
Deep Transcranial Magnetic Stimulation Affects Gut Microbiota Composition in Obesity: Results of Randomized Clinical Trial

 ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Participants Characteristics
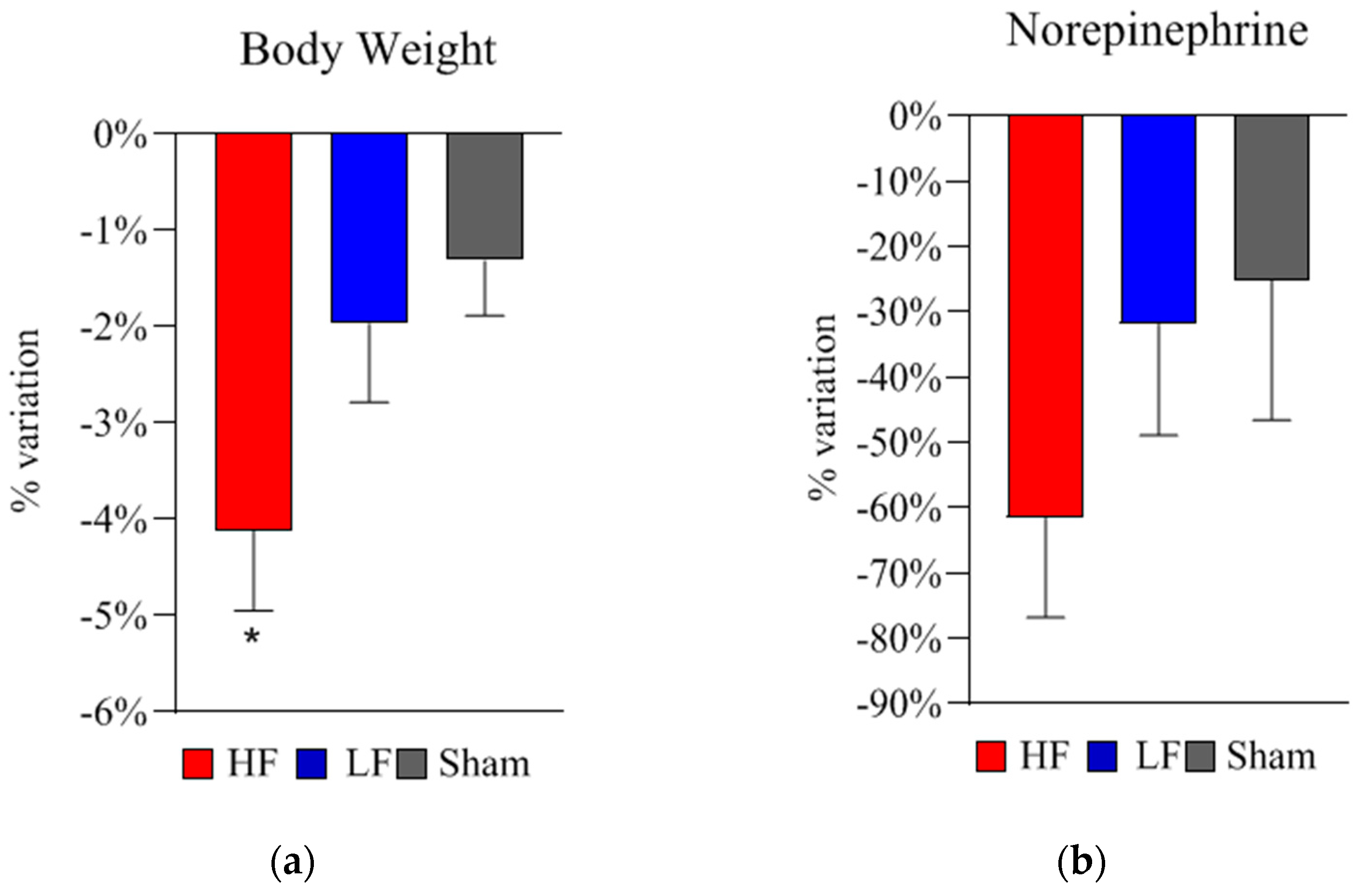
2.2. Bodyweight, Metabolic, and Neurohormonal Parameters
2.3. Bacterial Changes at Phylum Level in Gut Microbiota Composition after 5-Week dTMS Treatment
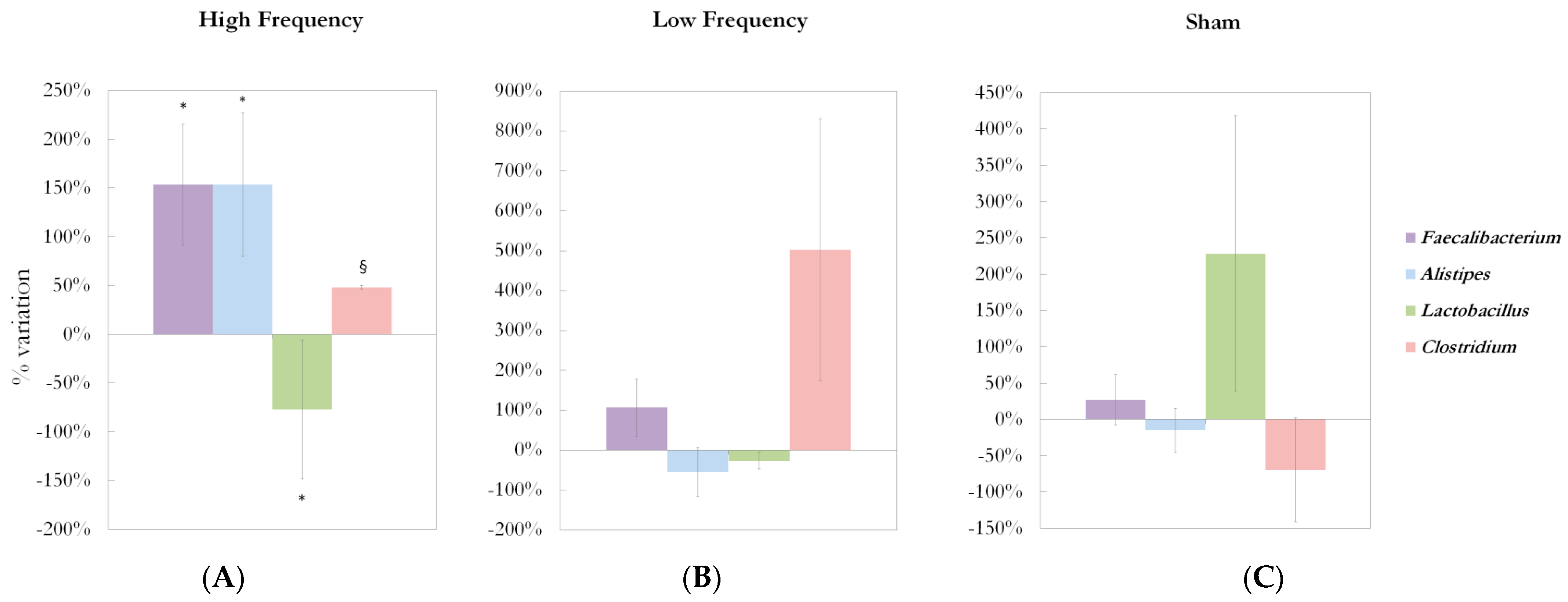
2.4. Bacterial Changes at Genus Level in Gut Microbiota Composition after 5-Week dTMS Treatment
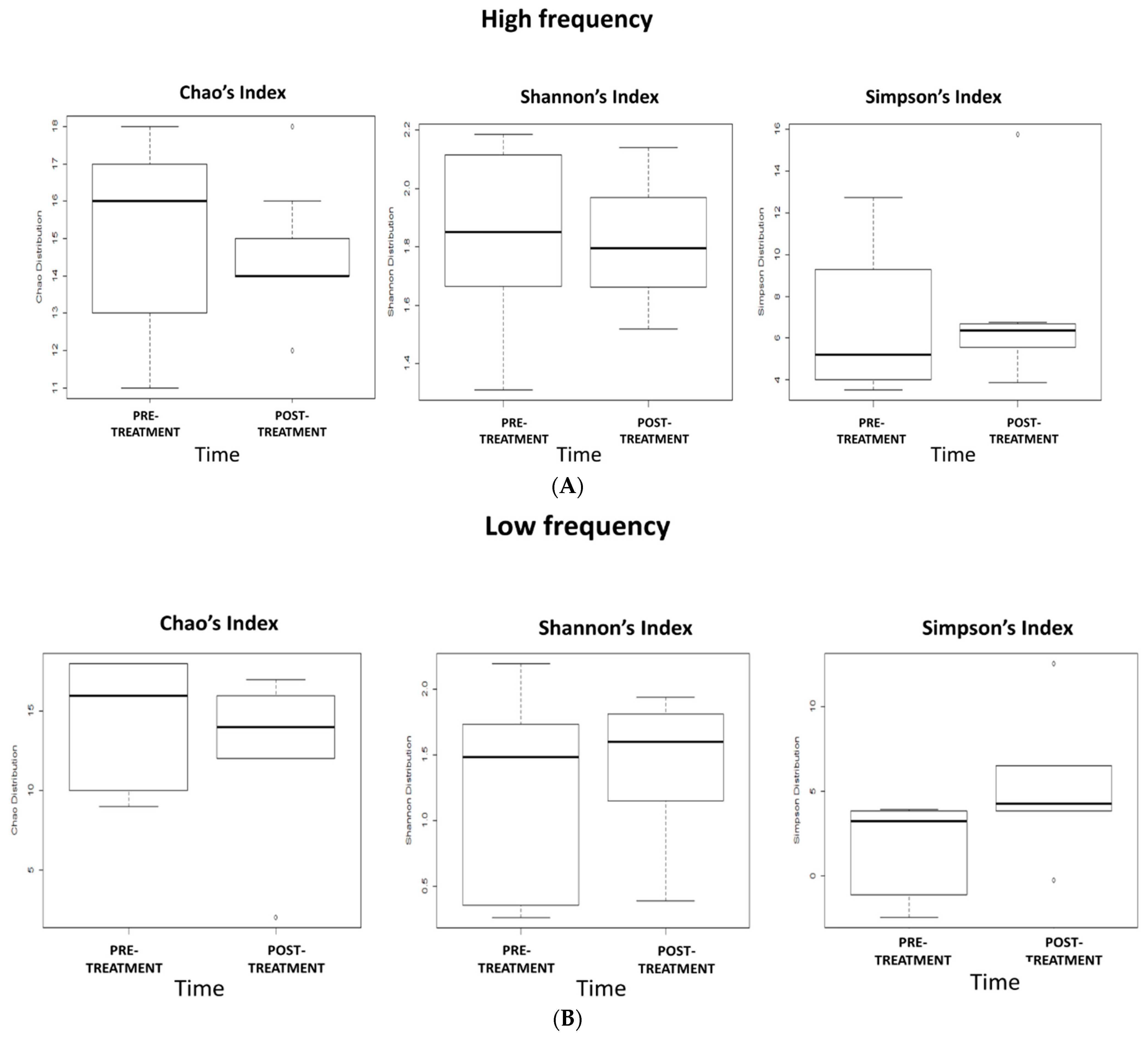
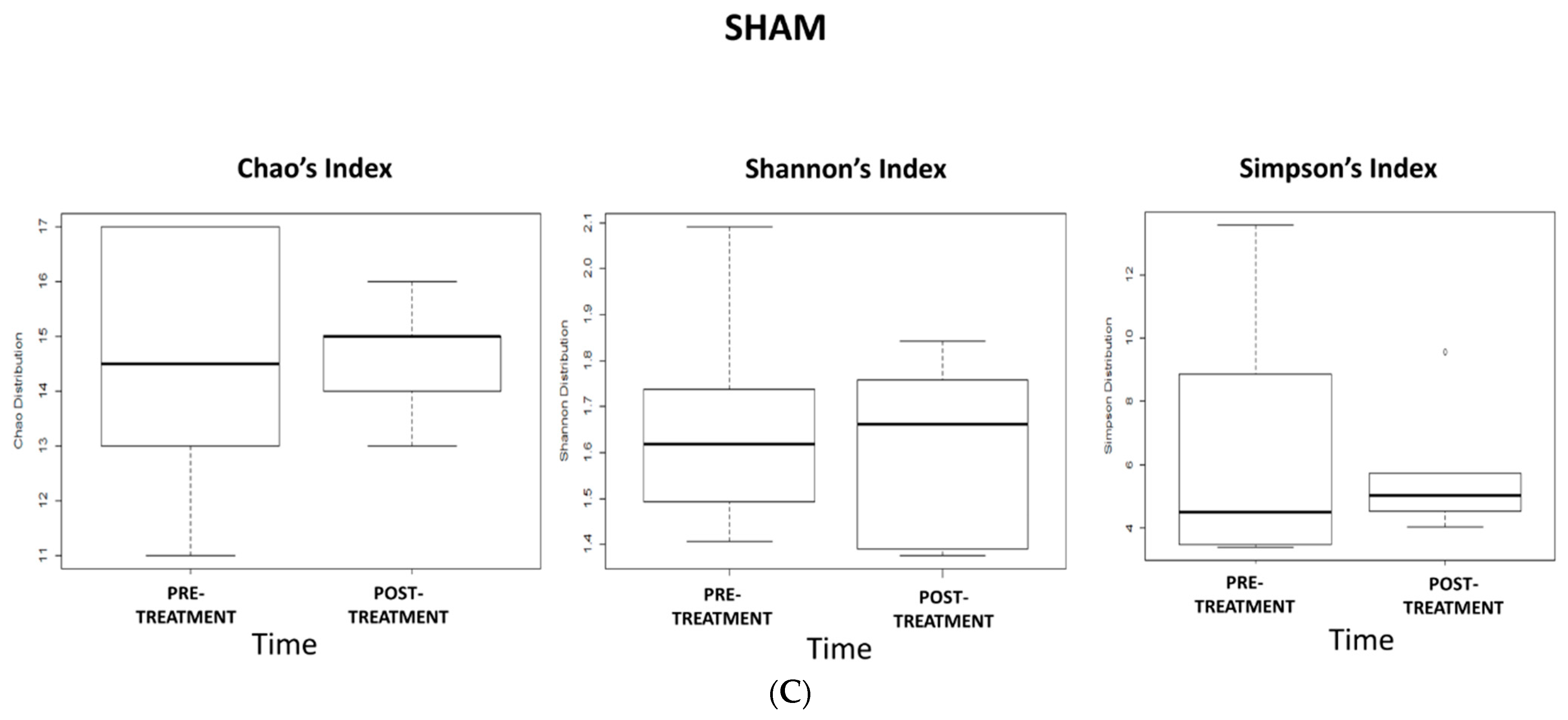
2.5. Effects of 5-Week dTMS Treatment on Gut Microbiota Biodiversity
2.6. Correlations Between Variations in Metabolic/Neurohormonal Parameters and Gut Microbiota Composition
2.7. Adverse Events and Safety
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Study Design
4.3. Repetitive Deep Transcranial Stimulation Procedure (dTMS)
4.4. Diet and Lifestyle Recommendations
4.5. Measurements
4.5.1. Anthropometric Values
4.5.2. Body Composition
4.5.3. Resting Energy Expenditure (REE) and Respiratory Quotient (RQ)
4.5.4. Laboratory Measurements
4.5.5. Gut Microbiota Analysis
DNA Extraction
16 S rRNA Gene Amplification
Ion Torrent PGM Sequencing of 16S rRNA Gene-Based Amplicons
4.5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Clercq, N.; Frissen, M.N.; Groen, A.K.; Nieuwdorp, M. Gut microbiota and the gut-brain axis: New insights in the pathophysiology of metabolic syndrome. Psychosom. Med. 2017, 79, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and metabolic diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Guirro, M.; Costa, A.; Gual-Grau, A.; Herrero, P.; Torrell, H.; Canela, N.; Arola, L. Effects from diet-induced gut microbiota dysbiosis and obesity can be ameliorated by fecal microbiota transplantation: A multiomics approach. PLoS ONE 2019, 14, e0218143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loke, Y.L.; Chew, M.T.; Ngeow, Y.F.; Lim, W.W.D.; Peh, S.C. Colon Carcinogenesis: The Interplay Between Diet and Gut Microbiota. Front. Cell Infect. Microbiol. 2020, 10, 603086. [Google Scholar] [CrossRef]
- Rogers, C.J.; Prabhu, K.S.; Vijay-Kumar, M. The microbiome and obesity-an established risk for certain types of cancer. Cancer J. 2014, 20, 176–180. [Google Scholar] [CrossRef]
- Cândido, F.G.; Valente, F.X.; Grześkowiak, Ł.M.; Moreira, A.P.B.; Rocha, D.M.U.P.; Alfenas, R.C.G. Impact of dietary fat on gut microbiota and low-grade systemic inflammation: Mechanisms and clinical implications on obesity. Int. J. Food Sci. Nutr. 2017, 4, 1–19. [Google Scholar] [CrossRef]
- Blaut, M. Gut microbiota and energy balance: Role in obesity. Proc. Nutr. Soc. 2015, 74, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Osadchiy, V.; Mayer, E.A. Brain-gut-microbiome interactions in obesity and food addiction. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 655–672. [Google Scholar] [CrossRef]
- Blum, K.; Thanos, P.K.; Gold, M.S. Dopamine and glucose, obesity, and reward deficiency syndrome. Front. Psychol. 2014, 5, 919. [Google Scholar] [CrossRef] [Green Version]
- Foster, J.A.; Lyte, M.; Meyer, E.; Cryan, J.F. Gut Microbiota and Brain Function: An Evolving Field in Neuroscience. Int. J. Neuropsychopharmacol. 2016, 19, pyv114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; Koga, Y.; Sudo, N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1288–G1295. [Google Scholar] [CrossRef] [Green Version]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA production and structure of gadB/gadC genes in Lactobacillus and Bifidobacterium strains from human microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef]
- Ko, C.Y.; Lin, H.T.V.; Tsai, G.J. Gamma-aminobutyric acid production in black soybean milk by Lactobacillus brevis FPA 3709 and the antidepressant effect of the fermented product on a forced swimming rat model. Process Biochem. 2013, 48, 559–568. [Google Scholar] [CrossRef]
- Kuwahara, A.; Matsuda, K.; Kuwahara, Y.; Asano, S.; Inui, T.; Marunaka, Y. Microbiota-gut-brain axis: Enteroendocrine cells and the enteric nervous system form an interface between the microbiota and the central nervous system. Biomed. Res. 2020, 41, 199–216. [Google Scholar] [CrossRef]
- Eisenhofer, G.; Aneman, A.; Friberg, P.; Hooper, D.; Fåndriks, L.; Lonroth, H.; Hunyady, B.; Mezey, E. Substantial production of dopamine in the human gastrointestinal tract. J. Clin. Endocrinol. Metab. 1997, 82, 3864–3871. [Google Scholar] [CrossRef]
- Strafella, A.P.; Paus, T.; Barrett, J.; Dagher, A. Repetitive transcranial magnetic stimulation of the human prefrontal cortex induces dopamine release in the caudate nucleus. J. Neurosci. 2001, 21, RC157. [Google Scholar] [CrossRef]
- Pascual-Leone, A.; Davey, N.; Rothwell, J.; Wassermann, E.; Puri, B. Handbook of Transcranial Magnetic Stimulation; Arnold: London, UK, 2002. [Google Scholar]
- Pell, G.S.; Roth, Y.; Zangen, A. Modulation of cortical excitability induced by repetitive transcranial magnetic stimulation: Influence of timing and geometrical parameters and underlying mechanisms. Prog. Neurobiol. 2011, 93, 59–98. [Google Scholar] [CrossRef]
- Terraneo, A.; Leggio, L.; Saladini, M.; Ermani, M.; Bonci, A.; Gallimberti, L. Transcranial magnetic stimulation of dorsolateral prefrontal cortex reduces cocaine use: A pilot study. Eur. Neuropsychopharmacol. 2016, 26, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, H.I.; Paulus, W.; Batsikadze, G.; Jamil, A.; Kuo, M.F.; Nitsche, M.A. Acute and chronic effects of noradrenergic enhancement on transcranial direct current stimulation-induced neuroplasticity in humans. J. Physiol. 2017, 595, 1305–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrulli, A.; Macrì, C.; Terruzzi, I.; Ambrogi, F.; Milani, V.; Adamo, M.; Luzi, L. High frequency deep transcranial magnetic stimulation acutely increases β-endorphins in obese humans. Endocrine 2019, 64, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ferrulli, A.; Macrì, C.; Terruzzi, I.; Massarini, S.; Ambrogi, F.; Adamo, M.; Milani, V.; Luzi, L. Weight loss induced by deep transcranial magnetic stimulation in obesity: A randomized, double-blind, sham-controlled study. Diabetes. Obes. Metab. 2019, 21, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Ferrulli, A.; Massarini, S.; Macrì, C.; Luzi, L. Safety and tolerability of repeated sessions of deep transcranial magnetic stimulation in obesity. Endocrine 2021, 71, 331–343. [Google Scholar] [CrossRef]
- Devoto, F.; Ferrulli, A.; Zapparoli, L.; Massarini, S.; Banfi, G.; Paulesu, E.; Luzi, L. Repetitive deep TMS for the reduction of body weight: Bimodal effect on the functional brain connectivity in “diabesity”. Nutr. Metab. Cardiovasc. Dis. 2021, in press. [Google Scholar] [CrossRef]
- Makovac, E.; Thayer, J.F.; Ottaviani, C. A meta-analysis of non-invasive brain stimulation and autonomic functioning: Implications for brain-heart pathways to cardiovascular disease. Neurosci. Biobehav. Rev. 2017, 74, 330–341. [Google Scholar] [CrossRef] [Green Version]
- Benarroch, E.E. The central autonomic network: Functional organization, dysfunction, and perspective. Mayo Clin. Proc. 1993, 68, 988–1001. [Google Scholar] [CrossRef]
- Artifon, M.; Schestatsky, P.; Griebler, N.; Tossi, G.M.; Beraldo, L.M.; Pietta-Dias, C. Effects of transcranial direct current stimulation on the gut microbiome: A case report. Brain Stimul. 2020, 13, 1451–1452. [Google Scholar] [CrossRef]
- Haney, M.M.; Ericsson, A.C.; Lever, T.E. Effects of Intraoperative Vagal Nerve Stimulation on the Gas-trointestinal Microbiome in a Mouse Model of Amyotrophic Lateral Sclerosis. Comp. Med. 2018, 68, 452–460. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA. 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Duranti, S.; Ferrario, C.; van Sinderen, D.; Ventura, M.; Turroni, F. Obesity and microbiota: An example of an intricate relationship. Genes Nutr. 2017, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Million, M.; Maraninchi, M.; Henry, M.; Armougom, F.; Richet, H.; Carrieri, P.; Valero, R.; Raccah, D.; Vialettes, B.; Raoult, D. Obesity-associated gut microbiota is enriched in Lactobacillus reuteri and depleted in Bifidobacterium animalis and Methanobrevibacter smithii. Int. J. Obes. 2012, 36, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.J.; Gerasimidis, K.; Edwards, C.A.; Shaikh, M.G. Role of Gut Microbiota in the Aetiology of Obesity: Proposed Mechanisms and Review of the Literature. J. Obes. 2016, 2016, 7353642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Huang, Z.P.; Liu, C.Q.; Qi, L.; Sheng, Y.; Zou, D.J. Modulation of the gut microbiome: A systematic review of the effect of bariatric surgery. Eur. J. Endocrinol. 2018, 178, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Sivamaruthi, B.S.; Kesika, P.; Suganthy, N.; Chaiyasut, C. A Review on Role of Microbiome in Obesity and Antiobesity Properties of Probiotic Supplements. BioMed Res. Int. 2019, 2019, 3291367. [Google Scholar] [CrossRef]
- Louis, S.; Tappu, R.M.; Damms-Machado, A.; Huson, D.H.; Bischoff, S.C. Characterization of the Gut Microbial Community of Obese Patients Following a Weight-Loss Intervention Using Whole Metagenome Shotgun Sequencing. PLoS ONE 2016, 11, e0149564. [Google Scholar] [CrossRef]
- Brahe, L.K.; Le Chatelier, E.; Prifti, E.; Pons, N.; Kennedy, S.; Hansen, T.; Pedersen, O.; Astrup, A.; Ehrlich, S.D.; Larsen, L.H. Specific gut microbiota features and metabolic markers in postmenopausal women with obesity. Nutr. Diabetes 2015, 5, e159. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Hijona, E.; Aguirre, L.; Milagro, F.I.; Bujanda, L.; Rimando, A.M.; Martínez, J.A.; Portillo, M.P. Pterostilbene-induced changes in gut microbiota composition in relation to obesity. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Fernández-Navarro, T.; Salazar, N.; Gutiérrez-Díaz, I.; de Los Reyes-Gavilán, C.G.; Gueimonde, M.; González, S. Different Intestinal Microbial Profile in Over-Weight and Obese Subjects Consuming a Diet with Low Content of Fiber and Antioxidants. Nutrients 2017, 9, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadal, I.; Santacruz, A.; Marcos, A.; Warnberg, J.; Garagorri, J.M.; Moreno, L.A.; Martin-Matillas, M.; Campoy, C.; Martí, A.; Moleres, A.; et al. Shifts in clostridia, bacteroides and immunoglobulin-coating fecal bacteria associated with weight loss in obese adolescents. Int. J. Obes. 2009, 33, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.; Burke, B.; Ford, D.; Garvin, G.; Korn, C.; Sulis, C.; Bhadelia, N. Possible association between obesity and Clostridium difficile infection. Emerg. Infect. Dis. 2013, 19, 1791–1798. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marlow, G.; Ellett, S.; Ferguson, I.R.; Zhu, S.; Karunasinghe, N.; Jesuthasan, A.C.; Han, D.Y.; Fraser, A.G.; Ferguson, L.R. Transcriptomics to study the effect of a Mediterranean-inspired diet on inflammation in Crohn’s disease patients. Hum. Genom. 2013, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.G.; Erbil, R.; Greve, J.W.; Buurman, W.A.; de Vos, W.M.; Rensen, S.S. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obesity 2013, 21, E607–E615. [Google Scholar] [CrossRef]
- Furet, J.P.; Kong, L.C.; Tap, J.; Poitou, C.; Basdevant, A.; Bouillot, J.L.; Mariat, D.; Corthier, G.; Doré, J.; Henegar, C.; et al. Differential adaptation of human gut microbiota to bariatric surgery-induced weight loss: Links with metabolic and lowgrade inflammation markers. Diabetes 2010, 59, 3049–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, D.; Moran, C.; Shanahan, F. The microbiota in inflammatory bowel disease. J. Gastroenterol. 2015, 50, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Andoh, A.; Nishida, A.; Takahashi, K.; Inatomi, O.; Imaeda, H.; Bamba, S.; Kito, K.; Sugimoto, M.; Kobayashi, T. Comparison of the gut microbial community between obese and lean peoples using 16S gene sequencing in a Japanese population. J. Clin. Biochem. Nutr. 2016, 59, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.F.; Murphy, E.F.; Nilaweera, K.; Ross, P.R.; Shanahan, F.; O’Toole, P.W.; Cotter, P.D. The gut microbiota and its relationship to diet and obesity: New insights. Gut Microbes 2012, 3, 186–202. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; García-Fuentes, E.; Cardona, F.; Queipo-Ortuño, M.I.; Tinahones, F.J. Insulin resistance is associated with specific gut microbiota in appendix samples from morbidly obese patients. Am. J. Transl. Res. 2016, 8, 5672–5684. [Google Scholar]
- Naderpoor, N.; Mousa, A.; Gomez-Arango, L.F.; Barrett, H.L.; Dekker Nitert, M.; de Courten, B. Faecal Microbiota Are Related to Insulin Sensitivity and Secretion in Overweight or Obese Adults. J. Clin. Med. 2019, 8, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, Y.; Motohashi, N.; Ito, H.; Ito, S.; Takano, A.; Nishikawa, T.; Suhara, T. Effects of repetitive transcranial magnetic stimulation on [11C]raclopride binding and cognitive function in patients with depression. J. Affect. Disord. 2006, 95, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Addolorato, G.; Antonelli, M.; Cocciolillo, F.; Vassallo, G.A.; Tarli, C.; Sestito, L.; Mirijello, A.; Ferrulli, A.; Pizzuto, D.A.; Camardese, G.; et al. Deep Transcranial Magnetic Stimulation of the Dorsolateral Prefrontal Cortex in Alcohol Use Disorder Patients: Effects on Dopamine Transporter Availability and Alcohol Intake. Eur. Neuropsychopharmacol. 2017, 27, 450–461. [Google Scholar] [CrossRef]
- Ikeda, T.; Kurosawa, M.; Uchikawa, C.; Kitayama, S.; Nukina, N. Modulation of monoamine transporter expression and function by repetitive transcranial magnetic stimulation. Biochem. Biophys. Res. Commun. 2005, 327, 218–224. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J. Cell Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef] [Green Version]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Haro, C.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Landa, B.B.; Navas-Cortés, J.A.; Tena-Sempere, M.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE 2016, 11, e0154090. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, M.; Pang, X.; Zhao, Y.; Wang, L.; Zhao, L. Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 2012, 6, 1848–1857. [Google Scholar] [CrossRef]
- Dinur-Klein, L.; Dannon, P.; Hadar, A.; Rosenberg, O.; Roth, Y.; Kotler, M.; Zangen, A. Smoking cessation induced by deep repetitive transcranial mag-netic stimulation of the prefrontal and insular cortices: A prospective, randomized controlled trial. Biol. Psychiatry 2014, 76, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Zangen, A.; Roth, Y.; Voller, B.; Hallett, M. Transcranial magnetic stimulation of deep brain regions: Evidence for efficacy of the H-coil. Clin. Neurophysiol. 2005, 116, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Roth, Y.; Amir, A.; Levkovitz, Y.; Zangen, A. Three-dimensional distribution of the electric field induced in the brain by transcranial magnetic stimulation using figure-8 and deep H-coils. J. Clin. Neurophysiol. 2007, 24, 31–38. [Google Scholar] [CrossRef]
- Bailey, B.W.; Tucker, L.A.; Peterson, T.R.; LeCheminant, J.D. Test-retest reliability of body fat percentage results using dual energy X-ray absorptiometry and the Bod Pod. Med. Sci. Sports Exerc. 2001, 33, S174. [Google Scholar] [CrossRef]
- Maddalozzo, G.F.; Cardinal, B.J.; Snow, C.M. Concurrent validity of the BOD POD and dual energy x-rayabsorptiometry techniques for assessing body composition in young women. J. Am. Diet. Assoc. 2002, 102, 1677–1679. [Google Scholar] [CrossRef]
- Haugen, H.A.; Chan, L.N.; Li, F. Indirect calorimetry: A practical guide for clinicians. Nutr. Clin. Pract. 2007, 22, 377–388. [Google Scholar] [CrossRef]
- Massarini, S.; Ferrulli, A.; Ambrogi, F.; Macrì, C.; Terruzzi, I.; Benedini, S.; Luzi, L. Routine resting energy expenditure measurement increases effectiveness of dietary intervention in obesity. Acta Diabetol. 2018, 55, 75–85. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
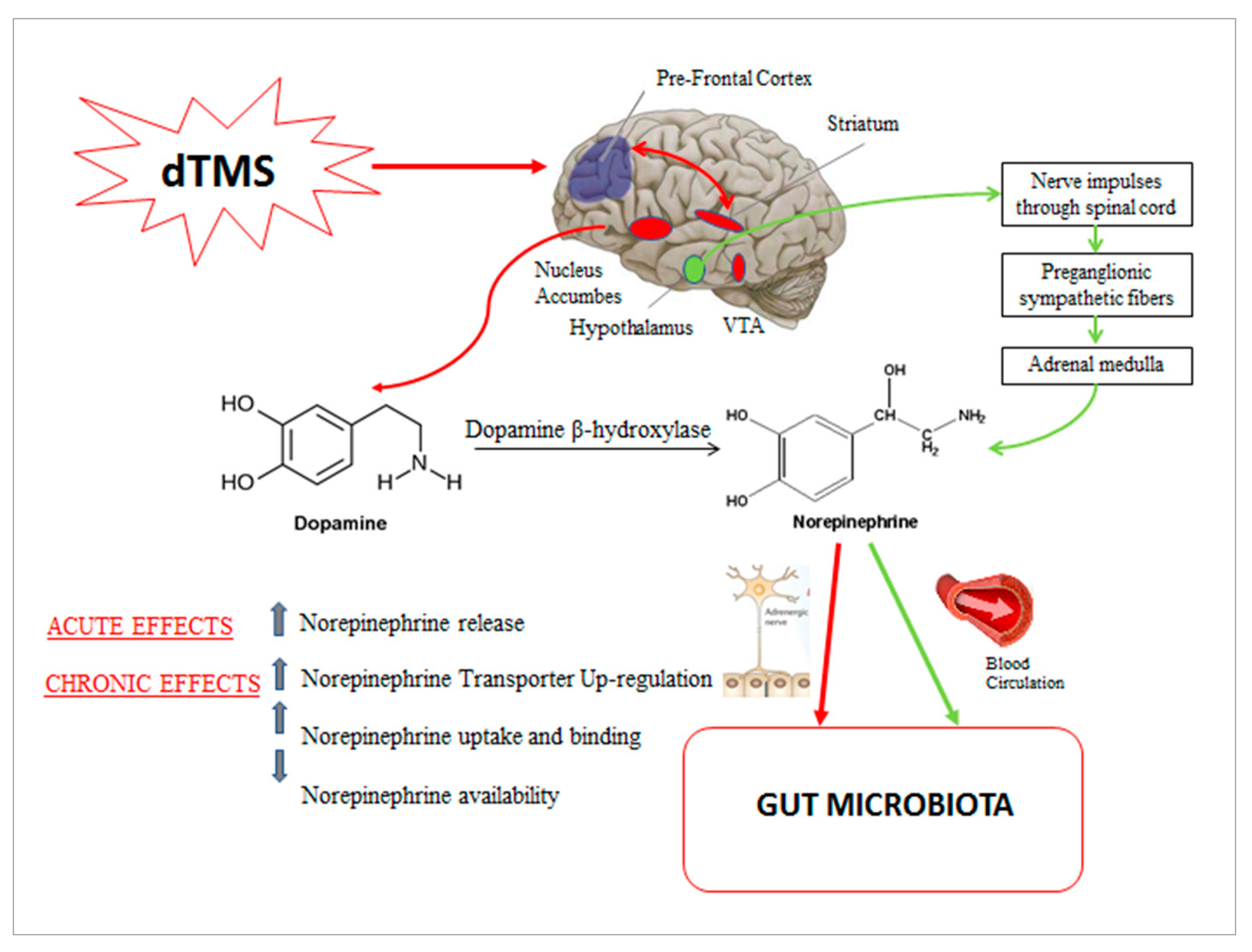{kind=link}
| Unit of Measure | HF (n = 9) | LF (n = 6) | Sham (n = 7) | p-Value | |
|---|---|---|---|---|---|
| Age, years | Years | 44.2 ± 3.5 | 41.8 ± 4.5 | 48.4 ± 3.9 | 0.453 |
| Sex | Males/females | 3/6 | 1/5 | 1/6 | - |
| BMI | kg/m2 | 37.3 ± 2.0 | 40.7 ± 2.1 | 35.5 ± 1.0 | 0.453 |
| FM | % | 46.4 ± 2.1 | 49.6 ± 2.5 | 46.3 ± 2.0 | 0.711 |
| REE | % | 101.0 ± 3.0 | 94.0 ± 6.2 | 93.1 ± 5.0 | 0.189 |
| RQ | 0.89 ± 0.02 | 0.89 ± 0.02 | 0.85 ± 0.03 | 0.144 | |
| Glucose | mg/dL | 95.0 ± 5.9 | 103.2 ± 14.9 | 95.9 ± 6.5 | 0.940 |
| Insulin | µU/mL | 27.1 ± 8.1 | 21.6 ± 2.6 | 20.1 ± 6.7 | 0.740 |
| HOMA-IR | - | 7.2 ± 2.5 | 5.6 ± 1.1 | 4.8 ± 1.7 | 0.798 |
| Glucagon | pg/mL | 40.7 ± 3.7 | 34.4 ± 2.2 | 43.2 ± 4.9 | 0.072 |
| Cholesterol | mg/dL | 208.9 ± 11.3 | 179.0 ± 16.4 | 189.0 ± 7.2 | 0.995 |
| Triglycerides | mg/dL | 146.1 ± 24.7 | 143.7 ± 35.1 | 103.1 ± 17.1 | 0.836 |
| Fructosamine | µmol/L | 243.9 ± 10.0 | 221.7 ± 8.8 | 234.1 ± 14.1 | 0.842 |
| Glycated hemoglobin | mmol/mol | 36.9 ± 1.5 | 41.0 ± 7.1 | 34.8 ± 2.8 | 0.235 |
| TSH | µUI/mL | 2.4 ± 0.3 | 2.38 ± 0.7 | 3.2 ± 0.7 | 0.354 |
| Cortisol | µg/dL | 0.43 ± 0.06 | 0.40 ± 0.06 | 0.35 ± 0.05 | 0.151 |
| Ghrelin | ng/mL | 13.1 ± 3.5 | 13.0 ± 3.6 | 7.9 ± 2.9 | 0.316 |
| Leptin | ng/mL | 64.7 ± 13.1 | 83.4 ± 22.5 | 108.3 ± 49.9 | 0.642 |
| Epinephrine | pg/mL | 1356.0 ± 261.8 | 996.7 ± 151.1 | 1487.7 ± 446.2 | 0.642 |
| Norepinephrine | ng/mL | 4.7 ± 0.8 | 4.7 ± 0.9 | 4.2 ± 0.9 | 0.400 |
| β-endorphin | ng/mL | 0.708 ± 0.12 | 0.742 ± 0.21 | 0.549 ± 0.09 | 0.878 |
| Norepinephrine (ng/mL) | |||||||
|---|---|---|---|---|---|---|---|
| T0 | T1 | T0 vs. T1 | T2 | T3 | T2 vs. T3 | T0 vs. T2 | |
| HF Group | 4.7 ± 0.8 | 5.7 ± 0.7 | * 0.046 | 1.8 ± 0.7 | 1.7 ± 0.6 | 0.250 | ** 0.007 |
| LF Group | 4.7 ± 0.9 | 4.0 ± 1.2 | 0.261 | 3.1 ± 0.7 | 2.2 ± 0.4 | 0.259 | * 0.041 |
| Sham Group | 4.2 ± 0.9 | 3.6 ± 1.4 | 0.465 | 3.4 ± 1.5 | 4.3 ± 1.6 | 0.578 | 0.473 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrulli, A.; Drago, L.; Gandini, S.; Massarini, S.; Bellerba, F.; Senesi, P.; Terruzzi, I.; Luzi, L. Deep Transcranial Magnetic Stimulation Affects Gut Microbiota Composition in Obesity: Results of Randomized Clinical Trial. Int. J. Mol. Sci. 2021, 22, 4692. https://doi.org/10.3390/ijms22094692
Ferrulli A, Drago L, Gandini S, Massarini S, Bellerba F, Senesi P, Terruzzi I, Luzi L. Deep Transcranial Magnetic Stimulation Affects Gut Microbiota Composition in Obesity: Results of Randomized Clinical Trial. International Journal of Molecular Sciences. 2021; 22(9):4692. https://doi.org/10.3390/ijms22094692
Chicago/Turabian StyleFerrulli, Anna, Lorenzo Drago, Sara Gandini, Stefano Massarini, Federica Bellerba, Pamela Senesi, Ileana Terruzzi, and Livio Luzi. 2021. "Deep Transcranial Magnetic Stimulation Affects Gut Microbiota Composition in Obesity: Results of Randomized Clinical Trial" International Journal of Molecular Sciences 22, no. 9: 4692. https://doi.org/10.3390/ijms22094692
APA StyleFerrulli, A., Drago, L., Gandini, S., Massarini, S., Bellerba, F., Senesi, P., Terruzzi, I., & Luzi, L. (2021). Deep Transcranial Magnetic Stimulation Affects Gut Microbiota Composition in Obesity: Results of Randomized Clinical Trial. International Journal of Molecular Sciences, 22(9), 4692. https://doi.org/10.3390/ijms22094692