 22, no. 9: 4680. https://doi.org/10.3390/ijms22094680
APA StyleMoriggi, M., Belloli, S., Barbacini, P., Murtaj, V., Torretta, E., Chaabane, L., Canu, T., Penati, S., Malosio, M. L., Esposito, A., Gelfi, C., Moresco, R. M., & Capitanio, D. (2021). Skeletal Muscle Proteomic Profile Revealed Gender-Related Metabolic Responses in a Diet-Induced Obesity Animal Model. International Journal of Molecular Sciences, 22(9), 4680. https://doi.org/10.3390/ijms22094680