
Photoperiod Following Inoculation of Arabidopsis with Pyricularia oryzae (syn. Magnaporthe oryzae) Influences on the Plant–Pathogen Interaction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
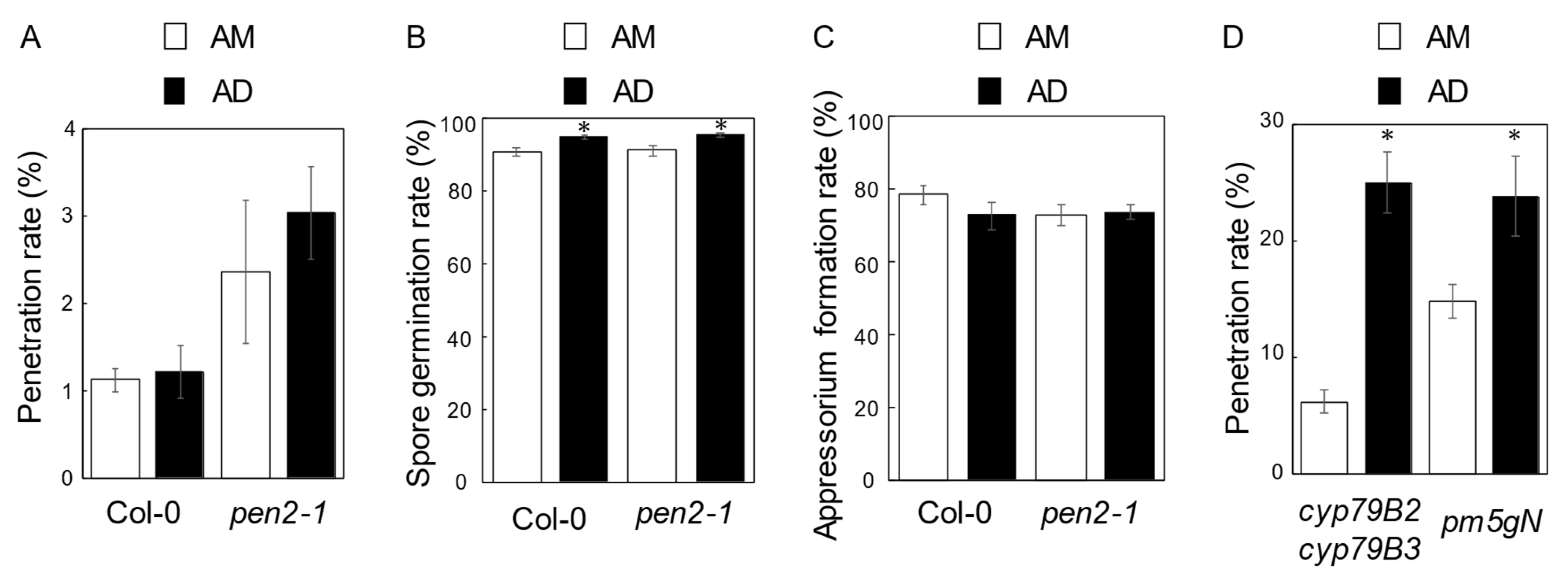
2. Results
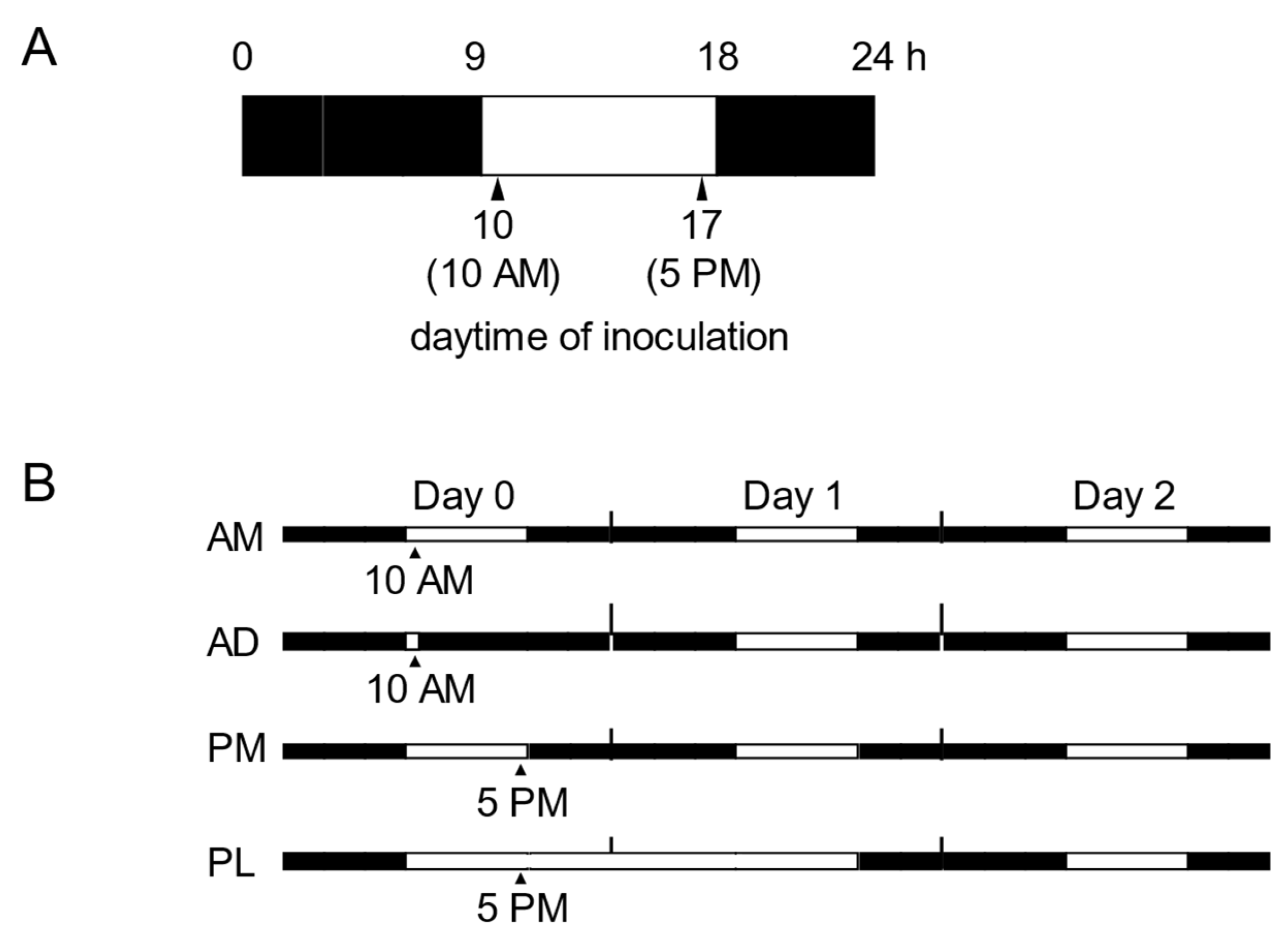
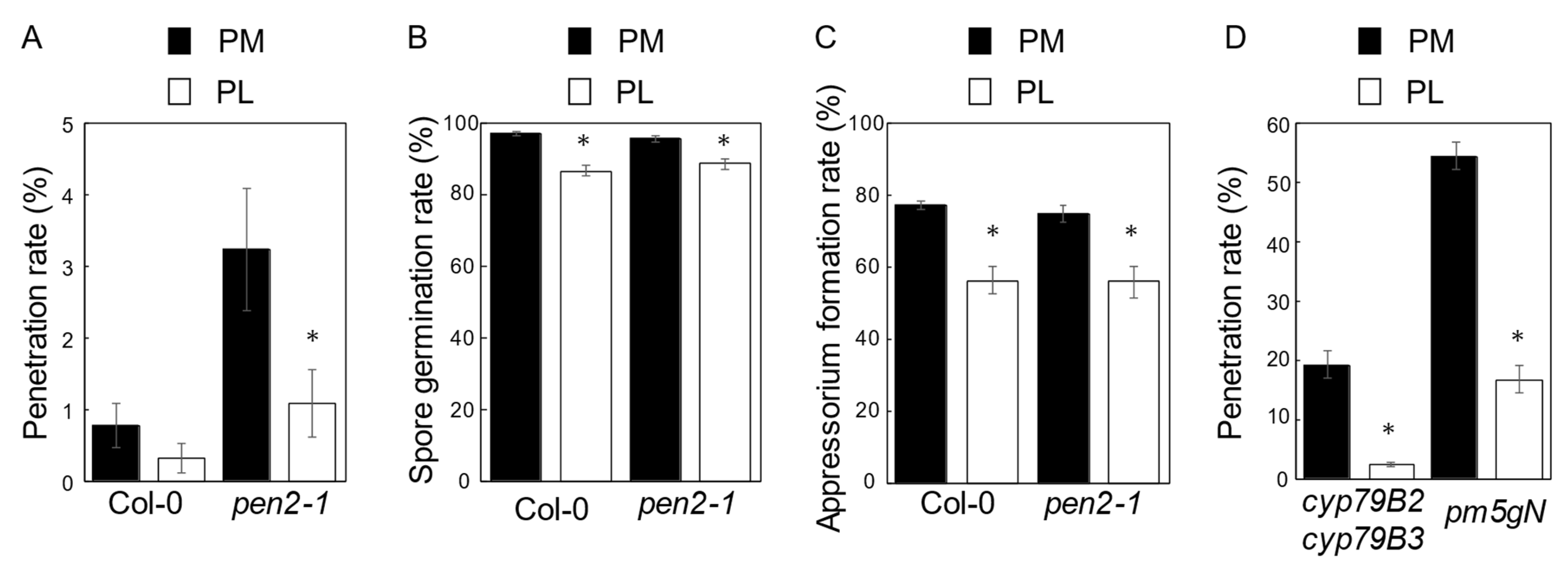
2.1. The Light Period Following Evening Inoculation Enhances NHR to P. oryzae in Arabidopsis
2.2. The Dark Period Following the Morning Inoculation Suppresses NHR to P. oryzae in Arabidopsis
2.3. Genes Controlling Photoperiod-Dependent NHR to P. oryzae in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Pathogen Inoculation and Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, M.R.; Paul, N.D. Seduced by the dark side: Integrating molecular and ecological perspectives on the influence of light on plant defence against pests and pathogens. New Phytol. 2006, 170, 677–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roden, L.C.; Ingle, R.A. Lights, rhythms, infection: The role of light and the circadian clock in determining the outcome of plant-pathogen interactions. Plant Cell 2009, 21, 2546–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballaré, C.L. Light Regulation of Plant Defense. Annu. Rev. Plant Biol. 2014, 65, 335–363. [Google Scholar] [CrossRef]
- Mawphlang, O.I.L.; Kharshiing, E.V. Photoreceptor Mediated Plant. Growth Responses: Implications for Photoreceptor Engineering toward Improved Performance in Crops. Front Plant Sci. 2017, 8, 1181. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Reimers, P.; Leach, J. Effect of light on incompatible interactions between Xanthomonas oryzae pv. oryzae and rice. Physiol. Mol. Plant. Pathol. 1993, 42, 413–425. [Google Scholar] [CrossRef]
- Genoud, T.; Buchala, A.J.; Chua, N.H.; Metraux, J.P. Phytochrome signaling modulates the SA-perceptive pathway in Arabidopsis. Plant J. 2002, 31, 87–95. [Google Scholar] [CrossRef]
- Zeier, J.; Pink, B.; Mueller, M.J.; Berger, S. Light conditions influence specific defence responses in incompatible plant-pathogen inter-actions: Uncoupling systemic resistance from salicylic acid and PR-1 accumulation. Planta 2004, 219, 673–683. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.; Navarre, R.; Kachroo, A.; Raina, S.; Raina, R.; Klessig, D.; Kachroo, P. Light dependent host defense sig-naling against Turnip Crinkle Virus in Arabidopsis. Phytopathology 2006, 96, S20. [Google Scholar]
- Littlejohn, G.R.; Breen, S.; Smirnoff, N.; Grant, M. Chloroplast immunity illuminated. New Phytol. 2021, 229, 3088–3107. [Google Scholar] [CrossRef]
- Joseph, M.E.; Hering, T.F. Effects of environment on spore germination and infection by broad bean rust (Uromyces viciae-fabae). J. Agric. Sci. 1997, 128, 73–78. [Google Scholar] [CrossRef]
- Tapsoba, H.; Wilson, J.P. Effects of temperature and light on germination of urediniospores of the pearl millet rust pathogen, Puccinia substriata var. indica. Plant Dis. 1997, 81, 1049–1052. [Google Scholar] [CrossRef] [Green Version]
- Mueller, D.S.; Buck, J.W. Effects of light, temperature, and leaf wetness duration on daylily rust. Plant Dis. 2003, 87, 442–445. [Google Scholar] [CrossRef] [Green Version]
- Beyer, M.; Roding, S.; Ludewig, A.; Verreet, J.A. Germination and survival of Fusarium graminearum macroconidia as affected by environmental factors. Phytopathology 2004, 152, 92–97. [Google Scholar] [CrossRef]
- Buck, J.W.; Dong, W.; Mueller, D.S. Effect of light exposure on in vitro germination and germ tube growth of eight species of rust fungi. Mycologia 2010, 102, 1134–1140. [Google Scholar] [CrossRef]
- Ayliffe, M.; Sorensen, C.K. Plant. nonhost resistance: Paradigms and new environments. Curr. Opin. Plant Biol. 2019, 50, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Panstruga, R.; Moscou, M.J. What is the Molecular Basis of Nonhost Resistance. Mol. Plant Microbe Interact. 2020, 33, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Collins, N.C.; Thordal-Christensen, H.; Lipka, V.; Bau, S.; Kombrink, E.; Qiu, J.L.; Huckelhoven, R.; Stein, M.; Freialdenhoven, A.; Somerville, S.C.; et al. SNARE-protein-mediated disease resistance at the plant cell wall. Nature 2003, 425, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Lipka, V.; Dittgen, J.; Bednarek, P.; Bhat, R.; Wiermer, M.; Stein, M.; Landtag, J.; Brandt, W.; Rosahl, S.; Scheel, D.; et al. Pre- and postinvasion defenses both contribute to nonhost resistance in Arabidopsis. Science 2005, 310, 1180–1183. [Google Scholar] [CrossRef] [Green Version]
- Kobae, Y.; Sekino, T.; Yoshioka, H.; Nakagawa, T.; Martinoia, E.; Maeshima, M. Loss of AtPDR8, a plasma membrane ABC trans-porter of Arabidopsis thaliana, causes hypersensitive cell death upon pathogen infection. Plant Cell Physiol. 2006, 47, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, M.; Dittgen, J.; Sanchez-Rodriguez, C.; Hou, B.H.; Molina, A.; Schulze-Lefert, P.; Lipka, V.; Somerville, S. Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration. Plant Cell 2006, 18, 731–746. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, P.; Pislewska-Bednarek, M.; Svatos, A.; Schneider, B.; Doubsky, J.; Mansurova, M.; Humphry, M.; Consonni, C.; Pan-struga, R.; Sanchez-Vallet, A.; et al. A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 2009, 323, 101–106. [Google Scholar] [CrossRef]
- Zhou, N.; Tootle, T.L.; Glazebrook, J. Arabidopsis PAD3, a gene required for camalexin biosynthesis, encodes a putative cytochrome P450 monooxygenase. Plant Cell 1999, 11, 2419–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.X.; Xu, J.; Wang, X.Y.; He, X.M.; Wang, Y.X.Y.; Zhou, J.G.; Zhang, S.Q.; Meng, X.Z. The Arabidopsis Pleiotropic Drug Re-sistance Transporters PEN3 and PDR12 Mediate Camalexin Secretion for Resistance to Botrytis cinerea. Plant Cell 2019, 31, 2206–2222. [Google Scholar] [CrossRef] [PubMed]
- Pastorczyk, M.; Bednarek, P. The Function of Glucosinolates and Related Metabolites in Plant. Innate Immunity. Glucosinolates 2016, 80, 171–198. [Google Scholar]
- Zhao, Y.D.; Hull, A.K.; Gupta, N.R.; Goss, K.A.; Alonso, J.; Ecker, J.R.; Normanly, J.; Chory, J.; Celenza, J.L. Trp-dependent auxin biosynthesis in Arabidopsis: Involvement of cytochrome P450s CYP79B2 and CYP79B3. Genes. Dev. 2002, 16, 3100–3112. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vallet, A.; Ramos, B.; Bednarek, P.; Lopez, G.; Pislewska-Bednarek, M.; Schulze-Lefert, P.; Molina, A. Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to necrotrophic Plectosphaerella cucumerina fungi. Plant J. 2010, 63, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, K.; Fukunaga, S.; Bednarek, P.; Pislewska-Bednarek, M.; Watanabe, S.; Narusaka, Y.; Shirasu, K.; Takano, Y. Glutathione and tryptophan metabolism are required for Arabidopsis immunity during the hypersensitive response to hemibiotrophs. Proc. Natl. Acad. Sci. USA 2013, 110, 9589–9594. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, A.; Pastorczyk, M.; Pislewska-Bednarek, M.; Nishiuchi, T.; Ono, E.; Suemoto, H.; Ishikawa, A.; Frerigmann, H.; Kaido, M.; Mise, K.; et al. Tryptophan-derived metabolites and BAK1 separately contribute to Arabidopsis postinvasive immunity against Alternaria bras-sicicola. Sci. Rep. 2021, 11, 1488. [Google Scholar] [CrossRef]
- Koga, H. Cytological aspects of infection by the rice blast fungus Pyricularia oryzae. In Major Fungal Diseases of Rice: Recent Advances; Sreenivasaprasad, S., Johnson, R., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 87–110. [Google Scholar]
- Ebbole, D.J. Magnaporthe as a Model for Understanding Host-Pathogen Interactions. Annu. Rev. Phytopathol. 2007, 45, 437–456. [Google Scholar] [CrossRef]
- Maeda, K.; Houjyou, Y.; Komatsu, T.; Hori, H.; Kodaira, T.; Ishikawa, A. AGB1 and PMR5 contribute to PEN2-mediated preinvasion resistance to Magnaporthe oryzae in Arabidopsis thaliana. Mol. Plant Microbe Interact. 2009, 22, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Nakao, M.; Nakamura, R.; Kita, K.; Inukai, R.; Ishikawa, A. Non-host resistance to penetration and hyphal growth of Magnaporthe oryzae in Arabidopsis. Sci. Rep. 2011, 1, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozaki, M.; Kita, K.; Kodaira, T.; Ishikawa, A. AtRbohF contributes to non-host resistance to Magnaporthe oryzae in Arabidopsis. Biosci. Biotechnol. Biochem. 2013, 77, 1323–1325. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Murano, T.; Ishikawa, A. SOBIR1 and AGB1 independently contribute to nonhost resistance to Pyricularia oryzae (syn. Magnaporthe oryzae) in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2018, 82, 1922–1930. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, Y.; Makihara, M.; Ishikawa, A. Leaf age and time of inoculation contribute to nonhost resistance to Pyricularia oryzae in Arabidopsis thaliana. Plant. Biotechnol. 2017, 34, 207–210. [Google Scholar] [CrossRef]
- Yamaura, S.; Yamauchi, Y.; Makihara, M.; Yamashino, T.; Ishikawa, A. CCA1 and LHY contribute to nonhost resistance to Pyricularia oryzae (syn. Magnaporthe oryzae) in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2020, 84, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Yoshino, R.; Mogi, S. The effects of light and darkness on the infection of rice blast fungus, Pyricularia oryzae. Proc. Assoc. Plant Prot. Hokuriku 1974, 22, 3–6. [Google Scholar]
- Xie, X.Z.; Xue, Y.J.; Zhou, J.J.; Zhang, B.; Chang, H.; Takano, M. Phytochromes Regulate SA and JA Signaling Pathways in Rice and Are Required for Developmentally Controlled Resistance to Magnaporthe grisea. Mol. Plant 2011, 4, 688–696. [Google Scholar] [CrossRef]
- Griebel, T.; Zeier, J. Light regulation and daytime dependency of inducible plant defenses in arabidopsis: Phytochrome signaling controls systemic acquired resistance rather than local defense. Plant Physiol. 2008, 147, 790–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staswick, P.E.; Tiryaki, I.; Rowe, M.L. Jasmonate Response Locus JAR1 and Several Related Arabidopsis Genes Encode Enzymes of the Firefly Luciferase Superfamily That Show Activity on Jasmonic, Salicylic, and Indole-3-Acetic Acids in an Assay for Adenylation. Plant Cell 2002, 14, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, J.; Chen, W.J.; Estes, B.; Chang, H.S.; Nawrath, C.; Metraux, J.P.; Zhu, T.; Katagiri, F. Topology of the network inte-grating salicylate and jasmonate signal transduction derived from global expression phenotyping. Plant J. 2003, 34, 217–228. [Google Scholar] [CrossRef]
- Torres, M.A.; Dangl, J.L.; Jones, J.D.G. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Pathogen-induced, NADPH oxidase-derived reactive oxygen intermediates suppress spread of cell death in Arabidopsis thaliana. Nat. Genet. 2005, 37, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Liebrand, T.W.; Burg, H.A.V.D.; Joosten, M.H. Two for all: Receptor-associated kinases SOBIR1 and BAK1. Trends Plant. Sci. 2014, 19, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Shibuya, H.; Ishikawa, A. SOBIR1 contributes to non-host resistance to Magnaporthe oryzae in Arabidopsis. Biosci. Biotechnol. Biochem. 2016, 80, 1577–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, S.; Yamauchi, Y.; Ishikawa, A. Photoperiod Following Inoculation of Arabidopsis with Pyricularia oryzae (syn. Magnaporthe oryzae) Influences on the Plant–Pathogen Interaction. Int. J. Mol. Sci. 2021, 22, 5004. https://doi.org/10.3390/ijms22095004
Shimizu S, Yamauchi Y, Ishikawa A. Photoperiod Following Inoculation of Arabidopsis with Pyricularia oryzae (syn. Magnaporthe oryzae) Influences on the Plant–Pathogen Interaction. International Journal of Molecular Sciences. 2021; 22(9):5004. https://doi.org/10.3390/ijms22095004
Chicago/Turabian StyleShimizu, Sayaka, Yuri Yamauchi, and Atsushi Ishikawa. 2021. "Photoperiod Following Inoculation of Arabidopsis with Pyricularia oryzae (syn. Magnaporthe oryzae) Influences on the Plant–Pathogen Interaction" International Journal of Molecular Sciences 22, no. 9: 5004. https://doi.org/10.3390/ijms22095004
APA StyleShimizu, S., Yamauchi, Y., & Ishikawa, A. (2021). Photoperiod Following Inoculation of Arabidopsis with Pyricularia oryzae (syn. Magnaporthe oryzae) Influences on the Plant–Pathogen Interaction. International Journal of Molecular Sciences, 22(9), 5004. https://doi.org/10.3390/ijms22095004