
Stretch-Induced Tenomodulin Expression Promotes Tenocyte Migration via F-Actin and Chromatin Remodeling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
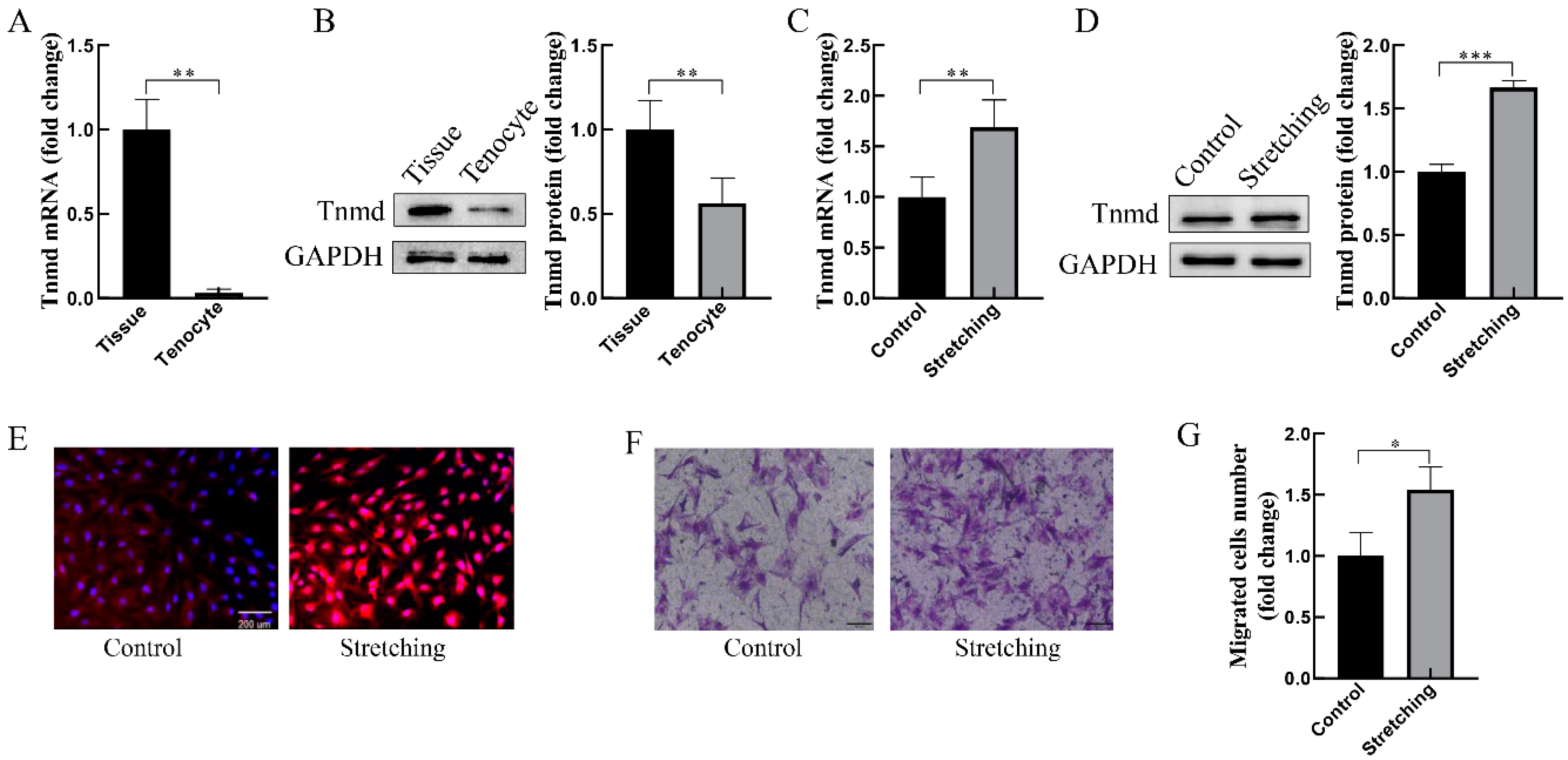
2.1. Mechanical Stretching Induces Tnmd Expression and Cell Migration in Cultured Tenocytes
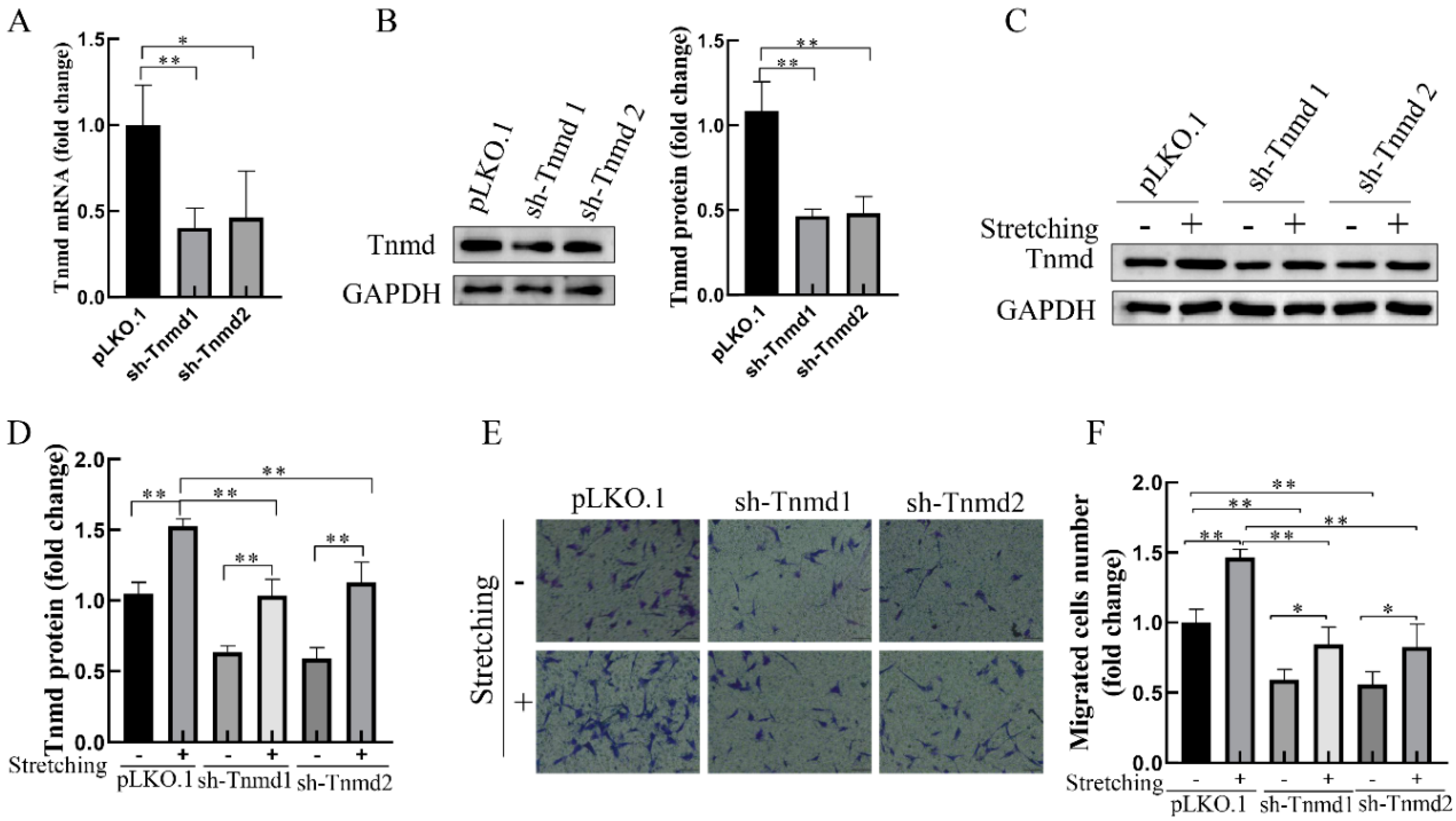
2.2. Stretching-Increased Tnmd Regulates the Migration of Tenocytes
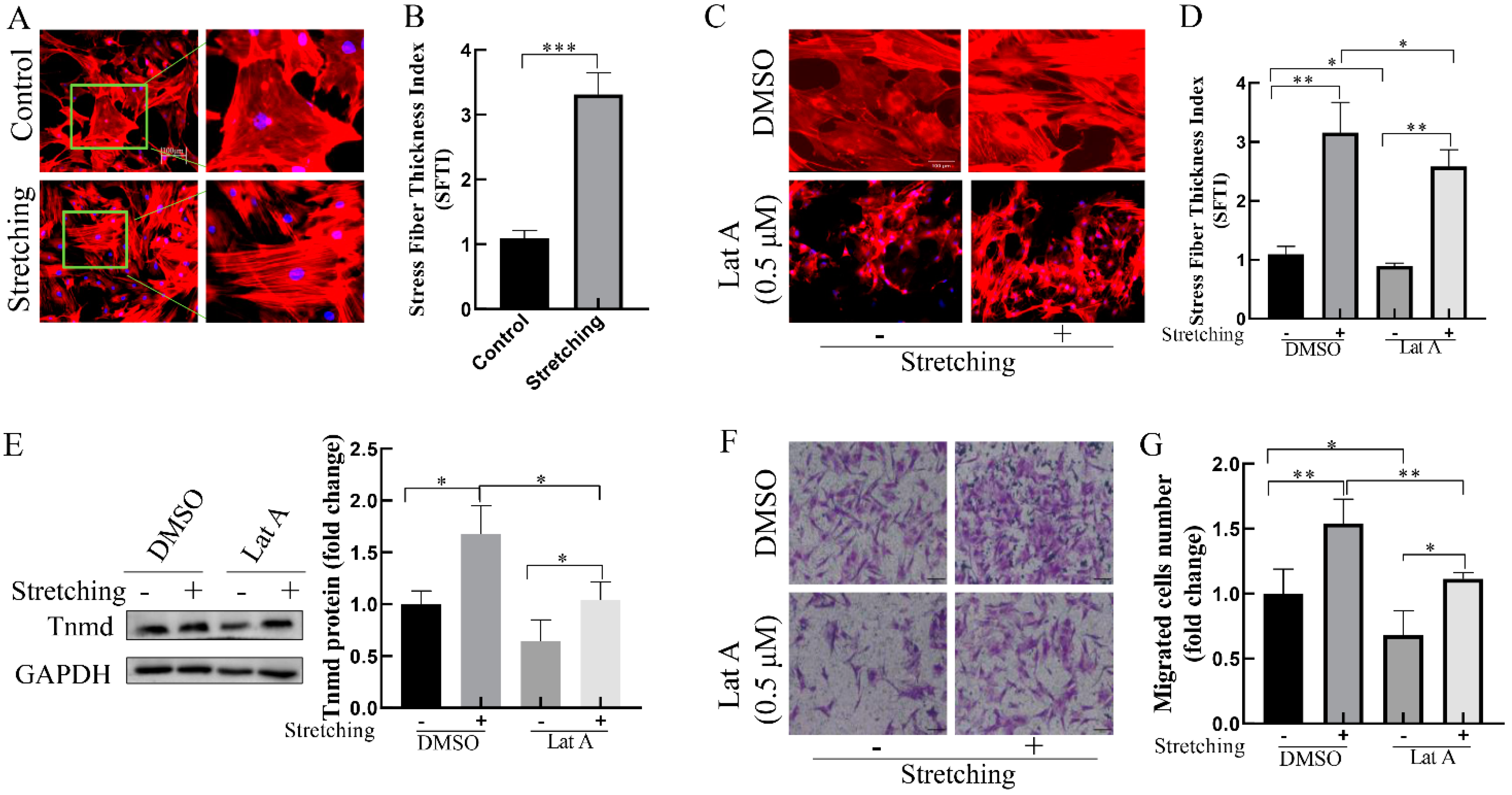
2.3. Stretch-Reinforced Actin Stress Fibers Increase Tnmd Expression and Tenocyte Migration
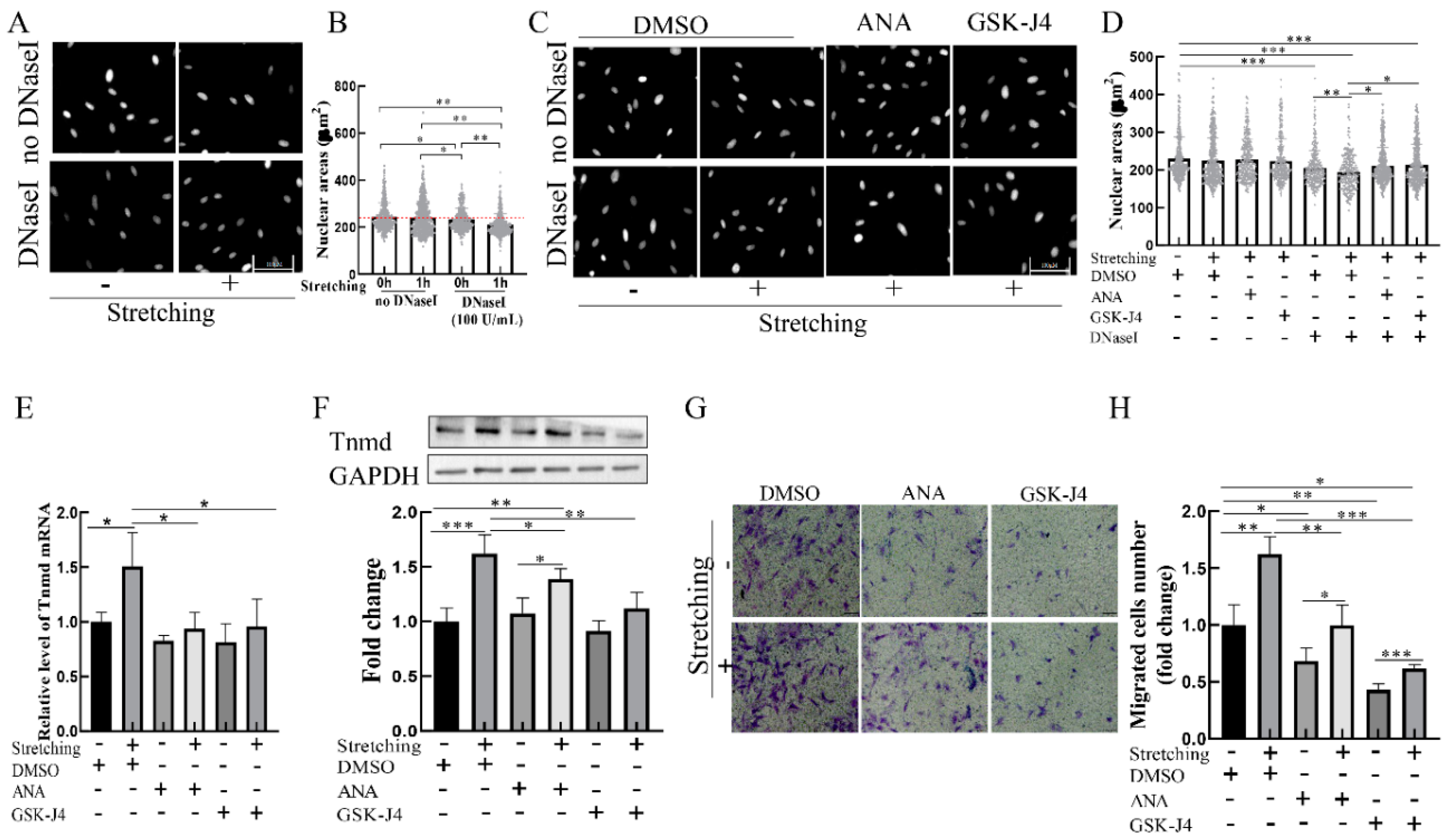
2.4. Stretch-Induced Chromatin Decondensation Increases Tnmd Expression and Tenocyte Migration
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Total RNA Extraction and Quantitative RT-PCR
4.3. Western Blotting
4.4. Uniaxial Cyclic Stretch
4.5. Transwell Migration Assay
4.6. Immunofluorescence
4.7. Stress Fiber Thickening Analysis
4.8. Construction of Lentiviral Vectors and Lentiviral Transduction
4.9. DNase I Sensitivity Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andarawis-Puri, N.; Flatow, E.L.; Soslowsky, L.J. Tendon basic science: Development, repair, regeneration, and healing. J. Orthop. Res. 2015, 33, 780–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayton, P. Anatomic, Vascular, and Mechanical Overview of the Achilles Tendon. Clin. Podiatr. Med. Surg. 2017, 34, 107–113. [Google Scholar] [CrossRef]
- Eliasson, P.; Andersson, T.; Aspenberg, P. Rat Achilles tendon healing: Mechanical loading and gene expression. J. Appl. Physiol. (1985) 2009, 107, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Galatz, L.M.; Gerstenfeld, L.; Heber-Katz, E.; Rodeo, S.A. Tendon regeneration and scar formation: The concept of scarless healing. J. Orthop. Res. 2015, 33, 823–831. [Google Scholar] [CrossRef]
- Alberton, P.; Dex, S.; Popov, C.; Shukunami, C.; Schieker, M.; Docheva, D. Loss of tenomodulin results in reduced self-renewal and augmented senescence of tendon stem/progenitor cells. Stem Cells Dev. 2015, 24, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Hiraki, Y.; Shukunami, C. Angiogenesis inhibitors localized in hypovascular mesenchymal tissues: Chondromodulin-I and tenomodulin. Connect Tissue Res. 2005, 46, 3–11. [Google Scholar] [CrossRef]
- Lin, D.; Alberton, P.; Caceres, M.D.; Volkmer, E.; Schieker, M.; Docheva, D. Tenomodulin is essential for prevention of adipocyte accumulation and fibrovascular scar formation during early tendon healing. Cell Death Dis. 2017, 8, e3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Caceres, M.D.; Yan, Z.; Schieker, M.; Nerlich, M.; Docheva, D. Tenomodulin regulates matrix remodeling of mouse tendon stem/progenitor cells in an ex vivo collagen I gel model. Biochem. Biophys. Res. Commun. 2019, 512, 691–697. [Google Scholar] [CrossRef]
- Hou, Y.; Ni, M.; Lin, S.; Sun, Y.; Lin, W.; Liu, Y.; Wang, H.; He, W.; Li, G.; Xu, L. Tenomodulin highly expressing MSCs as a better cell source for tendon injury healing. Oncotarget 2017, 8, 77424–77435. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.H. The Effects of Mechanical Loading on Tendons—an in vivo and in vitro model study. PLoS ONE 2013, 8, e71740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havis, E.; Bonnin, M.A.; Olivera-Martinez, I.; Nazaret, N.; Ruggiu, M.; Weibel, J.; Durand, C.; Guerquin, M.J.; Bonod-Bidaud, C.; Ruggiero, F.; et al. Transcriptomic analysis of mouse limb tendon cells during development. Development (Camb. Engl.) 2014, 141, 3683–3696. [Google Scholar] [CrossRef] [Green Version]
- Olson, E.N.; Nordheim, A. Linking actin dynamics and gene transcription to drive cellular motile functions. Nat. Rev. Mol. Cell Biol. 2010, 11, 353–365. [Google Scholar] [CrossRef] [Green Version]
- Schiele, N.R.; von Flotow, F.; Tochka, Z.L.; Hockaday, L.A.; Marturano, J.E.; Thibodeau, J.J.; Kuo, C.K. Actin cytoskeleton contributes to the elastic modulus of embryonic tendon during early development. J. Orthop. Res. 2015, 33, 874–881. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, L.; Jensen, C.C.; Yoshigi, M.; Beckerle, M. Mechanical signals activate p38 MAPK pathway-dependent reinforcement of actin via mechanosensitive HspB1. Mol. Biol. Cell 2017, 28, 2661–2675. [Google Scholar] [CrossRef] [PubMed]
- Tajik, A.; Zhang, Y.; Wei, F.; Sun, J.; Jia, Q.; Zhou, W.; Singh, R.; Khanna, N.; Belmont, A.S.; Wang, N. Transcription upregulation via force-induced direct stretching of chromatin. Nat. Mater. 2016, 15, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Gerlitz, G.; Bustin, M. Efficient cell migration requires global chromatin condensation. J. Cell Sci. 2010, 123, 2207–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawadkar, P.; Player, D.; Bozec, L.; Mudera, V. The mechanobiology of tendon fibroblasts under static and uniaxial cyclic load in a 3D tissue engineered model mimicking native ECM. J. Tissue Eng. Regen. Med. 2020, 14, 135–146. [Google Scholar] [CrossRef]
- Wall, M.E.; Banes, A.J. Early responses to mechanical load in tendon: Role for calcium signaling, gap junctions and intercellular communication. J. Musculoskelet. Neuronal. Interact. 2005, 5, 70–84. [Google Scholar] [PubMed]
- Wall, M.E.; Dyment, N.A.; Bodle, J.; Volmer, J.; Loboa, E.; Cederlund, A.; Fox, A.M.; Banes, A.J. Cell Signaling in Tenocytes: Response to Load and Ligands in Health and Disease. Adv. Exp. Med. Biol. 2016, 920, 79–95. [Google Scholar] [CrossRef]
- Docheva, D.; Hunziker, E.B.; Fassler, R.; Brandau, O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Dex, S.; Lin, D.; Shukunami, C.; Docheva, D. Tenogenic modulating insider factor: Systematic assessment on the functions of tenomodulin gene. Gene 2016, 587, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Dex, S.; Alberton, P.; Willkomm, L.; Sollradl, T.; Bago, S.; Milz, S.; Shakibaei, M.; Ignatius, A.; Bloch, W.; Clausen-Schaumann, H.; et al. Tenomodulin is Required for Tendon Endurance Running and Collagen I Fibril Adaptation to Mechanical Load. EBioMedicine 2017, 20, 240–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzocca, A.D.; Chowaniec, D.; McCarthy, M.B.; Beitzel, K.; Cote, M.P.; McKinnon, W.; Arciero, R. In vitro changes in human tenocyte cultures obtained from proximal biceps tendon: Multiple passages result in changes in routine cell markers. Knee Surg. Sports Traumatol. Arthrosc. Off. J. ESSKA 2012, 20, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, T.; Shukunami, C.; Okamoto, N.; Taniwaki, T.; Oka, K.; Sakamoto, H.; Ide, J.; Mizuta, H.; Hiraki, Y. FGF-2 Stimulates the Growth of Tenogenic Progenitor Cells to Facilitate the Generation of Tenomodulin-Positive Tenocytes in a Rat Rotator Cuff Healing Model. Am. J. Sports Med. 2015, 43, 2411–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelinsky, S.A.; Li, L.; Ellis, D.; Archambault, J.; Li, J.; St Andre, M.; Morris, C.; Seeherman, H. Treatment with rhBMP12 or rhBMP13 increase the rate and the quality of rat Achilles tendon repair. J. Orthop. Res. 2011, 29, 1604–1612. [Google Scholar] [CrossRef] [PubMed]
- Gouget, C.L.; Hwang, Y.; Barakat, A.I. Model of Cellular Mechanotransduction via Actin Stress Fibers. Biomech. Model Mechanobiol. 2016, 15, 331–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Li, J.; Wang, B.; Zhang, W.J.; Zhou, G.; Cao, Y.; Liu, W. The regulation of phenotype of cultured tenocytes by microgrooved surface structure. Biomaterials 2010, 31, 6952–6958. [Google Scholar] [CrossRef] [PubMed]
- Stephens, A.D. Chromatin rigidity provides mechanical and genome protection. Mutat. Res. 2020, 821, 111712. [Google Scholar] [CrossRef]
- Vankova Hausnerova, V.; Lanctot, C. Chromatin decondensation is accompanied by a transient increase in transcriptional output. Biol. Cell 2017, 109, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.M.; Zaganjor, E.; Cobb, M.H. Chromatin-tethered MAPKs. Curr. Opin. Cell Biol. 2013, 25, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Liang, Q.; Zhuang, W.; Zhou, J.; Zhang, B.; Xu, P.; Ju, Y.; Morita, Y.; Luo, Q.; Song, G. Tenocyte proliferation and migration promoted by rat bone marrow mesenchymal stem cell-derived conditioned medium. Biotechnol. Lett. 2018, 40, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Luo, Q.; Chen, Z.; Shi, Y.; Ju, Y.; Yang, L.; Song, G. Increased nuclear stiffness via FAK-ERK1/2 signaling is necessary for synthetic mechano-growth factor E peptide-induced tenocyte migration. Sci. Rep. 2016, 6, 18809. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, P.; Deng, B.; Zhang, B.; Luo, Q.; Song, G. Stretch-Induced Tenomodulin Expression Promotes Tenocyte Migration via F-Actin and Chromatin Remodeling. Int. J. Mol. Sci. 2021, 22, 4928. https://doi.org/10.3390/ijms22094928
Xu P, Deng B, Zhang B, Luo Q, Song G. Stretch-Induced Tenomodulin Expression Promotes Tenocyte Migration via F-Actin and Chromatin Remodeling. International Journal of Molecular Sciences. 2021; 22(9):4928. https://doi.org/10.3390/ijms22094928
Chicago/Turabian StyleXu, Pu, Bin Deng, Bingyu Zhang, Qing Luo, and Guanbin Song. 2021. "Stretch-Induced Tenomodulin Expression Promotes Tenocyte Migration via F-Actin and Chromatin Remodeling" International Journal of Molecular Sciences 22, no. 9: 4928. https://doi.org/10.3390/ijms22094928
APA StyleXu, P., Deng, B., Zhang, B., Luo, Q., & Song, G. (2021). Stretch-Induced Tenomodulin Expression Promotes Tenocyte Migration via F-Actin and Chromatin Remodeling. International Journal of Molecular Sciences, 22(9), 4928. https://doi.org/10.3390/ijms22094928