
Acid-Sensing Ion Channels and Mechanosensation



Abstract
:1. Introduction
2. ASIC-Associated Pathologies
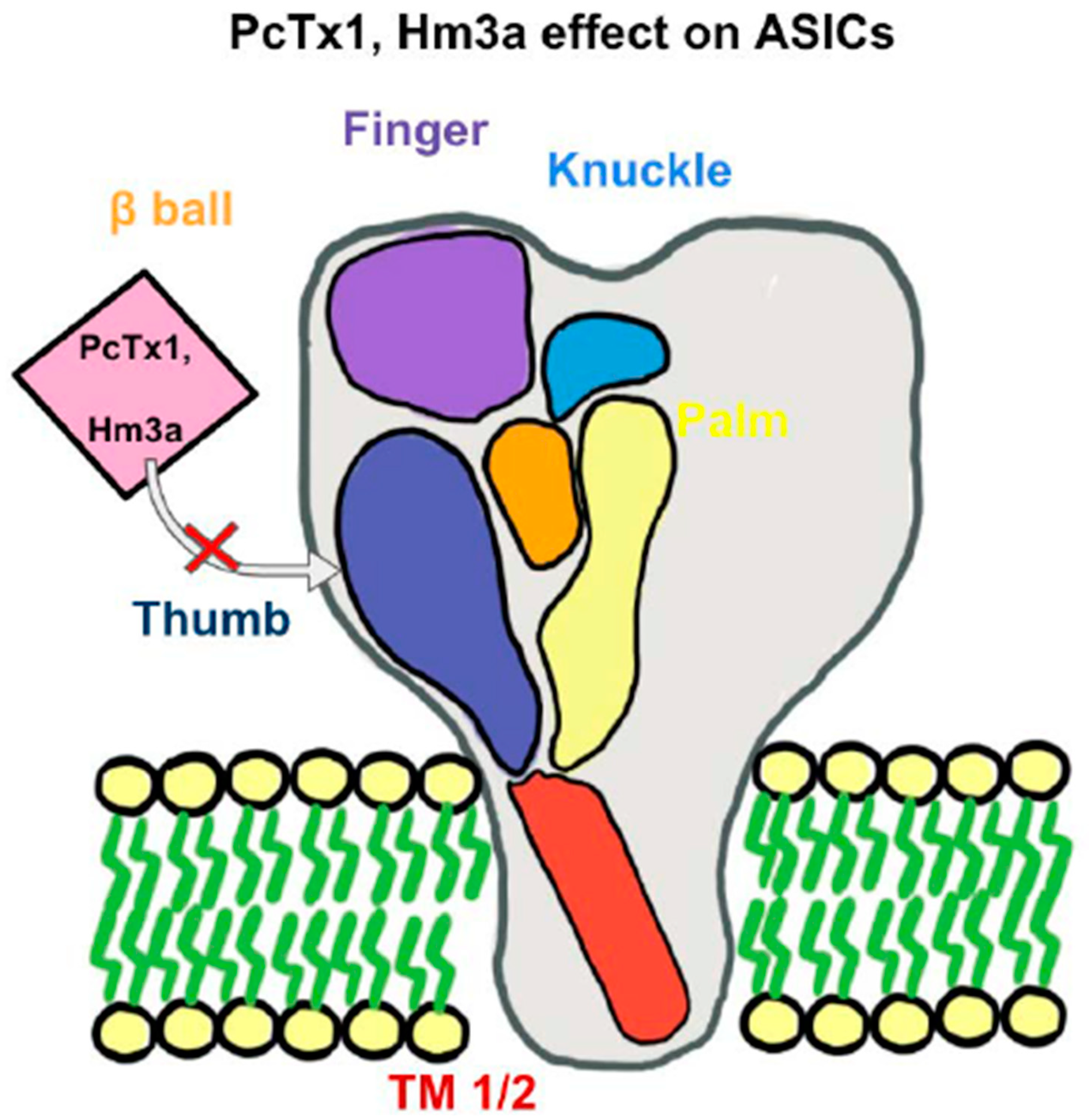
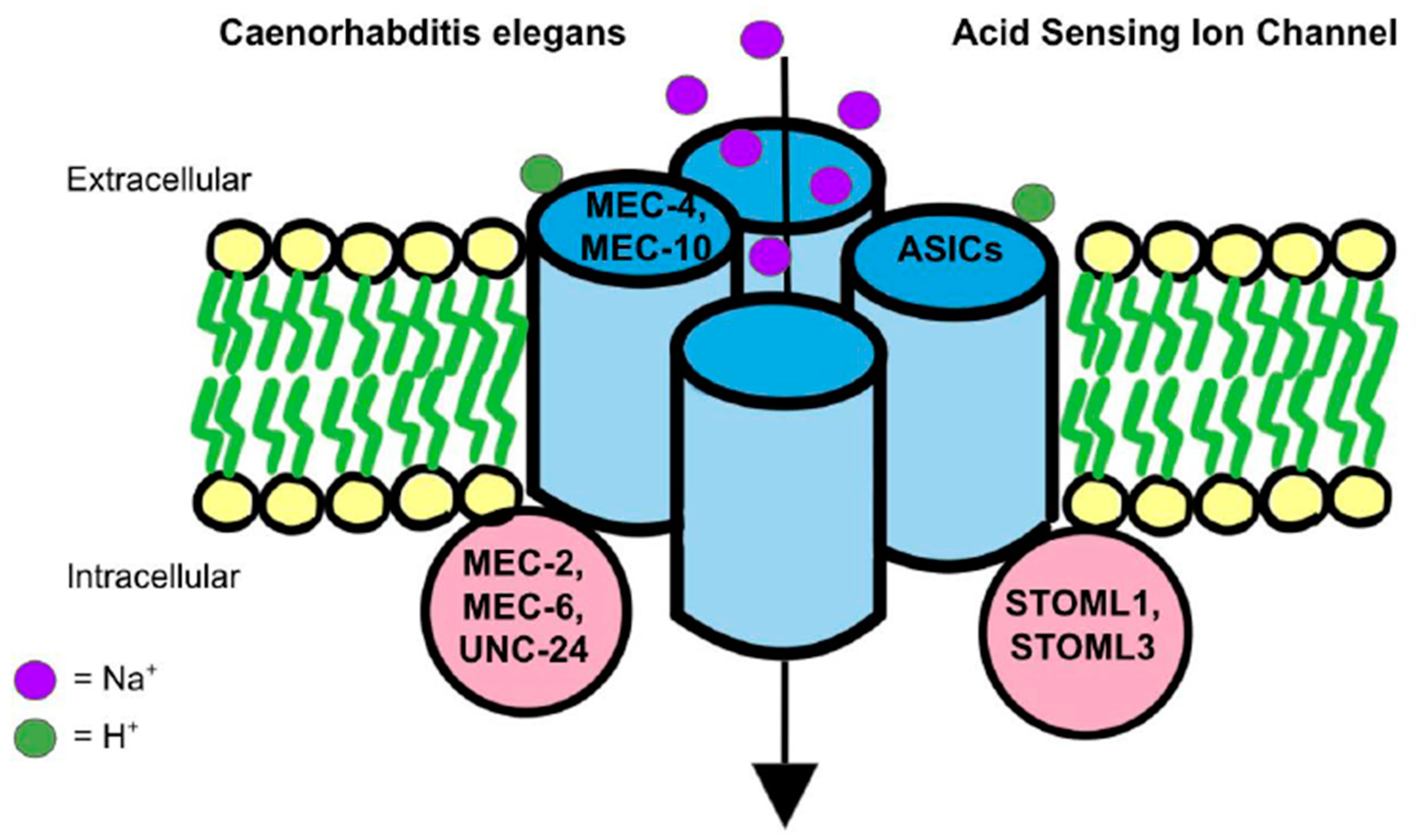
3. ASICs in Mechanosensation
4. Perspective for Future Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| β | beta |
| ASIC | acid-sensing ion channel |
| ASICs | acid sensing ion channels |
| STOML | stomatin-like |
| AD | Alzheimer’s disease |
| MS | multiple sclerosis |
| NMDA | N-Methyl-D-aspartic acid |
| Ca2+ | calcium |
| Na+ | sodium |
| H+ | proton |
| PcTx1 | Psalmotoxin 1 |
| MEC | mechanosensory abnormality |
| DRG | dorsal root ganglion |
| mGlu | metabotropic glutamate |
References
- Waldmann, R.; Champigny, G.; Bassilana, F.; Heurteaux, C.; Lazdunski, M. A proton-gated cation channel involved in acid-sensing. Nature 1997, 386, 173–177. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, Z.; Li, W.G.; Cao, H.; Feng, E.G.; Yu, F.; Liu, H.; Jiang, H.; Xu, T.L. A nonproton ligand sensor in the acid-sensing ion channel. Neuron 2010, 68, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellenberger, S.; Schild, L. International Union of Basic and Clinical Pharmacology. XCI. Structure, function, and pharmacology of acid-sensing ion channels and the epithelial Na+ channel. Pharmacol. Rev. 2015, 67, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.R.; Jiang, B.Y.; Chen, C.C. Acid-sensing ion channels: Dual function proteins for chemo-sensing and mechano-sensing. J. Biomed. Sci. 2018, 25, 46. [Google Scholar] [CrossRef] [PubMed]
- Gründer, S.; Chen, X. Structure, function, and pharmacology of acid-sensing ion channels (ASICs): Focus on ASIC1a. Int. J. Physiol. Pathophysiol. Pharmacol. 2010, 2, 73–94. [Google Scholar] [PubMed]
- Yoder, N.; Yoshioka, C.; Gouaux, E. Gating mechanisms of acid-sensing ion channels. Nature 2018, 555, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Rook, M.L.; Musgaard, M.; MacLean, D.M. Coupling structure with function in acid-sensing ion channels: Challenges in pursuit of proton sensors. J. Physiol. 2021, 599, 417–430. [Google Scholar] [CrossRef]
- Jasti, J.; Furukawa, H.; Gonzales, E.B.; Gouaux, E. Structure of acid-sensing ion channel 1 at 1.9 A resolution and low pH. Nature 2007, 449, 316–323. [Google Scholar] [CrossRef]
- Zeng, W.Z.; Liu, D.S.; Xu, T.L. Acid-sensing ion channels: Trafficking and pathophysiology. Channels 2014, 8, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Yermolaieva., O.; Leonard, A.S.; Schnizler, M.K.; Abboud, F.M.; Welsh, M.J. Extracellular acidosis increases neuronal cell calcium by activating acid-sensing ion channel 1a. Proc. Natl. Acad. Sci. USA 2004, 101, 6752–6757. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Z.; Zeng, W.Z.; Xiao, X.; Huang, Y.; Song, X.L.; Yu, Z.; Tang, D.; Dong, X.P.; Zhu, M.X.; Xu, T.L. Intracellular ASIC1a regulates mitochondrial permeability transition-dependent neuronal death. Cell Death Differ. 2013, 20, 1359–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.P.; Gong, H.; Parsons, M.G.; Kundert, J.R.; Reznikov, L.R.; Bernardinelli, L.; Chaloner, K.; Buchanan, G.F.; Wemmie, J.A.; Richerson, G.B.; et al. Localization and behaviors in null mice suggest that ASIC1 and ASIC2 modulate responses to aversive stimuli. Genes Brain Behav. 2014, 13, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Xu, Y.; Jiang, Y.Q.; Xu, J.; Hu, Y.; Zha, X.M. ASIC subunit ratio and differential surface trafficking in the brain. Mol. Brain 2016, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Taugher, R.J.; Lu, Y.; Fan, R.; Ghobbeh, A.; Kreple, C.J.; Faraci, F.M.; Wemmie, J.A. ASIC1A in neurons is critical for fear-related behaviors. Genes Brain Behav. 2017, 16, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Price, M.P.; Taugher, R.J.; Grigsby, D.; Ash, J.J.; Stark, A.C.; Saad, M.Z.; Singh, K.; Mandal, J.; Wemmie, J.A.; et al. Transient acidosis while retrieving a fear-related memory enhances its lability. Elife 2017, 6, e22564. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Q.; Song, X.L.; Jiang, Q.; Wu, Y.J.; Li, Y.; Yuan, T.F.; Zhang, S.; Xu, N.J.; Zhu, M.X.; et al. Fear extinction requires ASIC1a-dependent regulation of hippocampal-prefrontal correlates. Sci. Adv. 2018, 4, eaau3075. [Google Scholar] [CrossRef] [Green Version]
- Wemmie, J.A.; Chen, J.; Askwith, C.C.; Hruska-Hageman, A.M.; Price, M.P.; Nolan, B.C.; Yoder, P.G.; Lamani, E.; Hoshi, T.; Freeman, J.H., Jr.; et al. Welsh MJ. The acid-activated ion channel ASIC contributes to synaptic plasticity, learning, and memory. Neuron 2002, 34, 463–477. [Google Scholar] [CrossRef] [Green Version]
- Li, W.G.; Liu, M.G.; Deng, S.; Liu, Y.M.; Shang, L.; Ding, J.; Hsu, T.T.; Jiang, Q.; Li, Y.; Li, F.; et al. ASIC1a regulates insular long-term depression and is required for the extinction of conditioned taste aversion. Nat. Commun. 2016, 7, 13770. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.P.; Papasian, C.J.; Wang, J.Q.; Xiong, Z.G. Modulation of acid-sensing ion channels: Molecular mechanisms and therapeutic potential. Int. J. Physiol. Pathophysiol. Pharmacol. 2011, 3, 288–309. [Google Scholar]
- Ma, C.L.; Sun, H.; Yang, L.; Wang, X.T.; Gao, S.; Chen, X.W.; Ma, Z.Y.; Wang, G.H.; Shi, Z.; Zheng, Q.Y. Acid-sensing ion channel 1a modulates NMDA receptor function through targeting NR1/NR2A/NR2B triheteromeric receptors. Neuroscience 2019, 406, 389–404. [Google Scholar] [CrossRef]
- Herbert, L.M.; Resta, T.C.; Jernigan, N.L. RhoA increases ASIC1a plasma membrane localization and calcium influx in pulmonary arterial smooth muscle cells following chronic hypoxia. Am. J. Physiol. Cell. Physiol. 2018, 314, C166–C176. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.M.; Herbert, L.M.; Walker, B.R.; Resta, T.C.; Jernigan, N.L. Coupling of store-operated calcium entry to vasoconstriction is acid-sensing ion channel 1a dependent in pulmonary but not mesenteric arteries. PLoS ONE 2020, 15, e0236288. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.T.; Fong, S.W.; Lee, C.H.; Lin, S.H.; Chen, C.C. Involvement of acid-sensing ion channel 1b in the development of acid-induced chronic muscle pain. Front. Neurosci. 2019, 13, 1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofori-Armstrong, B.; Budusan, E.; Rash, L.D. Mambalgin-3 potentiates human acid-sensing ion channel 1b under mild to moderate acidosis: Implications as an analgesic lead. Proc. Natl. Acad. Sci. USA 2021, 118, e2021581118. [Google Scholar] [CrossRef]
- Lee, J.Y.; Saez, N.J.; Cristofori-Armstrong, B.; Anangi, R.; King, G.F.; Smith, M.T.; Rash, L.D. Inhibition of acid-sensing ion channels by diminazene and APETx2 evoke partial and highly variable antihyperalgesia in a rat model of inflammatory pain. Br. J. Pharmacol. 2018, 175, 2204–2218. [Google Scholar] [CrossRef] [PubMed]
- Ugawa, S.; Yamamoto, T.; Ueda, T.; Ishida, Y.; Inagaki, A.; Nishigaki, M.; Shimada, S. Amiloride-insensitive currents of the acid-sensing ion channel-2a (ASIC2a)/ASIC2b heteromeric sour-taste receptor channel. J. Neurosci. 2003, 23, 3616–3622. [Google Scholar] [CrossRef] [Green Version]
- Harding, A.M.; Kusama, N.; Hattori, T.; Gautam, M.; Benson, C.J. ASIC2 subunits facilitate expression at the cell surface and confer regulation by PSD-95. PLoS ONE 2014, 9, e93797. [Google Scholar] [CrossRef]
- Jiang, N.; Wu, J.; Leng, T.; Yang, T.; Zhou, Y.; Jiang, Q.; Wang, B.; Hu, Y.; Ji, Y.H.; Simon, R.P.; et al. Region specific contribution of ASIC2 to acidosis-and ischemia-induced neuronal injury. J. Cereb. Blood Flow Metab. 2017, 37, 528–540. [Google Scholar] [CrossRef]
- Yin, T.; Lindley, T.E.; Albert, G.W.; Ahmed, R.; Schmeiser, P.B.; Grady, M.S.; Howard, M.A.; Welsh, M.J. Loss of Acid sensing ion channel-1a and bicarbonate administration attenuate the severity of traumatic brain injury. PLoS ONE 2013, 8, e72379. [Google Scholar] [CrossRef]
- Koehn, L.M.; Noor, N.M.; Dong, Q.; Er, S.Y.; Rash, L.D.; King, G.F.; Dziegielewska, K.M.; Saunders, N.R.; Habgood, M.D. Selective inhibition of ASIC1a confers functional and morphological neuroprotection following traumatic spinal cord injury. F1000 Res. 2016, 5, 1822. [Google Scholar] [CrossRef]
- Komnig, D.; Imgrund, S.; Reich, A.; Gründer, S.; Falkenburger, B.H. ASIC1a deficient mice show unaltered neurodegeneration in the subacute MPTP model of Parkinson disease. PLoS ONE 2016, 11, e0165235. [Google Scholar] [CrossRef]
- Mango, D.; Nisticò, R. Role of ASIC1a in Aβ-induced synaptic alterations in the hippocampus. Pharmacol. Res. 2018, 131, 61–65. [Google Scholar] [CrossRef]
- Zhou, R.; Leng, T.; Yang, T.; Chen, F.; Hu, W.; Xiong, Z.G. β-estradiol protects against acidosis-mediated and ischemic neuronal injury by promoting ASIC1a (acid-sensing ion channel 1a) protein degradation. Stroke 2019, 50, 2902–2911. [Google Scholar] [CrossRef]
- Detweiler, N.D.; Herbert, L.M.; Garcia, S.M.; Yan, S.; Vigil, K.G.; Sheak, J.R.; Resta, T.C.; Walker, B.R.; Jernigan, N.L. Loss of acid-sensing ion channel 2 enhances pulmonary vascular resistance and hypoxic pulmonary hypertension. J. Appl. Physiol. 2019, 127, 393–407. [Google Scholar] [CrossRef]
- Lu, Y.; Ma, X.; Sabharwal, R.; Snitsarev, V.; Morgan, D.; Rahmouni, K.; Drummond, H.A.; Whiteis, C.A.; Costa, V.; Price, M.; et al. The ion channel ASIC2 is required for baroreceptor and autonomic control of the circulation. Neuron 2009, 24, 885–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.P.; Bo, Y.; Qin, Z.; Ran, H.; Li, W.; Li, Y.F.; Ming, G. Expression of acid-sensing ion channels in renal tubular epithelial cells and their role in patients with henoch-schönlein purpura nephritis. Med. Sci. Monit. 2017, 23, 1916. [Google Scholar] [CrossRef] [Green Version]
- Gannon, K.P.; McKey, S.E.; Stec, D.E.; Drummond, H.A. Altered myogenic vasoconstriction and regulation of whole kidney blood flow in the ASIC2 knockout mouse. Am. J. Physiol. Renal. Physiol. 2015, 308, F339–F348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.P.; Ni, W.L.; Dai, B.B.; Wu, X.S.; Wang, Z.S.; Xie, Y.Y.; Wang, Z.Q.; Yang, W.J.; Ge, J.F.; Hu, W.; et al. ASIC2a overexpression enhances the protective effect of PcTx1 and APETx2 against acidosis-induced articular chondrocyte apoptosis and cytotoxicity. Gene 2018, 642, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kweon, H.J.; Lee, H.; Suh, B.C. Rapid resensitization of ASIC2a is conferred by three amino acid residues in the N terminus. J. Gen. Physiol. 2019, 151, 944–953. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, T.W.; Lee, K.G.; Gormley, M.G.; Askwith, C.C. Heteromeric acid-sensing ion channels (ASICs) composed of ASIC2b and ASIC1a display novel channel properties and contribute to acidosis-induced neuronal death. J. Neurosci. 2011, 31, 9723–9734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kweon, H.J.; Kim, D.I.; Bae, Y.; Park, J.Y.; Suh, B.C. Acid-sensing ion channel 2a (ASIC2a) promotes surface trafficking of ASIC2b via heteromeric assembly. Sci. Rep. 2016, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.L.; Cheng, S.J.; Lin, S.H.; Chuang, Y.C.; Huang, E.Y.; Chen, C.C. The effect of ASIC3 knockout on corticostriatal circuit and mouse self-grooming behavior. Front. Cell. Neurosci. 2019, 13, 86. [Google Scholar] [CrossRef] [Green Version]
- Kweon, H.J.; Cho, J.H.; Jang, I.S.; Suh, B.C. ASIC2a-dependent increase of ASIC3 surface expression enhances the sustained component of the currents. BMB Rep. 2016, 49, 542. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Peterson, A.M.; Chu, Y.; Yao, X.; Zha, X.M.; Chu, X.P. Histidine residues are responsible for bidirectional effects of zinc on acid-sensing ion channel 1a/3 heteromeric channels. Biomolecules 2020, 10, 1264. [Google Scholar] [CrossRef]
- Stephan, G.; Huang, L.; Tang, Y.; Vilotti, S.; Fabbretti, E.; Yu, Y.; Nörenberg, W.; Franke, H.; Gölöncsér, F.; Sperlágh, B.; et al. The ASIC3/P2X3 cognate receptor is a pain-relevant and ligand-gated cationic channel. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hiasa, M.; Okui, T.; Allette, Y.M.; Ripsch, M.S.; Sun-Wada, G.H.; Wakabayashi, H.; Roodman, G.D.; White, F.A.; Yoneda, T. Bone pain induced by multiple myeloma is reduced by targeting V-ATPase and ASIC3. Cancer Res. 2017, 77, 1283–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, L.T.; Hsieh, C.L.; Hsu, H.C.; Lin, Y.W. Targeting ASIC3 for relieving mice fibromyalgia pain: Roles of electroacupuncture, opioid, and adenosine. Sci. Rep. 2017, 7, 46663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzer, P. Acid-sensing ion channels in gastrointestinal function. Neuropharmacology 2015, 94, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Montalbetti, N.; Rooney, J.G.; Marciszyn, A.L.; Carattino, M.D. ASIC3 fine-tunes bladder sensory signaling. Am. J. Physiol. Renal. Physiol. 2018, 315, F870–F879. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Reznikov, L.R.; Welsh, M.J. Expression and activity of acid-sensing ion channels in the mouse anterior pituitary. PLoS ONE 2014, 9, e115310. [Google Scholar] [CrossRef] [PubMed]
- Hoshikawa, M.; Kato, A.; Hojo, H.; Shibata, Y.; Kumamoto, N.; Watanabe, M.; Ugawa, S. Distribution of ASIC4 transcripts in the adult wild-type mouse brain. Neurosci. Lett. 2017, 651, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Chien, Y.C.; Chiang, W.W.; Liu, Y.Z.; Lien, C.C.; Chen, C.C. Genetic mapping of ASIC 4 and contrasting phenotype to ASIC 1a in modulating innate fear and anxiety. Eur. J. Neurosci. 2015, 41, 1553–1568. [Google Scholar] [CrossRef]
- Schwartz, V.; Friedrich, K.; Polleichtner, G.; Gründer, S. Acid-sensing ion channel (ASIC) 4 predominantly localizes to an early endosome-related organelle upon heterologous expression. Sci. Rep. 2015, 5, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storozhuka, M.; Cherninskyia, A.; Maximyuka, O.; Isaeva, D.; Krishtala, O. Acid-sensing ion channels: Focus on physiological and some pathological roles in the brain. Curr. Neuropharmacol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Huang, T.J.; Wu, M.H.; Li, Y.Y.; Lee, K.D. High expression of acid-sensing ion channel 2 (asic2) in bone cells in osteoporotic vertebral fractures. Biomed. Res. Int. 2019, 2019, 4714279. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.C.; Chung, C.Y.; Liao, F.; Chen, C.C.; Lee, C.H. Peripheral sensory neuron injury contributes to neuropathic pain in experimental autoimmune encephalomyelitis. Sci. Rep. 2017, 7, 42304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergo, S.; Craner, M.J.; Etzensperger, R.; Attfield, K.; Friese, M.A.; Newcombe, J.; Esiri, M.; Fugger, L. Acid-sensing ion channel 1 is involved in both axonal injury and demyelination in multiple sclerosis and its animal model. Brain 2011, 134, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, A.E.; Schnizler, M.K.; Albert, G.W.; Severson, M.A.; Howard, I.M.A.; Welsh, M.J.; Wemmie, J.A. Seizure termination by acidosis depends on ASIC1a. Nat. Neurosci. 2008, 11, 816–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzuca, M.; Heurteaux, C.; Alloui, A.; Diochot, S.; Baron, A.; Voilley, N.; Blondeau, N.; Escoubas, P.; Gélot, A.; Cupo, A.; et al. A tarantula peptide against pain via ASIC1a channels and opioid mechanisms. Nat. Neurosci. 2007, 10, 943–945. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- Gutman, A.L.; Cosme, C.V.; Noterman, M.F.; Worth, W.R.; Wemmie, J.A.; LaLumiere, R.T. Overexpression of ASIC1A in the nucleus accumbens of rats potentiates cocaine-seeking behavior. Addict. Biol. 2018, 25, e12690. [Google Scholar] [CrossRef]
- Kreple, C.J.; Lu, Y.; LaLumiere, R.T.; Wemmie, J.A. Drug abuse and the simplest neurotransmitter. ACS Chem. Neurosci. 2014, 5, 746–748. [Google Scholar] [CrossRef]
- Zhang, G.C.; Mao, L.M.; Wang, J.Q.; Chu, X.P. Upregulation of acid-sensing ion channel 1 protein expression by chronic administration of cocaine in the mouse striatum in vivo. Neurosci. Lett. 2009, 459, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreple, C.J.; Lu, Y.; Taugher, R.J.; Schwager-Gutman, A.L.; Du, J.; Stump, M.; Wang, Y.; Ghobbeh, A.; Fan, R.; Cosme, C.V.; et al. Acid-sensing ion channels contribute to synaptic transmission and inhibit cocaine-evoked plasticity. Nat. Neurosci. 2014, 17, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wang, C.M.; Fibuch, E.E.; Wang, J.Q.; Chu, X.P. Differential regulation of locomotor activity to acute and chronic cocaine administration by acid-sensing ion channel 1a and 2 in adult mice. Neuroscience 2013, 246, 170–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, N.; Lu, Z.; Zhang, J.; Shi, Y.; Ning, Y.; Chen, J.; Jin, S.; Shen, B.; Fang, Y.; Zou, J.; et al. Acid-sensing ion channel 1a is involved in ischaemia/reperfusion induced kidney injury by increasing renal epithelia cell apoptosis. J. Cell. Mol. Med. 2019, 23, 3429–3440. [Google Scholar] [CrossRef]
- Xiong, Z.G.; Zhu, X.M.; Chu, X.P.; Minami, M.; Hey, J.; Wei, W.L.; MacDonald, J.F.; Wemmie, J.A.; Price, M.P.; Welsh, M.J.; et al. Neuroprotection in ischemia: Blocking calcium-permeable acid-sensing ion channels. Cell 2004, 118, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Z.; Wang, J.J.; Huang, Y.; Liu, F.; Zeng, W.Z.; Li, Y.; Xiong, Z.G.; Zhu, M.X.; Xu, T.L. Tissue acidosis induces neuronal necroptosis via ASIC1a channel independent of its ionic conduction. Elife 2015, 4, e05682. [Google Scholar] [CrossRef]
- William, M.; Turnadzic, S.; Chu, X.P. Commentary: Therapeutic potential of targeting the auto-inhibition of ASIC1a for neuroprotection against ischemic brain injury. Front. Pharmacol. 2020, 11, 1763. [Google Scholar] [CrossRef] [PubMed]
- Qiang, M.; Dong, X.; Zha, Z.; Zuo, X.K.; Song, X.L.; Zhao, L.; Yuan, C.; Huang, C.; Tao, P.; Hu, Q.; et al. Selection of an ASIC1a-blocking combinatorial antibody that protects cells from ischemic death. Proc. Natl. Acad. Sci. USA 2018, 115, E7469–E7477. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.; Jiang, Q.; Chu, X.P. Commentary: Potential therapeutic consequences of an acid-sensing ion channel 1a-blocking antibody. Front. Pharmacol. 2019, 10, 954. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, T.W.; Askwith, C.C. Dynorphin opioid peptides enhance acid-sensing ion channel 1a activity and acidosis-induced neuronal death. J. Neurosci. 2009, 29, 14371–14380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vick, J.S.; Askwith, C.C. ASICs and neuropeptides. Neuropharmacology 2015, 94, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Ramírez, A.; Vega, R.; Soto, E. Acid-sensing ion channels as potential therapeutic targets in neurodegeneration and neuroinflammation. Mediat. Inflamm. 2017, 2017, 3728096. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Xu, T.L. Acid-sensing ion channels as a target for neuroprotection: Acidotoxicity revisited. Sheng Li Xue Bao 2016, 68, 403–413. [Google Scholar]
- Fazia, T.; Pastorino, R.; Notartomaso, S.; Busceti, C.; Imbriglio, T.; Cannella, M.; Gentilini, D.; Morani, G.; Ticca, A.; Bitti, P.; et al. Acid sensing ion channel 2: A new potential player in the pathophysiology of multiple sclerosis. Eur. J. Neurosci. 2019, 49, 1233–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Cheng, X.Y.; Wang, F.; Liu, C.F. Acid-sensing ion channels: Potential therapeutic targets for neurologic diseases. Transl. Neurodegener. 2015, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arun, T.; Tomassini, V.; Sbardella, E.; De Ruiter, M.B.; Matthews, L.; Leite, M.I.; Gelineau-Morel, R.; Cavey, A.; Vergo, S.; Craner, M.; et al. Targeting ASIC1 in primary progressive multiple sclerosis: Evidence of neuroprotection with amiloride. Brain 2013, 136, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.P.; Wu, X.S.; Wang, Z.S.; Xie, Y.Y.; Ge, J.F.; Chen, F.H. Novel insights into acid-sensing ion channels: Implications for degenerative diseases. Aging Dis. 2015, 7, 491–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.W.; Hu, Z.L.; Ni, M.; Fang, P.; Zhang, P.W.; Shu, Q.; Fan, H.; Zhou, H.Y.; Ni, L.; Zhu, L.Q.; et al. Acid-sensing ion channels promote the inflammation and migration of cultured rat microglia. Glia 2015, 63, 483–496. [Google Scholar] [CrossRef]
- Karsan, N.; Gonzales, E.B.; Dussor, G. Targeted acid-sensing ion channel therapies for migraine. Neurotherapeutics 2018, 15, 402–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikhonova, T.B.; Nagaeva, E.I.; Barygin, O.I.; Potapieva, N.M.; Bolshakov, K.V.; Tikhonov, D.B. Monoamine NMDA receptor channel blockers inhibit and potentiate native and recombinant proton-gated ion channels. Neuropharmacology 2015, 89, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cristofori-Armstrong, B.; Rash, L.D. Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venoms. Neuropharmacology 2017, 127, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Vullo, S.; Kellenberger, S. A molecular view of the function and pharmacology of acid-sensing ion channels. Pharmacol. Res. 2020, 154, 104166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristofori-Armstrong, B.; Saez, N.J.; Chassagnon, I.R.; King, G.F.; Rash, L.D. The modulation of acid-sensing ion channel 1 by PcTx1 is pH-, subtype- and species-dependent: Importance of interactions at the channel subunit interface and potential for engineering selective analogues. Biochem. Pharmacol. 2019, 163, 381–390. [Google Scholar] [CrossRef]
- Er, S.Y.; Cristofori-Armstrong, B.; Escoubas, P.; Rash, L.D. Discovery and molecular interaction studies of a highly stable, tarantula peptide modulator of acid-sensing ion channel 1. Neuropharmacology 2017, 127, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Rossetti, G.; Joussen, S.; Gründer, S. Diminazene is a slow pore blocker of acid-sensing ion channel 1a (ASIC1a). Mol. Pharmacol. 2017, 92, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Liu, S.; Li, S.; Zhang, M.; Yang, F.; Wen, M.; Shi, P.; Wang, T.; Pan, M.; Chang, S.; et al. Structural insights into human acid-sensing ion channel 1a inhibition by snake toxin mambalgin1. Elife 2020, 9, e57096. [Google Scholar] [CrossRef]
- Andreev, Y.A.; Osmakov, D.I.; Koshelev, S.G.; Maleeva, E.E.; Logashina, Y.A.; Palikov, V.A.; Palikova, Y.A.; Dyachenko, I.A.; Kozlov, S.A. Analgesic activity of acid-sensing ion channel 3 (ASIC3) inhibitors: Sea anemones peptides Ugr9-1 and APETx2 versus low molecular weight compounds. Mar. Drugs 2018, 16, 500. [Google Scholar] [CrossRef] [Green Version]
- Yen, L.T.; Hsieh, C.L.; Hsu, H.C.; Lin, Y.W. Preventing the induction of acid saline-induced fibromyalgia pain in mice by electroacupuncture or APETx2 injection. Acupunct. Med. 2020, 38, 188–193. [Google Scholar] [CrossRef]
- Dube, G.R.; Lehto, S.G.; Breese, N.M.; Baker, S.J.; Wang, X.; Matulenko, M.A.; Honoré, P.; Stewart, A.O.; Moreland, R.B.; Brioni, J.D. Electrophysiological and in vivo characterization of A-317567, a novel blocker of acid sensing ion channels. Pain 2005, 117, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, M.; Kobayashi, H.; Takeda, M.; Araki, I. Blockade of acid-sensing ion channels increases urinary bladder capacity with or without intravesical irritation in mice. Front. Physiol. 2020, 11, 592867. [Google Scholar] [CrossRef] [PubMed]
- Alcaino, C.; Farrugia, G.; Beyder, A. Mechanosensitive piezo channels in the gastrointestinal tract. Curr. Top. Membr. 2017, 79, 219–244. [Google Scholar] [PubMed] [Green Version]
- Abraira, V.E.; Ginty, D.D. The sensory neurons of touch. Neuron 2013, 79, 618–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Bharill, S.; Isacoff, E.Y.; Chalfie, M. Subunit composition of a DEG/ENaC mechanosensory channel of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2015, 112, 11690–11695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hagan, R.; Chalfie, M.; Goodman, M.B. The MEC-4 DEG/ENaC channel of Caenorhabditis elegans touch receptor neurons transduces mechanical signals. Nat. Neurosci. 2005, 8, 43–50. [Google Scholar] [CrossRef]
- Goodman, M.B.; Ernstrom, G.G.; Chelur, D.S.; O’Hagan, R.; Yao, C.A.; Chalfie, M. MEC-2 regulates C. elegans DEG/ENaC channels needed for mechanosensation. Nature 2002, 415, 1039–1042. [Google Scholar] [CrossRef]
- Huang, M.; Chalfie, M. Gene interactions affecting mechanosensory transduction in Caenorhabditis elegans. Nature 1994, 367, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Emtage, L.; Gu, G.; Hartwieg, E.; Chalfie, M. Extracellular proteins organize the mechanosensory channel complex in C. elegans touch receptor neurons. Neuron 2004, 44, 795–807. [Google Scholar] [CrossRef] [Green Version]
- Árnadóttir, J.; O’Hagan, R.; Chen, Y.; Goodman, M.B.; Chalfie, M. The DEG/ENaC protein MEC-10 regulates the transduction channel complex in Caenorhabditis elegans touch receptor neurons. J. Neurosci. 2011, 31, 12695–12704. [Google Scholar] [CrossRef]
- Kozlenkov, A.; Lapatsina, L.; Lewin, G.R.; Smith, E.S. Subunit-specific inhibition of acid sensing ion channels by stomatin-like protein 1. J. Physiol. 2014, 592, 557–569. [Google Scholar] [CrossRef]
- Klipp, R.C.; Cullinan, M.M.; Bankston, J.R. Insights into the molecular mechanisms underlying the inhibition of acid-sensing ion channel 3 gating by stomatin. J. Gen. Physiol. 2020, 152, e201912471. [Google Scholar] [CrossRef]
- Martinez-Salgado, C.; Benckendorff, A.G.; Chiang, L.Y.; Wang, R.; Milenkovic, N.; Wetzel, C.; Hu, J.; Stucky, C.L.; Parra, M.G.; Mohandas, N.; et al. Stomatin and sensory neuron mechanotransduction. J. Neurophysiol. 2007, 98, 3802–3808. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulos, A.; Thorpe, S.D.; Cortes, E.; Lachowski, D.; Rice, A.J.; Mykuliak, V.V.; Róg, T.; Lee, D.A.; Hytönen, V.P.; Armando, E. Syndecan-4 tunes cell mechanics by activating the kindlin-integrin-RhoA pathway. Nat. Mater. 2020, 19, 669–678. [Google Scholar] [CrossRef]
- Wu, J.; Lewis, A.H.; Grandl, J. Touch, tension, and transduction—The function and regulation of piezo ion channels. Trends Biochem. Sci. 2017, 42, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Murthy, S.E.; Dubin, A.E.; Patapoutian, A. Piezos thrive under pressure: Mechanically activated ion channels in health and disease. Nat. Rev. Mol. Cell. Biol. 2017, 18, 771–783. [Google Scholar] [CrossRef]
- Bagriantsev, S.N.; Gracheva, E.O.; Gallagher, P.G. Piezo proteins: Regulators of mechanosensation and other cellular processes. J. Biol. Chem. 2014, 289, 31673–31681. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.H.; Ranade, S.; Weyer, A.D.; Dubin, A.E.; Baba, Y.; Qiu, Z.; Petrus, M.; Miyamoto, T.; Reddy, K.; Lumpkin, E.A.; et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature 2014, 509, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, P.A.; Brierley, S.M.; Young, R.L.; Blackshaw, L.A. Localization and comparative analysis of acid-sensing ion channel (ASIC1, 2, and 3) mRNA expression in mouse colonic sensory neurons within thoracolumbar dorsal root ganglia. J. Comp. Neurol. 2007, 500, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Brierley, S.M.; Martin, C.M.; Price, M.P.; Symonds, E.; Butler, R.; Wemmie, J.A.; Blackshaw, L.A. Different contributions of ASIC channels 1a, 2, and 3 in gastrointestinal mechanosensory function. Gut 2005, 54, 1408–1415. [Google Scholar] [CrossRef] [Green Version]
- Corrow, K.; Girard, B.M.; Vizzard, M.A. Expression and response of acid-sensing ion channels in urinary bladder to cyclophosphamide-induced cystitis. Am. J. Physiol. Renal Physiol. 2010, 298, F1130–F1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Brierley, S.M.; Martin, C.M.; Martinez-Salgado, C.; Wemmie, J.A.; Brennan, T.J.; Symonds, E.; Omari, T.; Lewin, G.R.; Welsh, M.J.; et al. The ion channel ASIC1 contributes to visceral but not cutaneous mechanoreceptor function. Gastroenterology 2004, 127, 1739–1747. [Google Scholar] [CrossRef] [PubMed]
- Calavia, M.G.; Montaño, J.A.; García-Suárez, O.; Feito, J.; Guervós, M.A.; Germanà, A.; Del Valle, M.; Pérez-Piñera, P.; Cobo, J.; Vega, J.A. Differential localization of Acid-sensing ion channels 1 and 2 in human cutaneus pacinian corpuscles. Cell. Mol. Neurobiol. 2010, 30, 841–848. [Google Scholar] [CrossRef]
- Walder, R.Y.; Rasmussen, L.A.; Rainier, J.D.; Light, A.R.; Wemmie, J.A.; Sluka, K.A. ASIC1 and ASIC3 play different roles in the development of hyperalgesia after inflammatory muscle injury. J. Pain 2010, 11, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Verkest, C.; Piquet, E.; Diochot, S.; Dauvois, M.; Lanteri-Minet, M.; Lingueglia, E.; Baron, A. Effects of systemic inhibitors of acid-sensing ion channels 1 (ASIC1) against acute and chronic mechanical allodynia in a rodent model of migraine. Br. J. Pharmacol. 2018, 175, 4154–4166. [Google Scholar] [CrossRef] [Green Version]
- Jernigan, N.L.; Herbert, L.M.; Walker, B.R.; Resta, T.C. Chronic hypoxia upregulates pulmonary arterial ASIC1: A novel mechanism of enhanced store-operated Ca2+ entry and receptor-dependent vasoconstriction. Am. J. Physiol. Cell Physiol. 2012, 302, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Huque, T.; Cowart, B.J.; Dankulich-Nagrudny, L.; Pribitkin, E.A.; Bayley, D.L.; Spielman, A.I.; Feldman, R.S.; Mackler, S.A.; Brand, J.G. Sour ageusia in two individuals implicates ion channels of the ASIC and PKD families in human sour taste perception at the anterior tongue. PLoS ONE 2009, 4, e7347. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.M.; Parameswaran, S.; Sahu, G.; Verma, R.S.; Bera, A.K. Proton-gated ion channels in mouse bone marrow stromal cells. Stem Cell Res. 2012, 9, 59–68. [Google Scholar] [CrossRef]
- Dong, X.; Ko, K.H.; Chow, J.; Tuo, B.; Barrett, K.E.; Dong, H. Expression of acid-sensing ion channels in intestinal epithelial cells and their role in the regulation of duodenal mucosal bicarbonate secretion. Acta Physiol. 2011, 201, 97–107. [Google Scholar] [CrossRef]
- Cabo, R.; Alonso, P.; Viña, E.; Vázquez, G.; Gago, A.; Feito, J.; Pérez-Moltó, F.J.; García-Suárez, O.; Vega, J.A. ASIC2 is present in human mechanosensory neurons of the dorsal root ganglia and in mechanoreceptors of the glabrous skin. Histochem. Cell Biol. 2015, 143, 267–276. [Google Scholar] [CrossRef]
- La, J.H.; Feng, B.; Kaji, K.; Schwartz, E.S.; Gebhart, G.F. Roles of isolectin B4-binding afferents in colorectal mechanical nociception. Pain 2016, 157, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Dusenkova, S.; Ru, F.; Surdenikova, L.; Nassenstein, C.; Hatok, J.; Dusenka, R.; Banovcin, P., Jr.; Kliment, J.; Tatar, M.; Kollarik, M. The expression profile of acid-sensing ion channel (ASIC) subunits ASIC1a, ASIC1b, ASIC2a, ASIC2b, and ASIC3 in the esophageal vagal afferent nerve subtypes. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G922–G930. [Google Scholar] [CrossRef]
- Hattori, T.; Chen, J.; Harding, A.M.; Price, M.P.; Lu, Y.; Abboud, F.M.; Benson, C.J. ASIC2a and ASIC3 heteromultimerize to form pH-sensitive channels in mouse cardiac dorsal root ganglia neurons. Circ. Res. 2009, 105, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abboud, F.M.; Benson, C.J. ASICs and cardiovascular homeostasis. Neuropharmacology 2015, 94, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Leng, T.; Jing, L.; Jiang, N.; Chen, D.; Hu, Y.; Xiong, Z.G.; Zha, X.M. Two di-leucine motifs regulate trafficking and function of mouse ASIC2a. Mol. Brain. 2016, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Moshourab, R.A.; Wetzel, C.; Martinez-Salgado, C.; Lewin, G.R. Stomatin-domain protein interactions with acid-sensing ion channels modulate nociceptor mechanosensitivity. J. Physiol. 2013, 591, 5555–5574. [Google Scholar] [CrossRef] [Green Version]
- Price, M.P.; McIlwrath, S.L.; Xie, J.; Cheng, C.; Qiao, J.; Tarr, D.E.; Sluka, K.A.; Brennan, T.J.; Lewin, G.R.; Welsh, M.J. The DRASIC cation channel contributes to the detection of cutaneous touch and acid stimuli in mice. Neuron 2001, 32, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Dulai, J.S.; Smith, E.S.J.; Rahman, T. Acid-sensing ion channel 3: An analgesic target. Channels 2021, 15, 94–127. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Chen, C.C. Roles of ASICs in nociception and proprioception. Adv. Exp. Med. Biol. 2018, 1099, 37–47. [Google Scholar] [PubMed]
- Wemmie, J.A.; Taugher, R.J.; Kreple, C.J. Acid-sensing ion channels in pain and disease. Nat. Rev. Neurosci. 2013, 14, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Sluka, K.A.; Gregory, N.S. The dichotomized role for acid sensing ion channels in musculoskeletal pain and inflammation. Neuropharmacology 2015, 94, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deval, E.; Lingueglia, E. Acid-sensing ion channels and nociception in the peripheral and central nervous systems. Neuropharmacology 2015, 94, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Cheng, Y.R.; Banks, R.W.; Min, M.Y.; Bewick, G.S.; Chen, C.C. Evidence for the involvement of ASIC3 in sensory mechanotransduction in proprioceptors. Nat. Commun. 2016, 7, 11460. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Gao, Z.; Li, J. Femoral artery occlusion increases expression of ASIC3 in dorsal root ganglion neurons. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1357–H1364. [Google Scholar] [CrossRef]
- Delaunay, A.; Gasull, X.; Salinas, M.; Noël, J.; Friend, V.; Lingueglia, E.; Deval, E. Human ASIC3 channel dynamically adapts its activity to sense the extracellular pH in both acidic and alkaline directions. Proc. Natl. Acad. Sci. USA 2012, 109, 13124–13129. [Google Scholar] [CrossRef] [Green Version]
- Kung, C.C.; Huang, Y.C.; Hung, T.Y.; Teng, C.Y.; Lee, T.Y.; Sun, W.H. Deletion of acid-sensing ion channel 3 relieves the late phase of neuropathic pain by preventing neuron degeneration and promoting neuron repair. Cells 2020, 9, 2355. [Google Scholar] [CrossRef]
- Sluka, K.A.; Price, M.P.; Breese, N.M.; Stucky, C.L.; Wemmie, J.A.; Welsh, M.J. Chronic hyperalgesia induced by repeated acid injections in muscle is abolished by the loss of ASIC3, but not ASIC1. Pain 2003, 106, 229–239. [Google Scholar] [CrossRef]
- Jalalvand, E.; Robertson, B.; Tostivint, H.; Löw, P.; Wallén, P.; Grillner, S. Cerebrospinal fluid-contacting neurons sense pH changes and motion in the hypothalamus. J. Neurosci. 2018, 38, 7713–7724. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Subtype of ASICs | Mechanosensitive Function |
|---|---|
| ASIC1 |
|
| ASIC2 | |
| ASIC3 |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, N.; Tribble, J.; Peterson, A.M.; Jiang, Q.; Wang, J.Q.; Chu, X.-P. Acid-Sensing Ion Channels and Mechanosensation. Int. J. Mol. Sci. 2021, 22, 4810. https://doi.org/10.3390/ijms22094810
Ruan N, Tribble J, Peterson AM, Jiang Q, Wang JQ, Chu X-P. Acid-Sensing Ion Channels and Mechanosensation. International Journal of Molecular Sciences. 2021; 22(9):4810. https://doi.org/10.3390/ijms22094810
Chicago/Turabian StyleRuan, Nina, Jacob Tribble, Andrew M. Peterson, Qian Jiang, John Q. Wang, and Xiang-Ping Chu. 2021. "Acid-Sensing Ion Channels and Mechanosensation" International Journal of Molecular Sciences 22, no. 9: 4810. https://doi.org/10.3390/ijms22094810
APA StyleRuan, N., Tribble, J., Peterson, A. M., Jiang, Q., Wang, J. Q., & Chu, X. -P. (2021). Acid-Sensing Ion Channels and Mechanosensation. International Journal of Molecular Sciences, 22(9), 4810. https://doi.org/10.3390/ijms22094810