
Potential of Silver Nanoparticles in Overcoming the Intrinsic Resistance of Pseudomonas aeruginosa to Secondary Metabolites from Carnivorous Plants

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
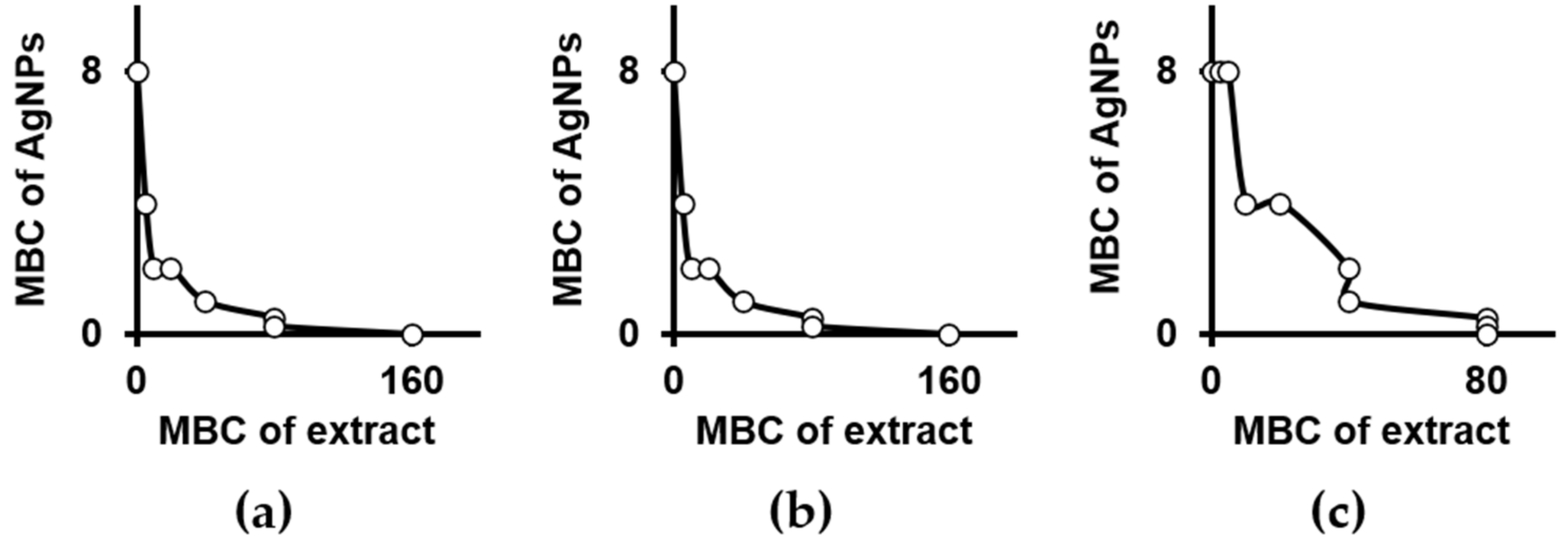
2.1. Antimicrobial Potential of Plant Extracts, Silver Nanoparticles and Their Combinations
2.2. Secondary Metabolites in Plant Extracts
2.3. Identification of Secondary Metabolites from Dionaea muscipula
2.4. Antimicrobial Potential of D. muscipula Constituents and Their Interaction with AgNPs
2.5. Activity of Selected Secondary Metabolites in Combination with AgNPs
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Bacteria, Antimicrobials and Pure Compounds
4.3. Silver Nanoparticles
4.4. Plant Tissue Extraction Method
4.5. Fractionation of D. muscipula Extract
4.6. HPLC-DAD-ESI/MS Analyses
4.7. Determination of Antimicrobial Potential
4.8. Determination of Combinatorial Effect of Antimicrobials
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evans, W.C.; Evans, D.; Trease, E.G. Trease & Evans’ Pharmacognosy, 15th ed.; WB Saunders: Philadelphia, PA, USA, 2002. [Google Scholar]
- Aminov, R.I. A Brief History of the Antibiotic Era: Lessons Learned and Challenges for the Future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrêa, R.C.; Heleno, S.A.; Alves, M.J.; Ferreira, I.C. Bacterial Resistance: Antibiotics of Last Generation used in Clinical Practice and the Arise of Natural Products as New Therapeutic Alternatives. Curr. Pharm. Des. 2020, 26, 815–837. [Google Scholar] [CrossRef]
- Hatcher, C.R.; Ryves, D.B.; Millett, J. The function of secondary metabolites in plant carnivory. Ann. Bot. 2019, 125, 399–411. [Google Scholar] [CrossRef]
- Egan, P.A.; van der Kooy, F. Phytochemistry of the carnivorous sundew genus Drosera (Droseraceae)—future perspectives and ethnopharmacological relevance. Chem. Biodivers. 2013, 10, 1774–1790. [Google Scholar] [CrossRef]
- Kovacik, J.; Klejdus, B.; Repcakova, K. Phenolic metabolites in carnivorous plants: Inter-specific comparison and physiological studies. Plant Physiol. Biochem. 2012, 52, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Šamaj, J.; Blehová, A.; Repčák, M.; Ovečka, M.; Bobák, M. Drosera Species (Sundew): In Vitro Culture and the Production of Plumbagin and Other Secondary Metabolites. In Biotechnology in Agriculture and Forestry 43. Medicinal and Aromatic Plants XI; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1999; pp. 105–135. [Google Scholar]
- Grevenstuk, T.; Gonçalves, S.; Domingos, T.; Quintas, C.; Van Der Hooft, J.J.; Vervoort, J.; Romano, A. Inhibitory activity of plumbagin produced by Drosera intermedia on food spoilage fungi. J. Sci. Food Agric. 2011, 92, 1638–1642. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Berchová-Bímová, K.; Petráš, J. Plumbagin, a Plant-Derived Compound, Exhibits Antifungal Combinatory Effect with Amphotericin B against Candida albicans Clinical Isolates and Anti-hepatitis C Virus Activity. Phytother. Res. 2016, 30, 1487–1492. [Google Scholar] [CrossRef]
- Lajubutu, B.A.; Pinney, R.J.; Roberts, M.F.; Odelola, H.A.; Oso, B.A. Antibacterial activity of diosquinone and plumbagin from the root of Diospyros mespiliformis (Hostch) (Ebenaceae). Phytother. Res. 1995, 9, 346–350. [Google Scholar] [CrossRef]
- Krolicka, A.; Szpitter, A.; Gilgenast, E.; Romanik, G.; Kaminski, M.; Lojkowska, E. Stimulation of antibacterial naphthoquinones and flavonoids accumulation in carnivorous plants by addition of elicitors. Enzyme Microb. Technol. 2008, 42, 216–221. [Google Scholar] [CrossRef]
- Ferreira, D.T.; Andrei, C.C.; Saridakis, H.O.; Faria, T.D.J.; Vinhato, E.; Carvalho, K.E.; Daniel, J.F.S.; Machado, S.L.; Saridakis, D.P.; Braz-Filho, R. Antimicrobial activity and chemical investigation of Brazilian Drosera. Memórias Inst. Oswaldo Cruz 2004, 99, 753–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krolicka, A.; Szpitter, A.; Maciag, M.; Biskup, E.; Gilgenast, E.; Romanik, G.; Kaminski, M.; Wegrzyn, G.; Lojkowska, E. Anti-bacterial and antioxidant activity of the secondary metabolites from in vitro cultures of the Alice sundew (Drosera aliciae). Biotechnol. Appl. Biochem. 2009, 53, 175–184. [Google Scholar] [CrossRef]
- Makowski, W.; Królicka, A.; Nowicka, A.; Zwyrtková, J.; Tokarz, B.; Pecinka, A.; Banasiuk, R.; Tokarz, K.M. Transformed tissue of Dionaea muscipula J. Ellis as a source of biologically active phenolic compounds with bactericidal properties. Appl. Microbiol. Biotechnol. 2021, 105, 1215–1226. [Google Scholar] [CrossRef]
- Veesenmeyer, J.L.; Hauser, A.R.; Lisboa, T.; Rello, J. Pseudomonas aeruginosa virulence and therapy: Evolving translational strategies*. Crit. Care Med. 2009, 37, 1777–1786. [Google Scholar] [CrossRef] [Green Version]
- Strateva, T.; Yordanov, D. Pseudomonas aeruginosa—a phenomenon of bacterial resistance. J. Med. Microbiol. 2009, 58, 1133–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuete, V.; Alibert-Franco, S.; Eyong, K.; Ngameni, B.; Folefoc, G.; Nguemeving, J.; Tangmouo, J.; Fotso, G.; Komguem, J.; Ouahouo, B.; et al. Antibacterial activity of some natural products against bacteria expressing a multidrug-resistant phenotype. Int. J. Antimicrob. Agents 2011, 37, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Krychowiak, M.; Grinholc, M.; Banasiuk, R.; Krauze-Baranowska, M.; Głód, D.; Kawiak, A.; Królicka, A. Combination of Silver Nanoparticles and Drosera binata Extract as a Possible Alternative for Antibiotic Treatment of Burn Wound Infections Caused by Resistant Staphylococcus aureus. PLoS ONE 2014, 9, e115727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atiyeh, B.S.; Costagliola, M.; Hayek, S.N.; Dibo, S.A. Effect of silver on burn wound infection control and healing: Review of the literature. Burns 2007, 33, 139–148. [Google Scholar] [CrossRef]
- Gravante, G.; Caruso, R.; Sorge, R.; Nicoli, F.; Gentile, P.; Cervelli, V. Nanocrystalline silver: A systematic review of random-ized trials conducted on burned patients and an evidence-based assessment of potential advantages over older silver formulations. Ann. Plast. Surg. 2009, 63, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Church, D.; Elsayed, S.; Reid, O.; Winston, B.; Lindsay, R. Burn Wound Infections. Clin. Microbiol. Rev. 2006, 19, 403–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, M.; Yadav, A.; Gade, A. Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 2009, 27, 76–83. [Google Scholar] [CrossRef]
- Feng, Q.L.; Wu, J.; Chen, G.Q.; Cui, F.Z.; Kim, T.N.; Kim, J.O. A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mater. Res. 2000, 52, 662–668. [Google Scholar] [CrossRef]
- Guzman, M.; Dille, J.; Godet, S. Synthesis and antibacterial activity of silver nanoparticles against gram-positive and gram-negative bacteria. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 37–45. [Google Scholar] [CrossRef]
- Braunberger, C.; Zehl, M.; Conrad, J.; Fischer, S.; Adhami, H.R.; Beifuss, U.; Krenn, L. LC-NMR, NMR, and LC-MS identifi-cation and LC-DAD quantification of flavonoids and ellagic acid derivatives in Drosera peltata. J. Chromatogr. B 2013, 932, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Braunberger, C.; Zehl, M.; Conrad, J.; Wawrosch, C.; Strohbach, J.; Beifuss, U.; Krenn, L. Flavonoids as chemotaxonomic markers in the genus Drosera. Phytochemistry 2015, 118, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Gaascht, F.; Dicato, M.; Diederich, M. Venus Flytrap (Dionaea muscipula Solander ex Ellis) Contains Powerful Compounds that Prevent and Cure Cancer. Front. Oncol. 2013, 3, 202. [Google Scholar] [CrossRef] [Green Version]
- Khallouki, F.; Haubner, R.; Hull, W.; Erben, G.; Spiegelhalder, B.; Bartsch, H.; Owen, R. Isolation, purification and identification of ellagic acid derivatives, catechins, and procyanidins from the root bark of Anisophyllea dichostyla R. Br. Food Chem. Toxicol. 2007, 45, 472–485. [Google Scholar] [CrossRef]
- Culham, A.; Gornall, R.J. The taxonomic significance of naphthoquinones in the Droseraceae. Biochem. Syst. Ecol. 1994, 22, 507–515. [Google Scholar] [CrossRef]
- Chakainresu, C. Synthesis and Structure-Activity Relationship Studies of 1,4-Naphthoquinone Derivatives as Potential An-Ti-trypanosomal Agents. Master’s Thesis, Rhodes University, Makhanda, South Africa, February 2014. [Google Scholar]
- Pei, J.; Hsu, C.-C.; Zhang, R.; Wang, Y.; Yu, K.; Huang, G. Unexpected Reduction of Iminoquinone and Quinone Derivatives in Positive Electrospray Ionization Mass Spectrometry and Possible Mechanism Exploration. J. Am. Soc. Mass Spectrom. 2017, 28, 2454–2461. [Google Scholar] [CrossRef] [PubMed]
- Higa, M.; Takashima, Y.; Yokaryo, H.; Harie, Y.; Suzuka, T.; Ogihara, K. Naphthoquinone derivatives from Diospyros maritima. Chem. Pharm. Bull. 2017, 65, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Markham, K.R. Techniques of Flavonoid Identification; Academic Press: New York, NY, USA, 1982. [Google Scholar]
- Krychowiak, M.; Kawiak, A.; Narajczyk, M.; Borowik, A.; Królicka, A. Silver Nanoparticles Combined With Naphthoquinones as an Effective Synergistic Strategy Against Staphylococcus aureus. Front. Pharmacol. 2018, 9, 816. [Google Scholar] [CrossRef] [Green Version]
- WHO. Antibacterial Agents in Clinical Development: An Analysis of the Antibacterial Clinical Development Pipeline; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- CDC. Antibiotic Resistance Threats in the United States; Department of Health and Human Services: Atlanta, GA, USA, 2019. [Google Scholar]
- French, G.L. Clinical impact and relevance of antibiotic resistance. Adv. Drug Deliv. Rev. 2005, 57, 1514–1527. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Chen, Y.; Ma, B.; He, J.; Tong, J.; Wang, Y. Drosera peltata Smith var. lunata (Buch.-Ham.) C.B. Clarke as a feasible source of plumbagin: phytochemical analysis and antifungal activity assay. World J. Microbiol. Biotechnol. 2014, 30, 737–745. [Google Scholar] [CrossRef]
- Makowski, W.; Tokarz, K.M.; Tokarz, B.; Banasiuk, R.; Witek, K.; Królicka, A. Elicitation-Based Method for Increasing the Production of Antioxidant and Bactericidal Phenolic Compounds in Dionaea muscipula J. Ellis Tissue. Molecules 2020, 25, 1794. [Google Scholar] [CrossRef]
- Kawiak, A.; Krolicka, A.; Lojkowska, E. In vitro cultures of Drosera aliciae as a source of a cytotoxic naphthoquinone: ramen-taceone. Biotechnol. Lett. 2011, 33, 2309–2316. [Google Scholar] [CrossRef]
- Krolicka, A.; Szpitter, A.; Maciag, M.; Biskup, E.; Gilgenast, E.; Romanik, G.; Kaminski, M.; Wegrzyn, G.; Lojkowska, E. An-tibacterial activity of the secondary metabolites from in vitro cultures of Drosera aliciae. Planta Med. 2008, 74, 961. [Google Scholar] [CrossRef]
- Fischbach, M.A. Combination therapies for combating antimicrobial resistance. Curr. Opin. Microbiol. 2011, 14, 519–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acar, J.F. Antibiotic synergy and antagonism. Med. Clin. N. Am. 2000, 84, 1391–1406. [Google Scholar] [CrossRef]
- Kastoris, A.C.; Rafailidis, P.I.; Vouloumanou, E.K.; Gkegkes, I.D.; Falagas, M.E. Synergy of fosfomycin with other antibiotics for Gram-positive and Gram-negative bacteria. Eur. J. Clin. Pharmacol. 2010, 66, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Shao, X.; Di, X.; Cui, J.; Wang, R.; Liu, Y. In vitro synergy of polymyxins with other antibiotics for Acinetobacter bau-mannii: A systematic review and meta-analysis. Int. J. Antimicrob. Agents 2015, 45, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Wright, G.D. Drug combinations: a strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Genet. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Krychowiak-Masnicka, M. Combining Silver Nanoparticles and Secondary Metabolites from Tissues of Carnivorous Plants as A Strategy to Combat Human Pathogenic Bacteria. Ph.D. Thesis, University of Gdansk, Gdansk, Poland, 11 January 2019. [Google Scholar]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic Basis of Antimicrobial Actions of Silver Nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klotz, L.-O.; Hou, X.; Jacob, C. 1,4-Naphthoquinones: From Oxidative Damage to Cellular and Inter-Cellular Signaling. Molecules 2014, 19, 14902–14918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Tokarz, K.; Makowski, W.; Banasiuk, R.; Krolicka, A.; Piwowarczyk, B. Response of Dionaea muscipula J. Ellis to light stress in in vitro: Physiological study. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Barenbaum, M.C. A method for testing for synergy with any number of agents. J. Infect. Dis. 1978, 137, 122–130. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Extracts | MBC | Unit |
|---|---|---|
| Dionaea muscipula | 160 | |
| Drosera binata | 160 | mg FW mL−1 |
| Drosera gigantea | 80 |
| Plant Species | FBCI | Plumbagin (µg g−1 FW) |
|---|---|---|
| D. muscipula | 0.31 | 3450 ± 245 |
| D. binata | 0.31 | 2035 ± 145 |
| D. gigantea | 0.625 | 145 ± 25.5 |
| Peak Number | tR | UV (λmax nm) | MS m/z | Compound [Reference] | |
|---|---|---|---|---|---|
| [M+H]+/[M-H]−/Ag+ | [M+2H]+/[M+H]− | ||||
| 1 | 9.4 | 253, 263sh, 302sh, 357 | 564+/-/- | - | unidentified flavonoid, probably quercetin glycoside |
| 2 | 10.0 | 265, 298sh, 357 | 548+/-/- | - | unidentified flavonoid, probably kaempferol glycoside |
| 3 | 10.8 | 247, 301sh, 350sh, 363 | 608+/-/- | - | unidentified compound |
| 4 | 11.31 | 252, 298sh, 366 | 303+/301− | - | ellagic acid [25,27] |
| 5 | 12.00 | 240, 290sh, 367 | 331+/329−/- | - | dimethylellagic acid isomer |
| 6 | 12.15 | 253, 266sh, 300sh, 355 | 465+/-/303+ | - | hyperoside-quercetin 3-O-galactoside [5,26,27] |
| 7 | 13.12 | 260, 289sh, 350sh | 631− | unknown compound | |
| 8 | 13.25 | 258, 266sh, 293sh, 357 | 617+/615− | - | quercetin-3-(6′′-O-galloyl)-glucoside/galactoside [26,27] |
| 9 | 15.51 | 265, 299sh, 351 | 601+/599− | - | kaempferol -3-(6′′-O-galloyl)-glucoside [25,26] |
| 10 | 16.90 | 249, 300sh, 370 | 317+/315− | - | 3-O-methylellagic acid [27] |
| 11 | 22.75 | 245, 289sh, 374 | 331+/329− | - | 3,3′-di-O-methylellagic acid [25,27] |
| 12 | 24.32 | 267, 346 | 192+/- | dihydroplumbagin [32] | |
| 13 | 31.38 | 266, 418 | - | 190+/189− | plumbagin [5,27] |
| 14 | 34.17 | 271, 412 | -/223− | 3-chloroplumbagin [5,27] | |
| 15 | 37.50 | 276, 414 | - | 376+/- | 8,8′-biplumbagin [27] |
| 16 | 40.90 | 280, 412 | 476+ | unknown naphthoquinone | |
| Compound | Presence of Compound in Fraction of D. muscipula Extract | ||||
|---|---|---|---|---|---|
| Peak Number | Name | 20% MeOH | 40% MeOH | 60% MeOH | 80% MeOH |
| 1 | unidentified flavonoid (probably quercetin glycoside) | + | |||
| 2 | unidentified flavonoid (probably kaempferol glycoside) | + | |||
| 3 | unidentified compound | ++ | + | ||
| 4 | ellagic acid | + | +++ | ++ | + |
| 5 | dimethylellagic acid isomer | ++ | + | ||
| 6 | hyperoside (quercetin 3-O-galactoside) | + | + | ||
| 7 | unknown compound | + | + | ||
| 8 | quercetin -3-(6″-O-galloyl)-glucoside/galactoside | + | + | ||
| 9 | kaempferol -3-(6″-O-galloyl)-glucoside | + | |||
| 10 | 3-O-methylellagic acid | ++ | +++ | + | |
| 11 | 3,3′-di-O-methylellagic acid | + | + | + | |
| 12 | 2,3-dihydroplumbagin | + | |||
| 13 | plumbagin | + | + | +++ | |
| 14 | 3-chloroplumbagin | + | |||
| 15 | 8,8′-biplumbagin | + | |||
| 16 | unknown naphthoquinone | + | |||
| Fraction | Prevalent Secondary Metabolites * | FBC (g FW mL−1) | Plumbagin (µg g−1 FW) |
|---|---|---|---|
| 20% methanol | phenolic acids | >4.48 | 0 |
| 40% methanol | flavonoids, flavonoid glycosides, phenolic acids | 2.24 | 6.14 ± 0.06 |
| 60% methanol | flavonoid glycosides, phenolic acids | 2.24 | 5.38 ± 0.02 |
| 80% methanol | naphthoquinones, phenolic acids | 0.14 | 1052 ± 12 |
| Secondary Metabolite | FBC (µg mL−1) | FBC Index |
|---|---|---|
| plumbagin | 64 | ≤0.31 |
| 3-chloroplumbagin | 16 | ≤0.128 |
| quercetin | >512 | ≥1.015 |
| kaempferol | >512 | ≥2.015 |
| ellagic acid | >512 | ≥1.015 |
| 3,3′-di-O-methylellagic acid | >512 | ≥1.015 |
| hyperoside | >512 | ≥1.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krychowiak-Maśnicka, M.; Krauze-Baranowska, M.; Godlewska, S.; Kaczyński, Z.; Bielicka-Giełdoń, A.; Grzegorczyk, N.; Narajczyk, M.; Frackowiak, J.E.; Krolicka, A. Potential of Silver Nanoparticles in Overcoming the Intrinsic Resistance of Pseudomonas aeruginosa to Secondary Metabolites from Carnivorous Plants. Int. J. Mol. Sci. 2021, 22, 4849. https://doi.org/10.3390/ijms22094849
Krychowiak-Maśnicka M, Krauze-Baranowska M, Godlewska S, Kaczyński Z, Bielicka-Giełdoń A, Grzegorczyk N, Narajczyk M, Frackowiak JE, Krolicka A. Potential of Silver Nanoparticles in Overcoming the Intrinsic Resistance of Pseudomonas aeruginosa to Secondary Metabolites from Carnivorous Plants. International Journal of Molecular Sciences. 2021; 22(9):4849. https://doi.org/10.3390/ijms22094849
Chicago/Turabian StyleKrychowiak-Maśnicka, Marta, Mirosława Krauze-Baranowska, Sylwia Godlewska, Zbigniew Kaczyński, Aleksandra Bielicka-Giełdoń, Natalia Grzegorczyk, Magdalena Narajczyk, Joanna E. Frackowiak, and Aleksandra Krolicka. 2021. "Potential of Silver Nanoparticles in Overcoming the Intrinsic Resistance of Pseudomonas aeruginosa to Secondary Metabolites from Carnivorous Plants" International Journal of Molecular Sciences 22, no. 9: 4849. https://doi.org/10.3390/ijms22094849
APA StyleKrychowiak-Maśnicka, M., Krauze-Baranowska, M., Godlewska, S., Kaczyński, Z., Bielicka-Giełdoń, A., Grzegorczyk, N., Narajczyk, M., Frackowiak, J. E., & Krolicka, A. (2021). Potential of Silver Nanoparticles in Overcoming the Intrinsic Resistance of Pseudomonas aeruginosa to Secondary Metabolites from Carnivorous Plants. International Journal of Molecular Sciences, 22(9), 4849. https://doi.org/10.3390/ijms22094849