
Aberrant Early in Life Stimulation of the Stress-Response System Affects Emotional Contagion and Oxytocin Regulation in Adult Male Mice

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Body Weight Monitoring
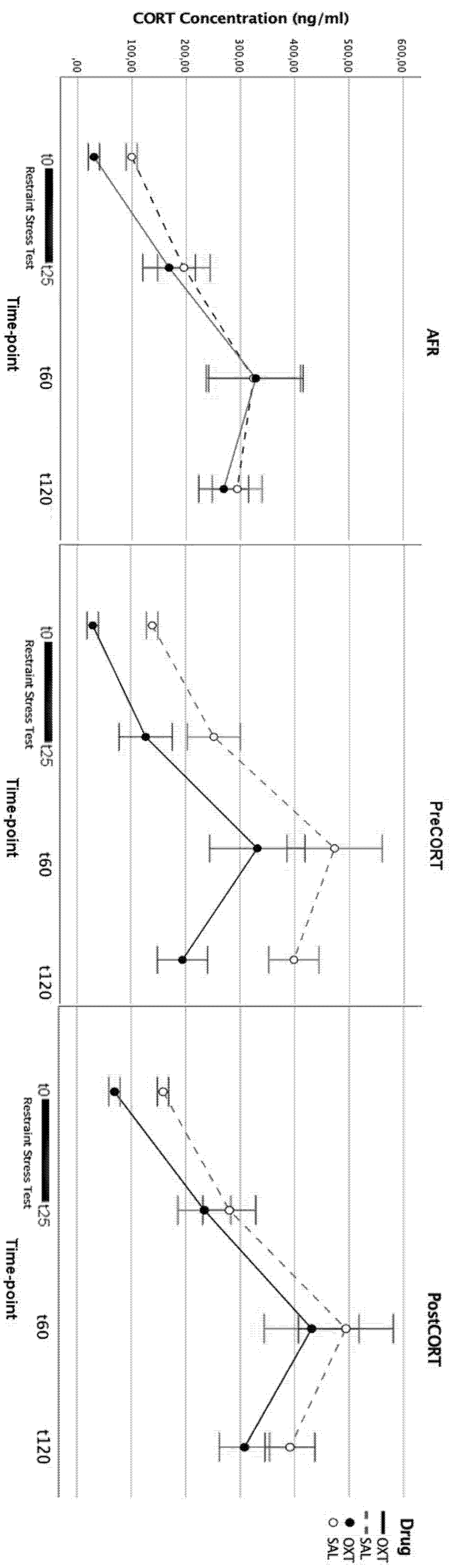
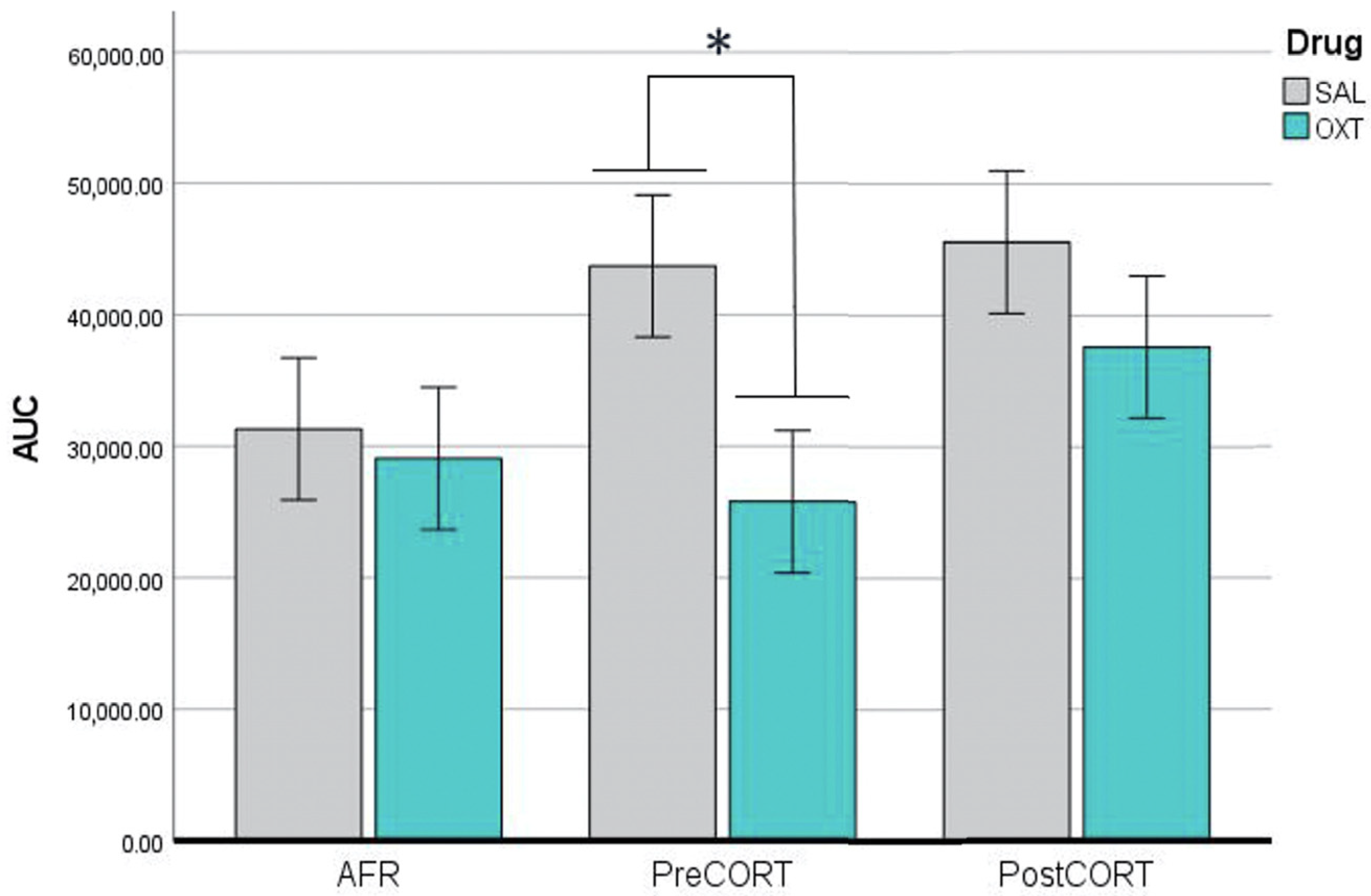
2.2. Plasma Corticosterone Assessment
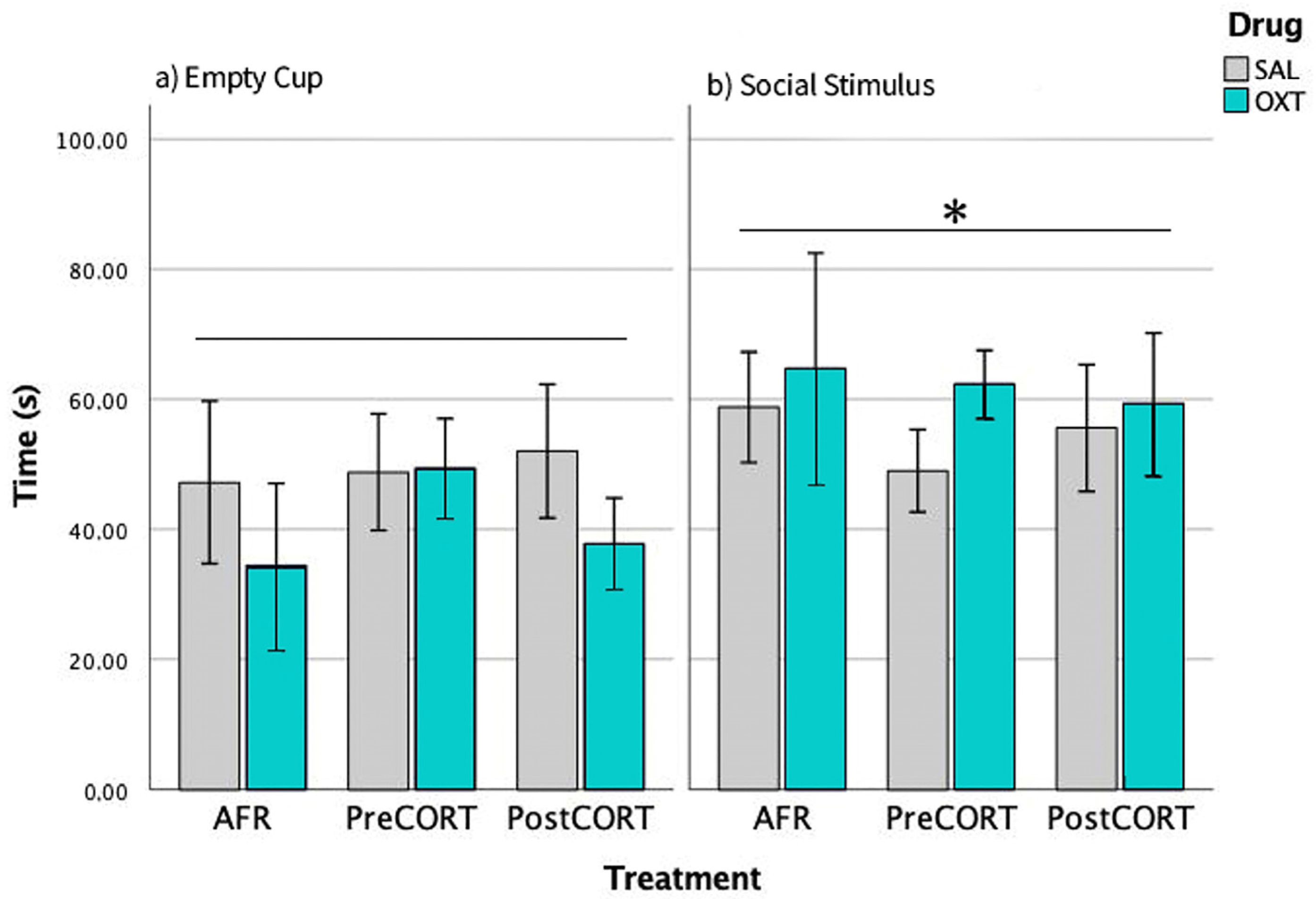
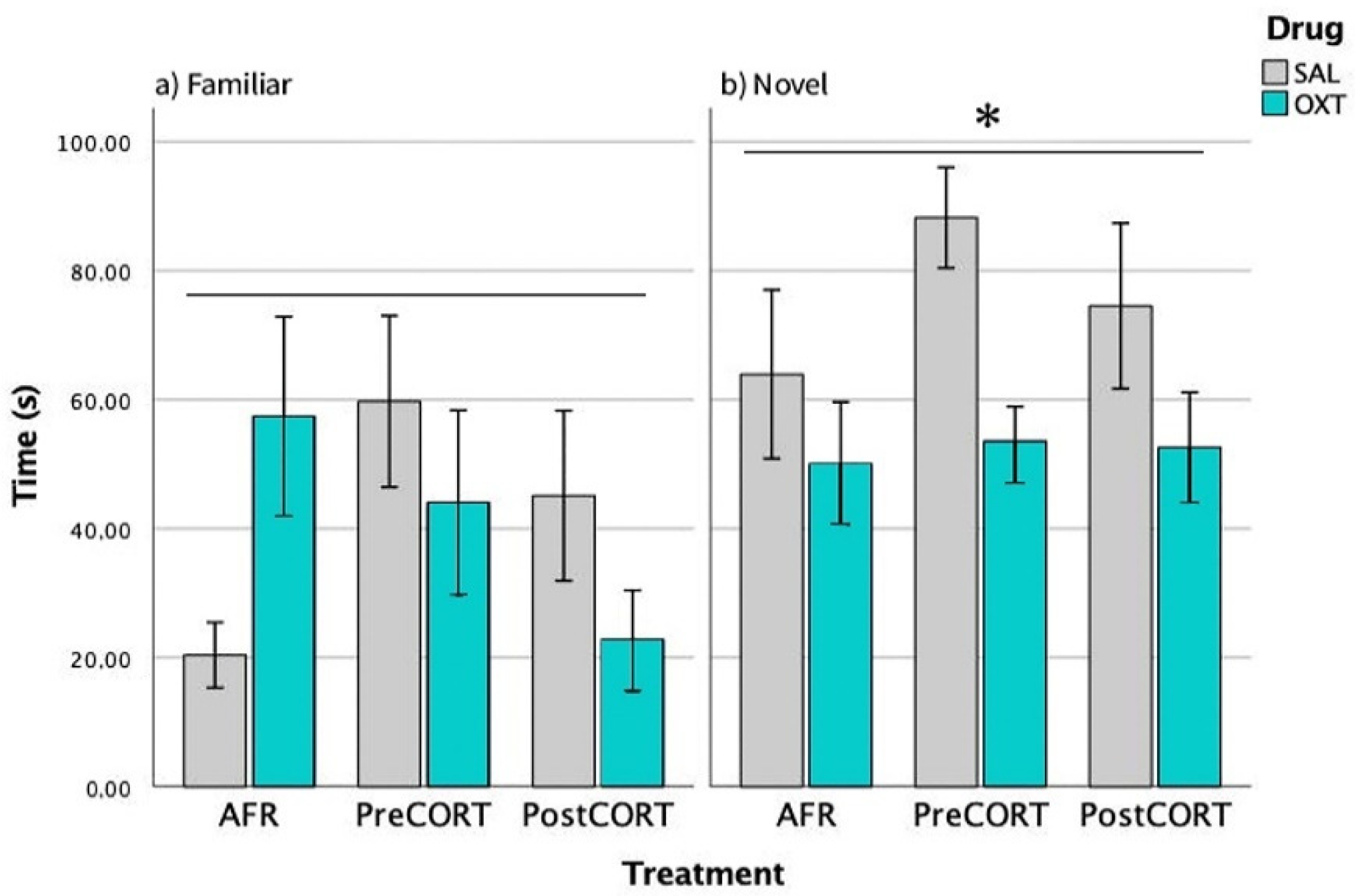
2.3. Sociability Test
2.3.1. Episode 1—Habituation Phase
2.3.2. Episode 2—Social Preference
2.3.3. Episode 3—Social Novelty Preference
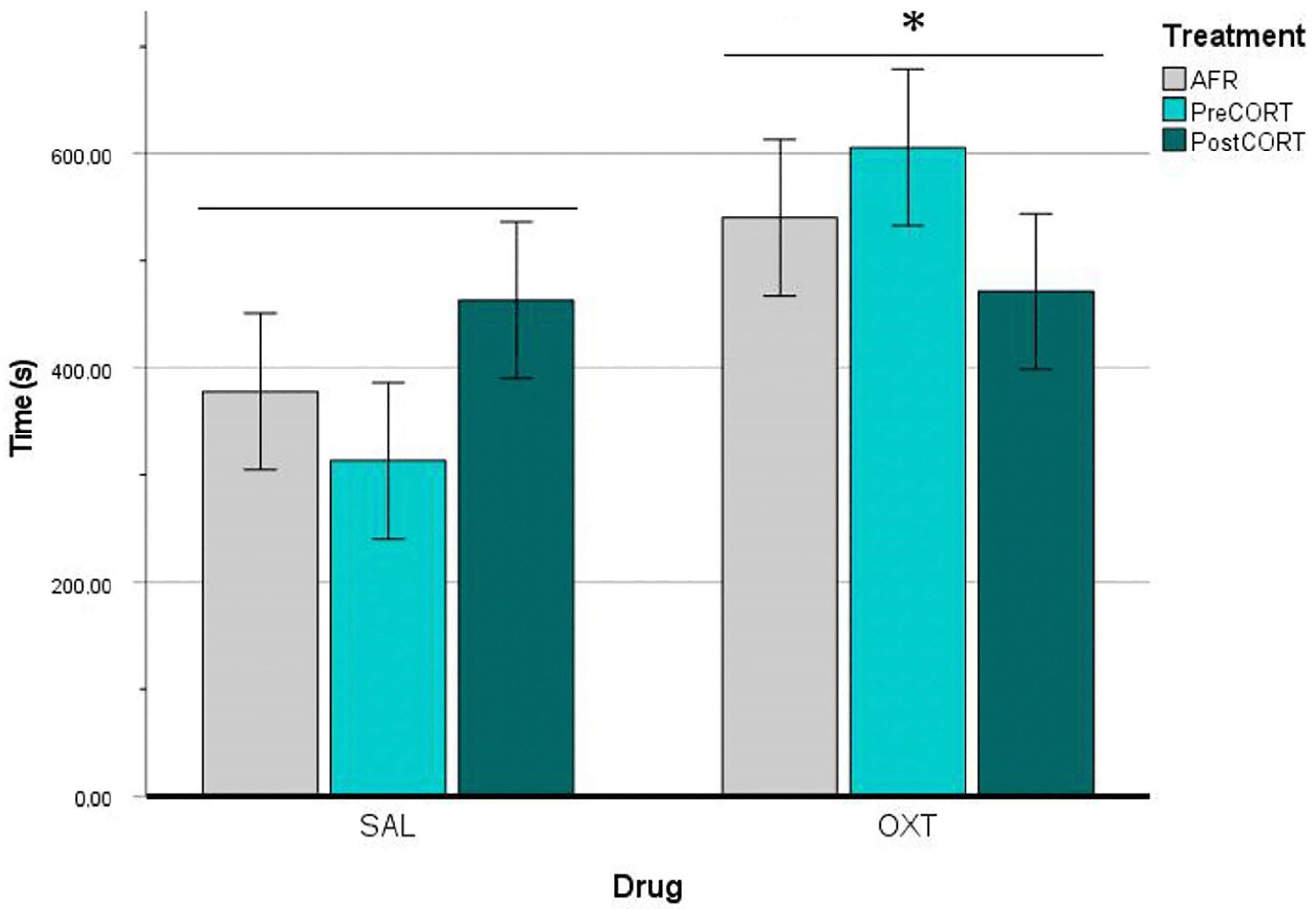
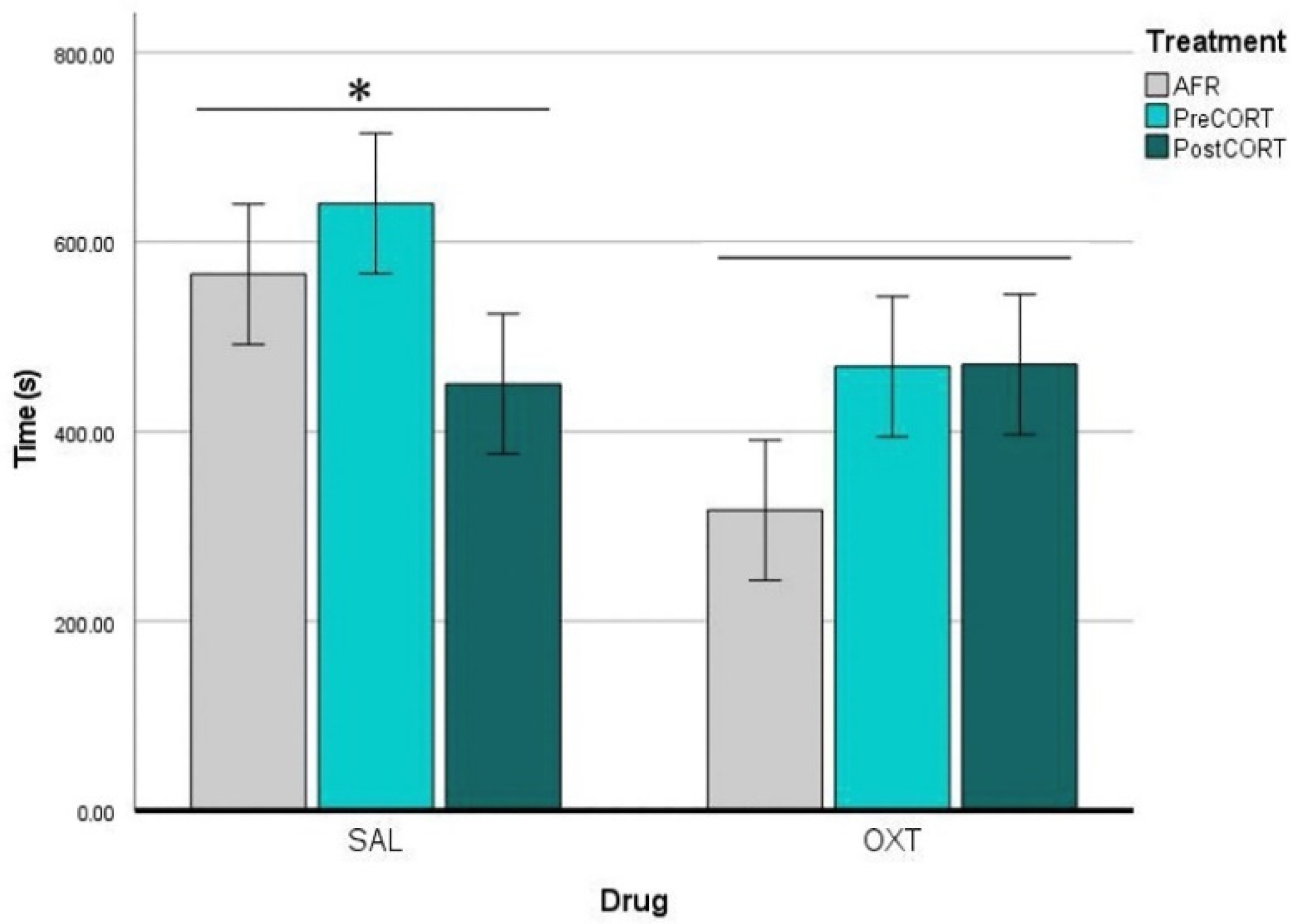
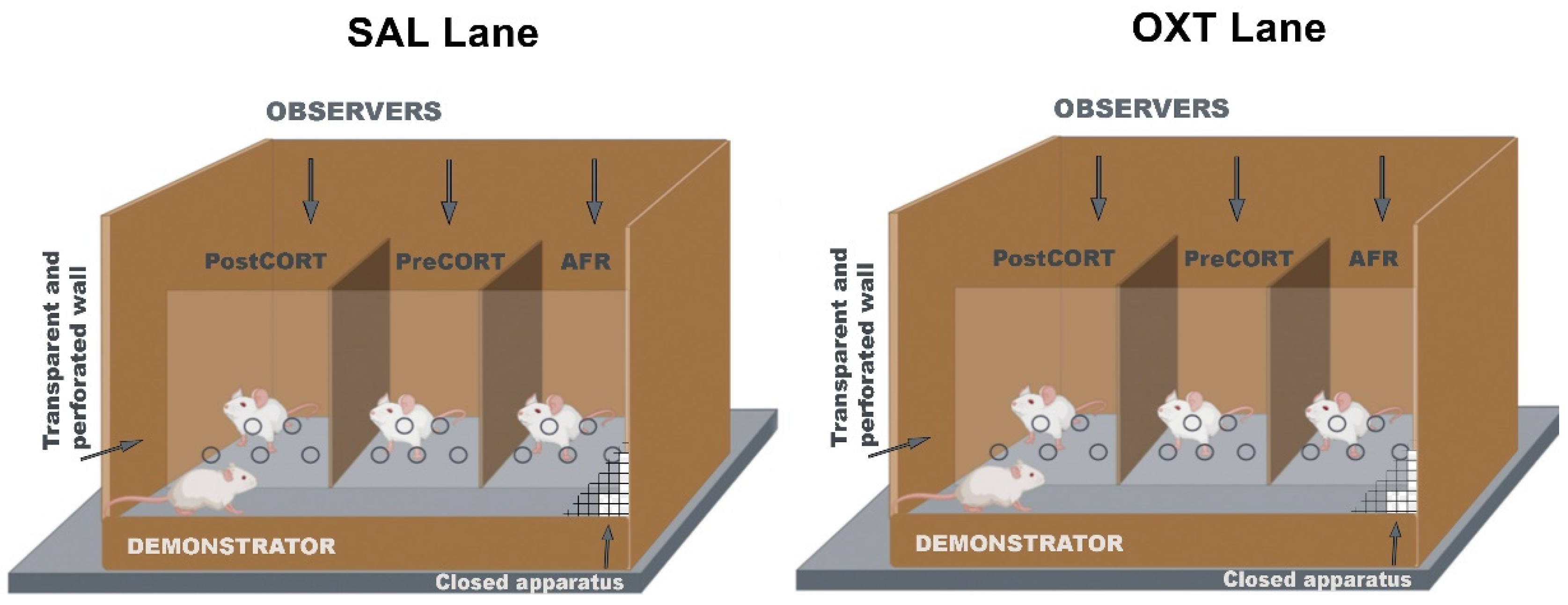
2.4. Emotional Contagion Assay
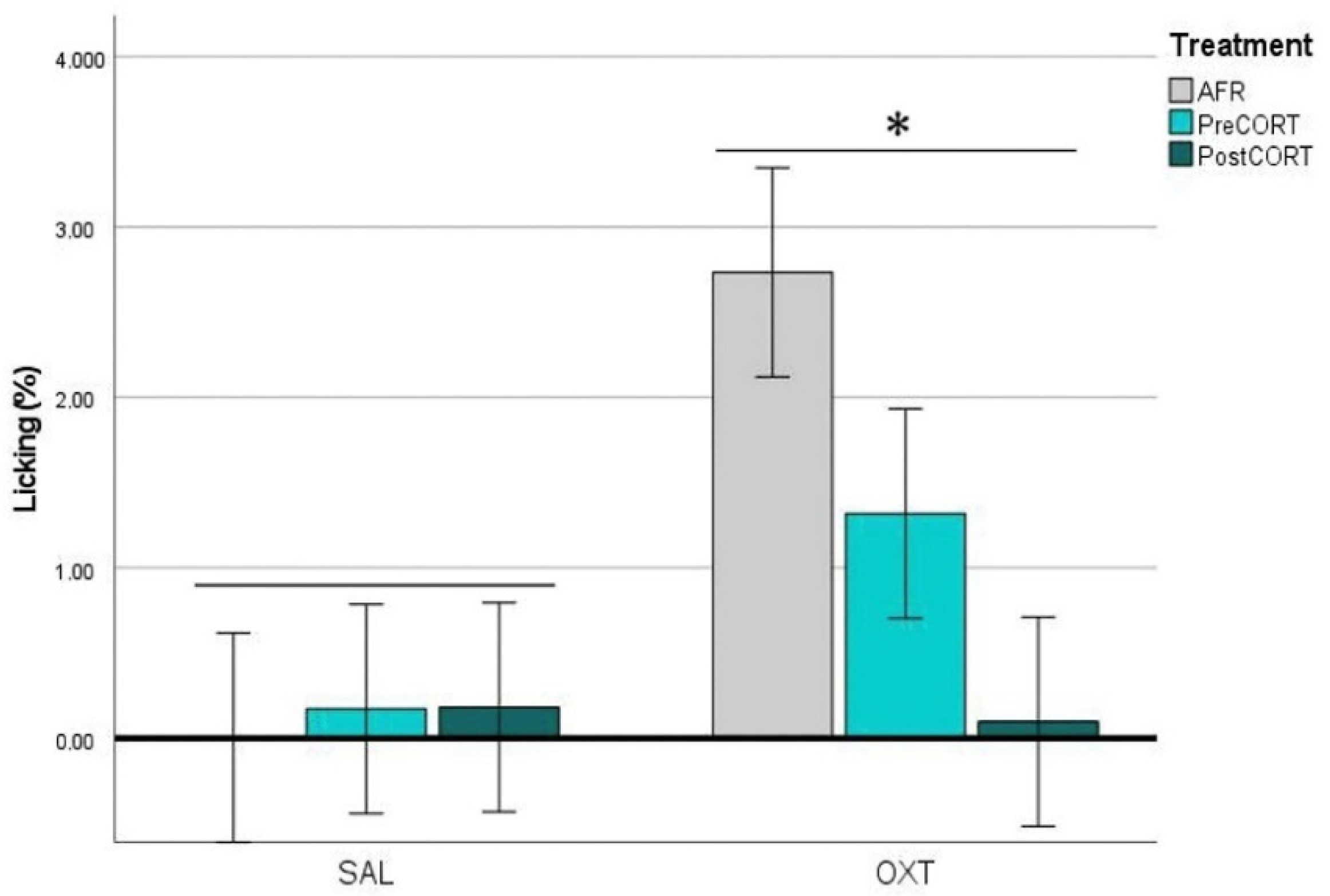
2.4.1. Observers’ Paw-Licking Behaviour
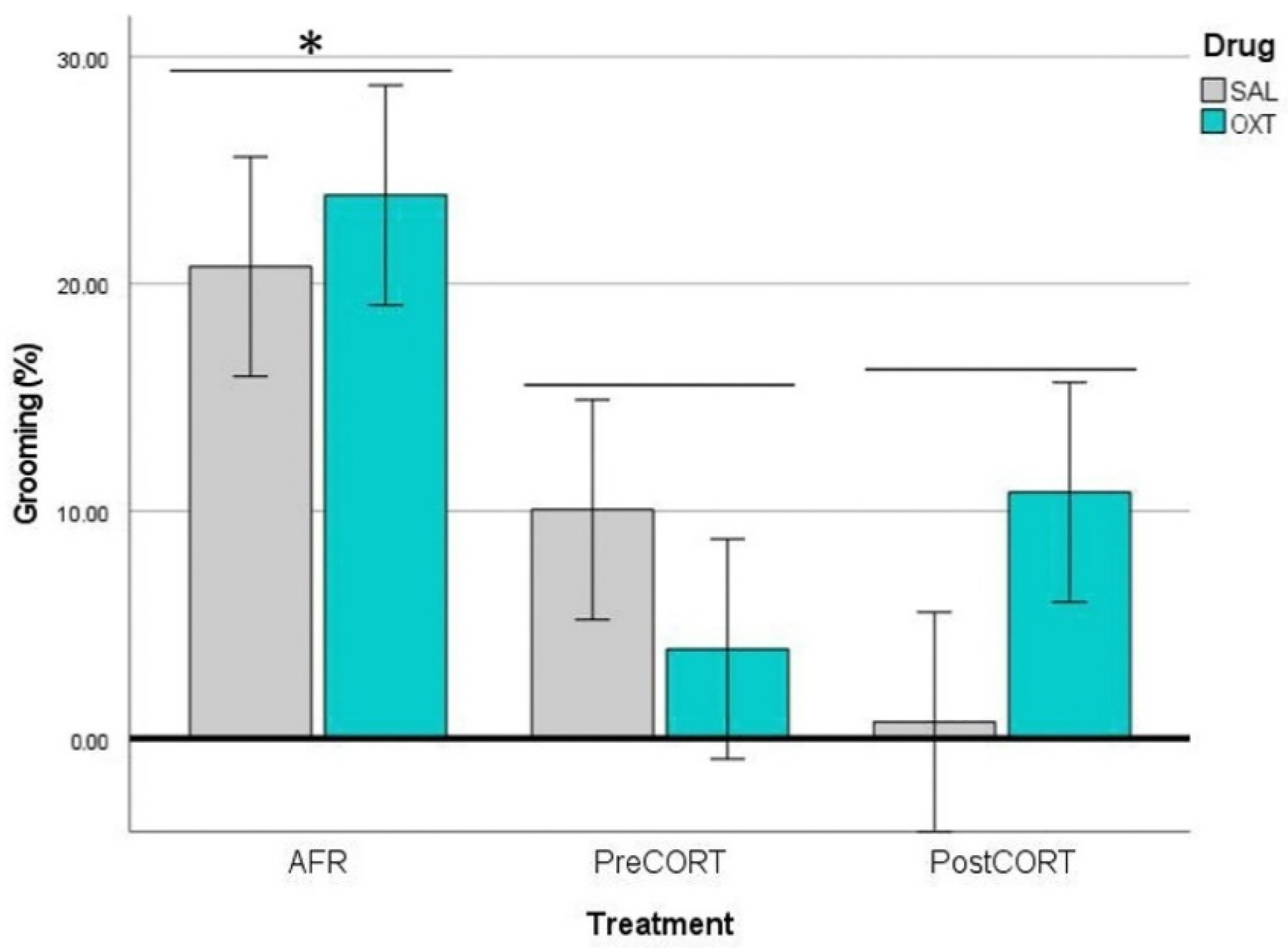
2.4.2. Observers’ Self-Grooming
2.4.3. Observers’ Hole-Probe Behaviour
2.4.4. Demonstrators’ Paw-Licking Behaviour
2.4.5. Demonstrators’ Self-Grooming
2.4.6. Demonstrators’ Hole-Probe Behaviour
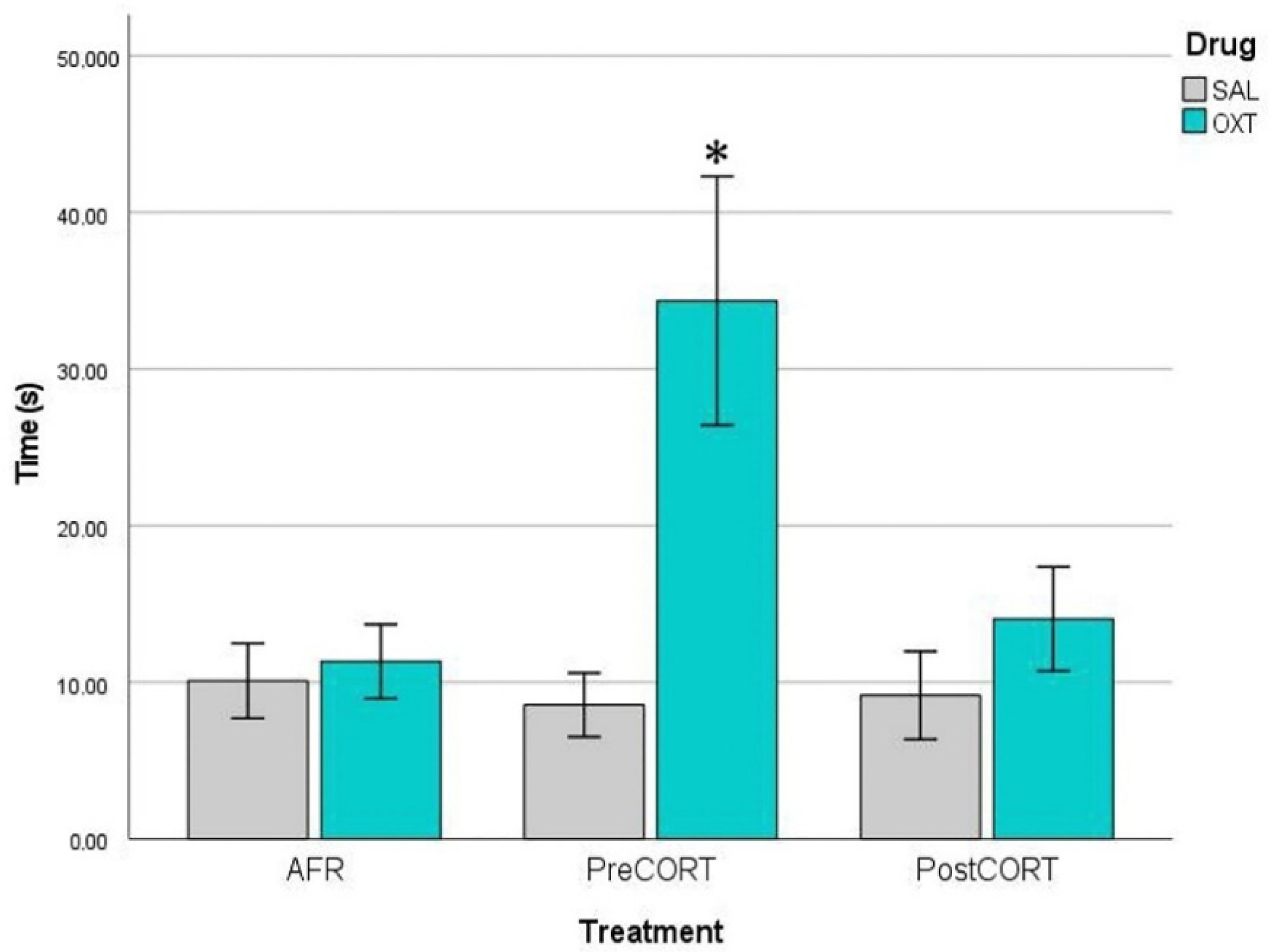
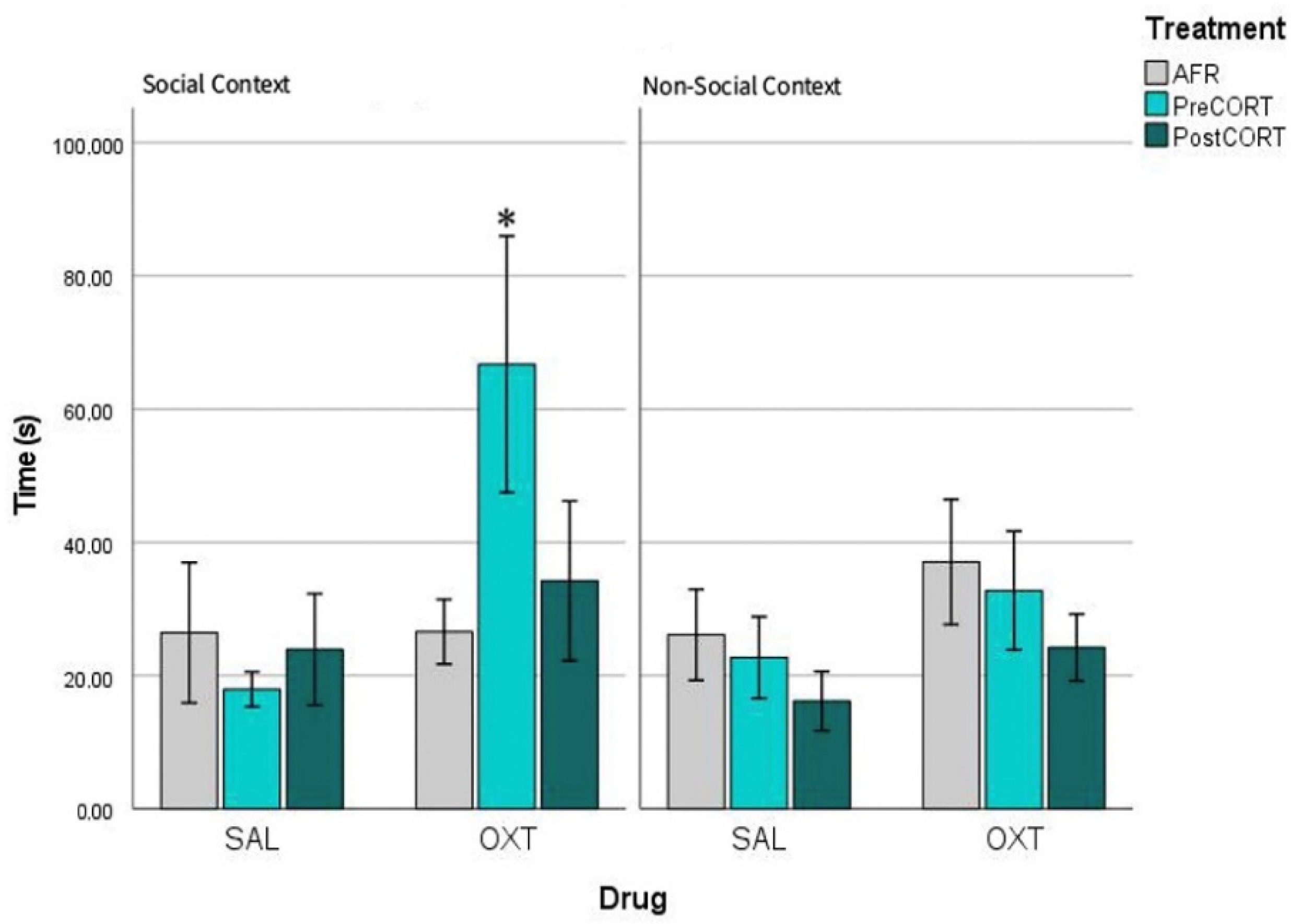
2.5. Profiles of Pain Sensitivity and of Response to Oxytocin in a Social vs. a Non-Social Context
2.6. Molecular Analyses
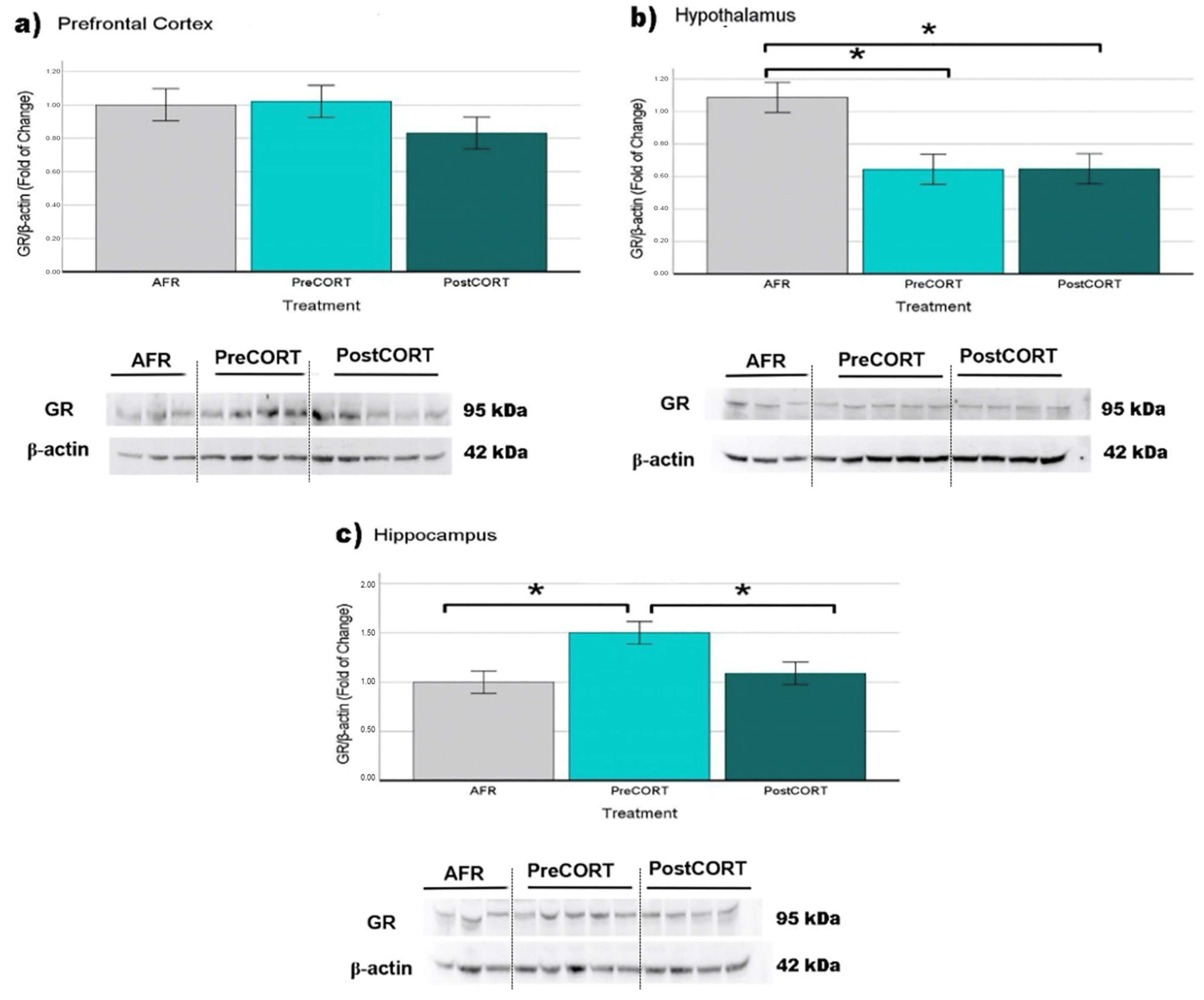
2.6.1. Glucocorticoid Receptors (GR)
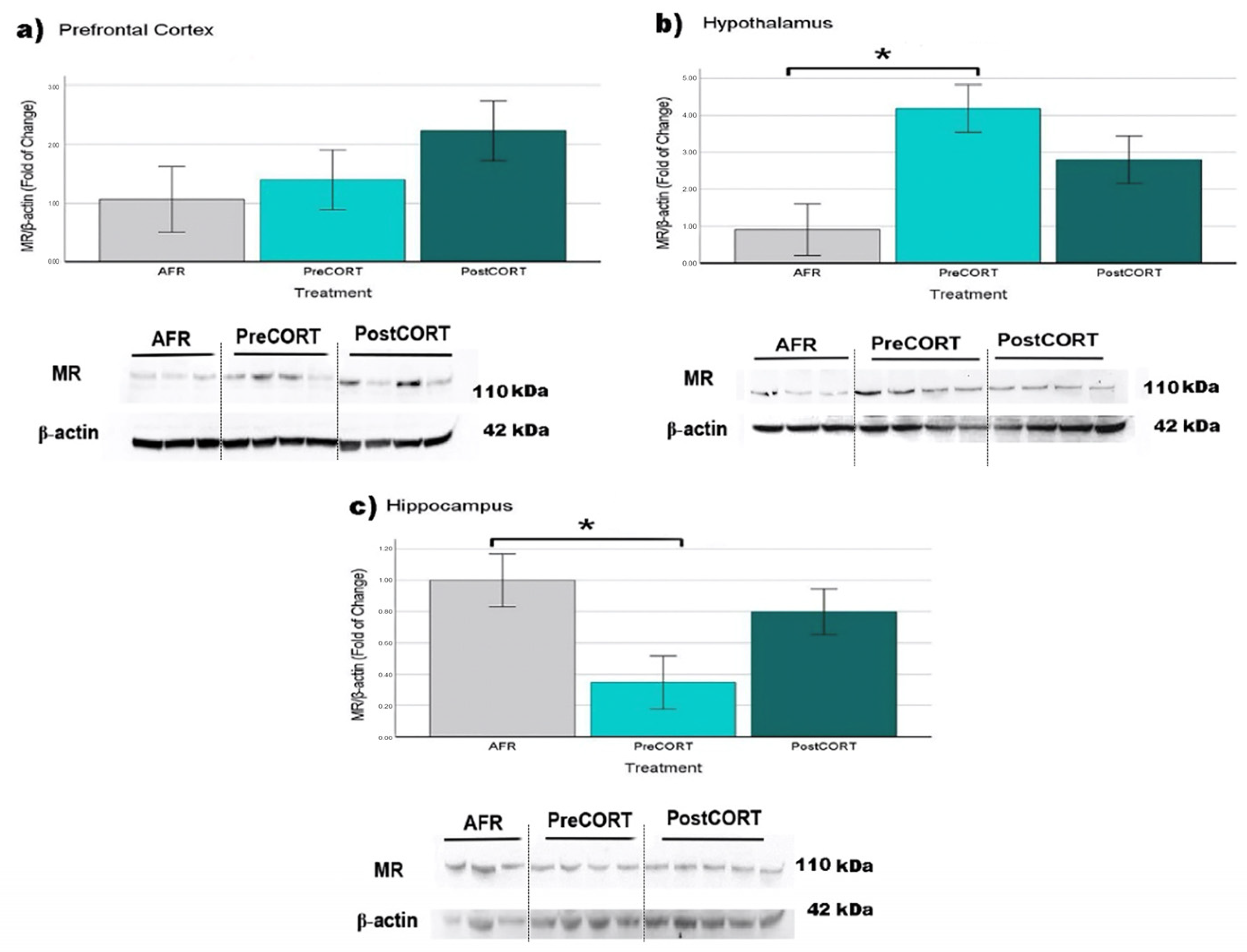
2.6.2. Mineralocorticoid Receptors (MR)
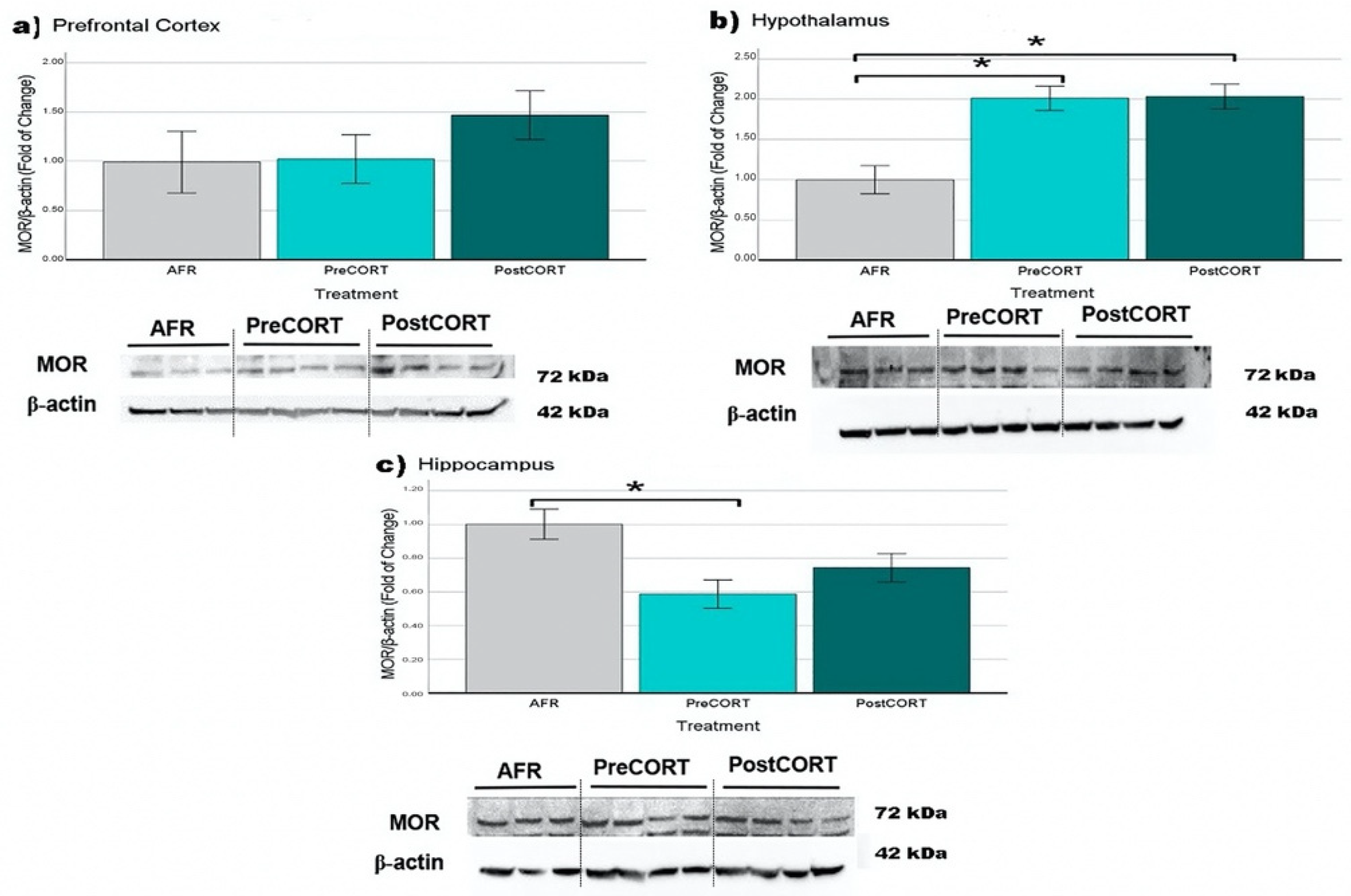
2.6.3. µ-Opioid Receptors (MOR)
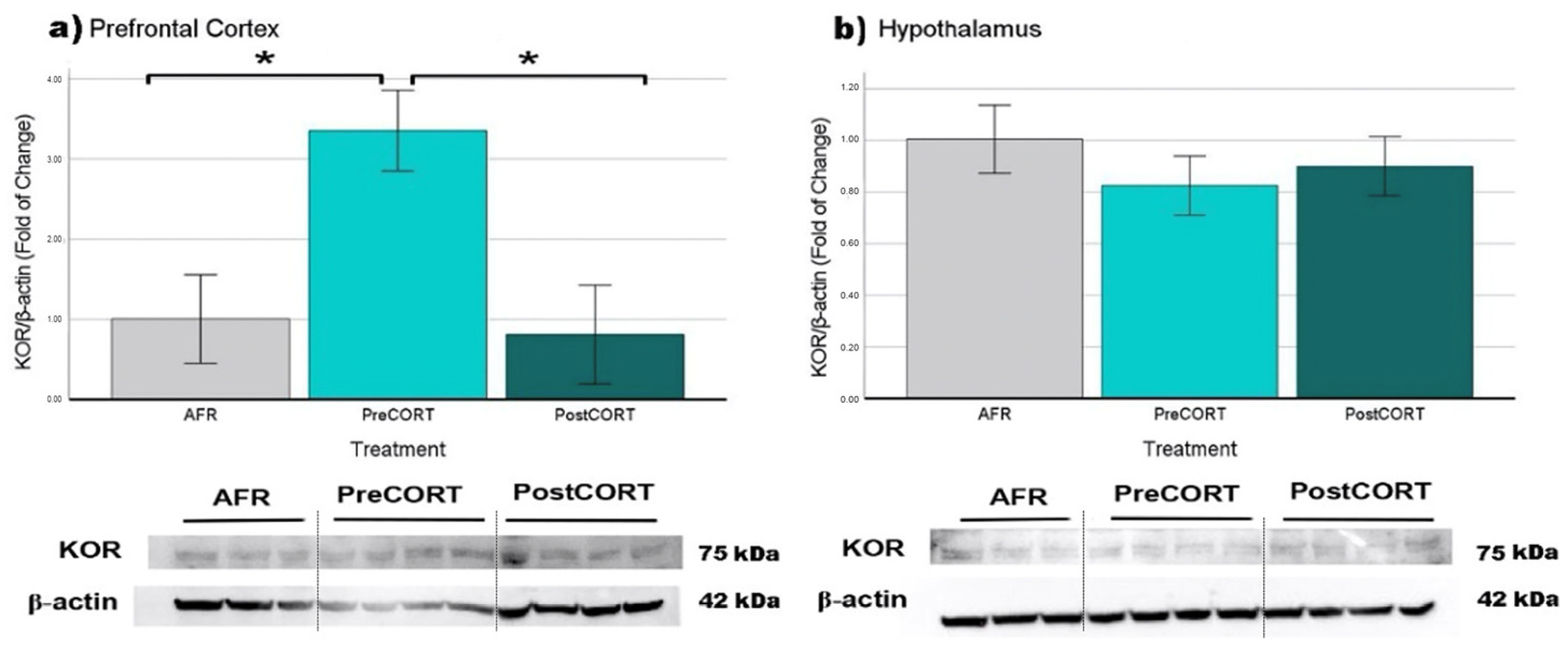
2.6.4. κ-Opioid Receptors (KOR)
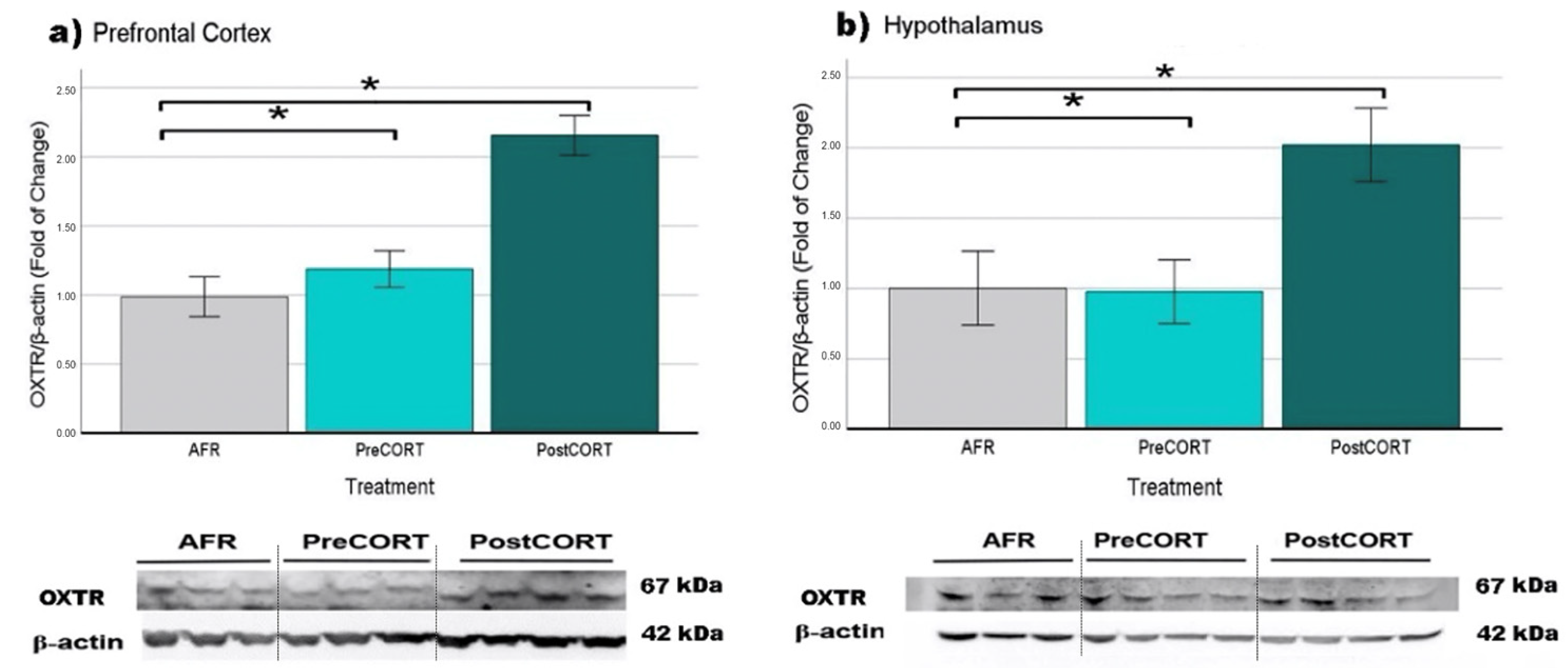
2.6.5. Oxytocin Receptors (OXTR)
3. Discussion
3.1. Titers of Plasma Corticosterone and Its Receptors in Brain Areas of Adult Offspring
3.2. Profiles of Sociability and Modulation by Oxytocin
3.3. Profiles of Emotional Contagion and Involvement of Oxytocin Regulation
3.3.1. Observers’ Point of View
3.3.2. Demonstrator’s Point of View
3.4. Profiles of Pain Sensitivity and Effects of Oxytocin as a Function of Test Setting
3.5. Molecular Analyses in Brain Areas of Adult Offspring
3.6. Modulation of Profiles of Opioid Receptors in Adult Offspring
4. Materials and Methods
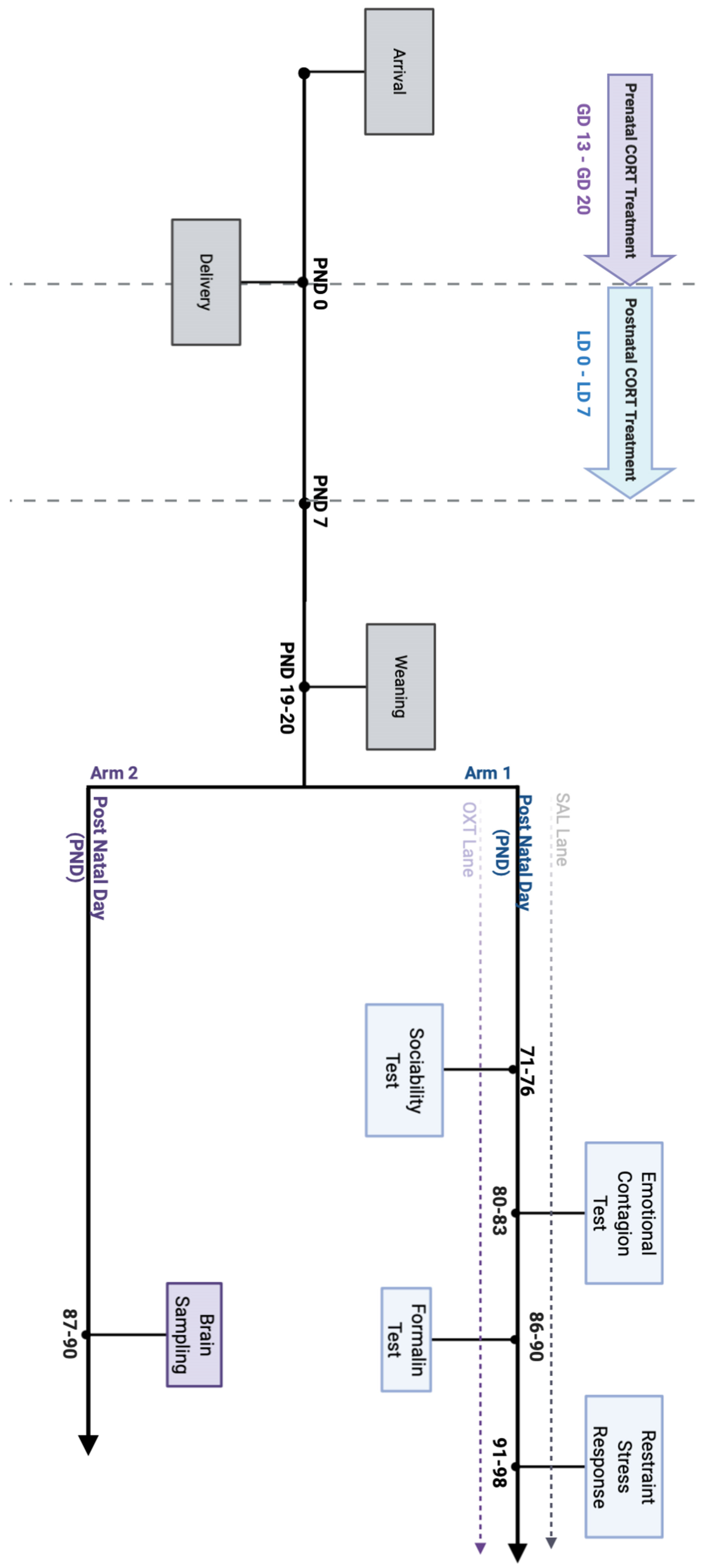
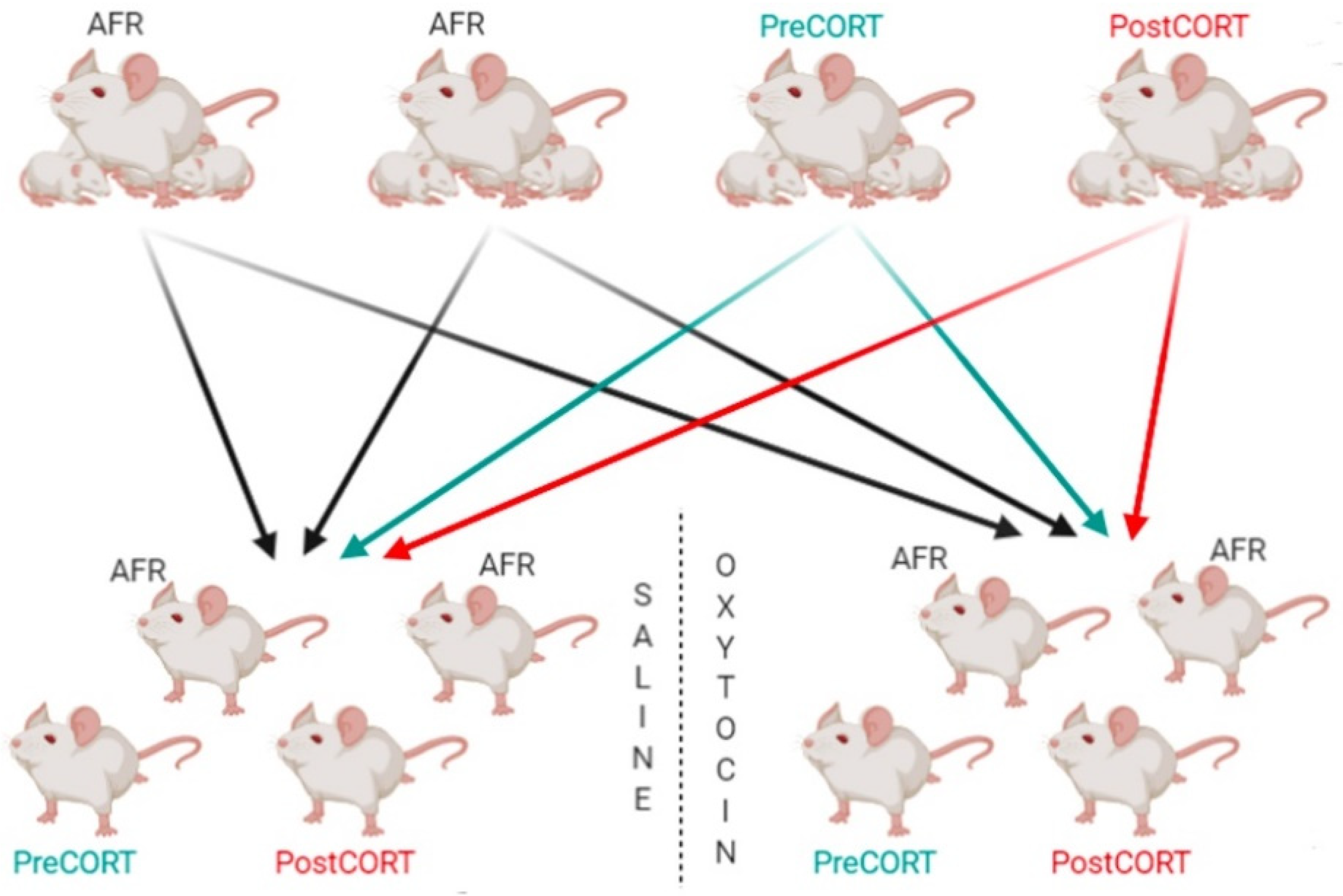
4.1. Animal Rearing and Conditions
4.2. Experimental Timeline
4.3. Oxytocin Administration
4.4. Sociability Test
- Episode 1—Habituation Phase: Mice were individually placed in a black Plexiglas® box (20 × 40 cm2 and 40 cm height) containing two identical empty wire cups (10 × 10 cm2 and 11 cm height; pencil holder, IKEA) placed close to the two opposite corners of the apparatus. During this phase, no social stimuli were present, and the experimental mice were free to explore the entire arena.
- Episode 2—Social Preference: One unfamiliar male mouse was placed under a wire cup, serving as social stimulus (Stranger 1), whilst the other cup remained empty. The position of the social stimulus was counterbalanced across subjects. The preference for the cup containing the social stimulus (Stranger 1) over the empty cup was evaluated. In addition, further analysis concerned the interaction with the social stimulus only.
- Episode 3—Social Novelty Preference: An additional unfamiliar male mouse (Stranger 2) was placed in the previously empty cup, whilst the other cup contained the mouse encountered during Ep. 2 (Stranger 1). The preference for the cup containing the novel social stimulus (Stranger 2) over the cup containing the familiar social stimulus (Stranger 1) was evaluated. Moreover, further analysis concerned the interaction with the novel stimulus only.
4.5. Emotional Contagion Assay
4.6. Pain Sensitivity in the Formalin Test
4.7. Restraint Stress and Assessment of Plasma Corticosterone
4.8. Molecular Analyses from Adult Offspring’s Brain Areas
Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gluckman, P.D.; Hanson, M.A. Living with the past: Evolution, development, and patterns of disease. Science 2004, 305, 1733–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babenko, O.; Kovalchuk, I.; Metz, G.A. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neurosci. Biobehav. R 2015, 48, 70–91. [Google Scholar] [CrossRef] [PubMed]
- Laviola, G.; Macrì, S. Adaptive and Maladaptive Aspects of Developmental Stress; Springer: Berlin/Heidelberg, Germany, 2013; Volume 3. [Google Scholar]
- Brown, E.S.; Varghese, F.P.; McEwen, B.S. Association of Depression with Med Illness: Does Cortisol Play a Role? Biol. Psychiat. 2004, 55, 1–9. [Google Scholar] [CrossRef]
- Bunea, I.M.; Szentágotai-Tătar, A.; Miu, A.C. Early-life adversity and cortisol response to social stress: A meta-analysis. Transl. Psychiat. 2017, 7, 1274. [Google Scholar] [CrossRef]
- Laurent, H.K.; Neiderhiser, J.M.; Natsuaki, M.N.; Shaw, D.S.; Fisher, P.A.; Reiss, D.; Leve, L.D. Stress system development from age 4.5 to 6: Family environment predictors and adustment implications of HPA activity stability versus change. Dev. Psychobiol. 2014, 56, 340–354. [Google Scholar] [CrossRef]
- Nishi, M.; Horii-Hayashi, N.; Sasagawa, T. Effects of early life adverse experiences on the brain: Implications from maternal separation models in rodents. Front. Neurosci. 2014, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Wehmeier, P.M.; Schacht, A.; Barkley, R.A. Social and emotional impairment in children and adolescents with ADHD and the impact on quality of life. J. Adolesc. Health 2010, 46, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Myers, W.C.; Burket, R.C.; Otto, T.A. Conduct Disorder and Personality Disorders in Hospitalized Adolescents. J. Clin. Psychiatry 1993, 54, 21–26. [Google Scholar] [PubMed]
- Salmanian, M.; Mohammadi, M.R.; Keshavarzi, Z.; Brand, S. An update on the global prevalence of conduct disorder (2011-2017): Study protocol for a systematic review and meta-analysis. J. Forensic Leg. Med. 2018, 59, 1–3. [Google Scholar] [CrossRef]
- Szentiványi, D.; Halász, J.; Horváth, L.O.; Kocsis, P.; Miklósi, M.; Vida, P.; Balázs, J. Quality of life of adolescents with conduct disorder: Gender differences and comorbidity with oppositional defiant disorder. Psychiatr. Hung. 2019, 34, 280–286. [Google Scholar]
- Franklin, T.B.; Linder, N.; Russig, H.; Thony, B.; Mansuy, I.M. Influence of Early Stress on Social Abilities and Serotonergic Functions Across Generations in Mice. PLoS ONE 2011, 6, e21842. [Google Scholar] [CrossRef] [Green Version]
- De Souza, M.A.; Centenaro, L.A.; Rossi Menegotto, P.; Pereira Henriques, T.; Bonini, J.; Achaval, M.; Bolten Lucion, A. Prenatal Stress produces Social Behavior Deficits and Alters the Number of Oxytocin and Vasopressin Neurons in Adult Rat. Neurochem. Res. 2013, 38, 1479–1489. [Google Scholar] [CrossRef]
- Torner, L.; Toschi, N.; Nava, G.; Clapp, C.; Neumann, I.D. Increased Hypothalamic Expression of Prolactin in Lactation: Involvement in Behavioural and Neuroendocrine Stress Responses. Eur. J. Neurosci. 2002, 15, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.J. Central noradrenergic mechanisms underlying acute stress responses of the Hypothalamo-pituitary-adrenal axis: Adaptations through pregnancy and lactation. Stress 2005, 8, 5–18. [Google Scholar] [CrossRef]
- Slattery, D.A.; Neumann, I.D. No stress please! Mechanisms of stress hyporesponsiveness of the maternal brain. J. Physiol. 2008, 586, 377–385. [Google Scholar] [CrossRef]
- Rosenfeld, P.; Suchecki, D.; Levine, S. Multifactorial regulation of the hypothalamic-pituitary-adrenal axis during development. Neurosci. Biobehav. Rev. 1992, 16, 553–568. [Google Scholar] [CrossRef]
- Cirulli, F.; Berry, A.; Alleva, E. Early disruption of the mother-infant relationship: Effects on brain plasticity and implications for psychopathology. Neurosci. Biobehav. Rev. 2003, 27, 73–82. [Google Scholar] [CrossRef]
- Welberg, L.A.M.; Seckl, J.R. Prenatal Stress, Glucocorticoids and the Programming of the Brain. J. Neuroendocrinol. 2001, 13, 113–128. [Google Scholar] [CrossRef]
- Lindsay, R.S.; Lindsay, R.M.; Waddell, B.J.; Seckl, J.R. Prenatal glucocorticoid exposure leads to offspring hyperglycaemia in the rat: Studies with the 11 beta-hydroxysteroid dehydrogenase inhibitor carbenoxolone. Diabetologia 1996, 39, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Plotsky, P.M.; Meaney, M.J. Early, postnatal experience alters hypothalamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF content and stress-induced release in adult rats. Mol. Brain Res. 1993, 18, 195–200. [Google Scholar] [CrossRef]
- Casolini, P.; Cigliana, G.; Alemà, G.S.; Ruggieri, V.; Angelucci, L.; Catalani, A. Effect of increased maternal corticosterone during lactation on hippocampal corticosteroid receptors, stress response and learning in offspring in the early stages of life. Neuroscience 1997, 79, 1005–1012. [Google Scholar] [CrossRef]
- Catalani, A.; Casolini, P.; Scaccianoce, S.; Patacchioli, F.R.; Spinozzi, P.; Angelucci, L. Maternal corticosterone during lactation permanently affects brain corticosteroid receptors, stress response and behaviour in rat progeny. Neuroscience 2000, 100, 319–325. [Google Scholar] [CrossRef]
- Catalani, A.; Alemà, G.S.; Cinque, C.; Zuena, A.R.; Casolini, P. Maternal corticosterone effects on hypothalamus-pituitary-adrenal axis regulation and behavior of the offspring in rodents. Neurosci. Biobehav. Rev. 2011, 35, 1502–1517. [Google Scholar] [CrossRef]
- Meaney, M.J.; Sapolski, R.M.; Mc Ewen, R. The Development of the Glucocorticoid Receptor System in the Rat Limbic Brain—Ontogeny and Autoregulation. Dev. Brain Res. 1985, 18, 159–164. [Google Scholar] [CrossRef]
- Owen, D.; Matthews, S.G. Glucocorticoids and sex-dependent development of brain glucocorticoid and mineralocorticoid receptors. Endocrinology 2003, 144, 2775–2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macrì, S.; Granstrem, O.; Shumilina, M.; Antunes Gomes dos Santos, F.J.; Berry, A.; Saso, L.; Laviola, G. Resilience and vulnerability are dose-dependently related to neonatal stressors in mice. Horm. Behav. 2009, 56, 391–398. [Google Scholar] [CrossRef]
- Macrì, S.; Zoratto, F.; Laviola, G. Early-stress regulates resilience, vulnerability and experimental validity in laboratory rodents through mother-offspring hormonal transfer. Neurosci. Biobehav. Rev. 2011, 35, 1534–1543. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, I.E. Empathy in Autism Spectrum Disorder. J. Autism Dev. Disord. 2019, 49, 3939–3955. [Google Scholar] [CrossRef]
- Veenema, A.H.; Neumann, I.D. Neurobiological mechanisms of aggression and stress coping: A comparative study in mouse and rat selection lines. Brain Behav. Evol. 2007, 70, 274–285. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, Diagnostic and Statistical Manual of Mental Disorder, 5th ed.; (DSM-5); Psychiatric Association: Arlington, VA, USA, 2013. [Google Scholar]
- Preston, S.D.; de Waal, F.B.M. Empathy: Its ultimate and proximate bases. Behav. Brain Sci. 2002, 25, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Brothers, L. The Neural Basis of Primate Social Communication Motivation and Emotion; Springer: Berlin/Heidelberg, Germany, 1990; Volume 14, pp. 81–91. [Google Scholar]
- Martin, L.J.; Hathaway, G.; Isbester, K.; Mirali, S.; Acland, E.L.; Niederstrasser, N.; Slepian, P.M.; Trost, Z.; Bartz, J.A.; Sapolsky, R.M.; et al. Reducing social stress elicits emotional contagion of pain in mouse and human strangers. Curr. Biol. 2015, 25, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Sala, M.; Braida, D.; Lentini, D.; Busnelli, M.; Bulgheroni, E.; Capurro, V.; Finardi, A.; Donzelli, A.; Pattini, L.; Rubino, T.; et al. Pharmacologic rescue of impaired cognitive flexibility, social deficits, increased aggression, and seizure susceptibility in oxytocin receptor null mice: A neurobehavioral model of autism. Biol. Psychiatry 2011, 69, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Monakhov, M.; Pratt, M.; Ebstein, R.P. Oxytocin Pathway Genes: Evolutionary Ancient System Impacting on Human Affiliation, Sociality, and Psychopathology. Biol. Psychiatry 2016, 79, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Shamay-Tsoory, S.; Young, L.J. Understanding the Oxytocin System and Its Relevance to Psychiatry. Biol. Psychiatry 2016, 79, 150–152. [Google Scholar] [CrossRef] [Green Version]
- Veenema, A.H.; Neumann, I.D. Central vasopressin and oxytocin release: Regulation of complex social behaviours. Prog. Brain Res. 2008, 170, 261–276. [Google Scholar] [PubMed]
- Baracz, S.J.; Everett, N.A.; Cornish, J.L. The impact of early life stress on the central oxytocin system and susceptibility for drug addiction: Applicability of oxytocin as a pharmacotherapy. Neurosci. Biobehav. Rev. 2020, 110, 114–132. [Google Scholar] [CrossRef]
- Romano, A.; Tempesta, B.; Di Bonaventura, M.V.M.; Gaetani, S. From autism to eating disorders and more: The role of oxytocin in neuropsychiatric disorders. Front. Neurosci. 2016, 9, 497. [Google Scholar] [CrossRef] [Green Version]
- Sippel, L.M.; Allington, C.E.; Pietrzak, R.H.; Harpaz-Rotem, I.; Mayes, L.C.; Olff, M. Oxytocin and Stress-related Disorders: Neurobiological Mechanisms and Treatment Opportunities. Chronic Stress 2017, 1, 247054701668799. [Google Scholar] [CrossRef]
- Jones, C.; Barrera, I.; Brothers, S.; Ring, R.; Wahlestedt, C. Oxytocin and Social Functioning. Dialogues Clin. Neurosci. 2017, 19, 193–202. [Google Scholar]
- Zoratto, F.; Sbriccoli, M.; Martinelli, A.; Glennon, J.C.; Macrì, S.; Laviola, G. Intranasal oxytocin administration promotes emotional contagion and reduces aggression in a mouse model of callousness. Neuropharmacology 2018, 143, 250–267. [Google Scholar]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and vasopressin in the human brain: Social neuropeptides for translational medicine. Nature reviews. Neuroscience 2011, 12, 524–538. [Google Scholar]
- Striepens, N.; Kendrick, K.M.; Maier, W.; Hurlemann, R. Prosocial effects of oxytocin and clinical evidence for its therapeutic potential. Front. Neuroendocrinol. 2011, 32, 426–450. [Google Scholar]
- Kumsta, R.; Heinrichs, M. Oxytocin, stress and social behavior: Neurogenetics of the human oxytocin system. Curr. Opin. Neurobiol. 2013, 23, 11–16. [Google Scholar] [CrossRef]
- Abu-Akel, A.; Palgi, S.; Klein, E.; Decety, J.; Shamay-Tsoory, S. Oxytocin increases empathy to pain when adopting the other- but not the self-perspective. Soc. Neurosci. 2015, 10, 7–15. [Google Scholar] [CrossRef]
- Laviola, G.; Zoratto, F.; Ingiosi, D.; Carito, V.; Huzard, D.; Fiore, M.; Macrì, S. Low empathy-like behaviour in male mice associates with impaired sociability, emotional memory, physiological stress reactivity and variations in neurobiological regulations. PLoS ONE 2017, 12, e0188907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langford, D.J.; Crager, S.E.; Shehzad, Z.; Smith, S.B.; Sotocinal, S.G.; Levenstadt, J.S.; Chanda, M.L.; Levitin, D.J.; Mogil, J.S. Social modulation of pain as evidence for empathy in mice. Science 2006, 312, 1967–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderschuren, L.J.; Niesink, R.J.; Spruijt, B.M.; Van Ree, J.M. Mu- and kappa-opioid receptor-mediated opioid effects on social play in juvenile rats. Eur. J. Pharmacol. 1995, 276, 257–266. [Google Scholar] [CrossRef]
- Pellissier, L.P.; Gandía, J.; Laboute, T.; Becker, J.A.J.; Le Merrer, J. μ opioid receptor, social behaviour and autism spectrum disorder: Reward matters. Br. J. Pharmacol. 2018, 175, 2750–2769. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, M. The potential influence of maternal stress hormones on development and mental health of the offspring. Brain Behav. Immun. 2005, 19, 296–308. [Google Scholar] [CrossRef]
- Catalani, A.; Marinelli, M.; Scaccianoce, S.; Nicolai, R.; Muscolo, L.A.; Porcu, A.; Korányi, L.; Piazza, P.V.; Angelucci, L. Progeny of mothers drinking corticosterone during lactation has lower stress-induced corticosterone secretion and better cognitive performance. Brain Res. 1993, 624, 209–215. [Google Scholar] [CrossRef]
- Macrì, S.; Pasquali, P.; Bonsignore, L.T.; Pieretti, S.; Cirulli, F.; Chiarotti, F.; Laviola, G. Moderate neonatal stress decreases within-group variation in behavioral, immune and HPA responses in adult mice. PLoS ONE 2007, 2, e1015. [Google Scholar] [CrossRef] [PubMed]
- Maccari, S.; Morley-Fletcher, S. Effects of prenatal restraint stress on the hypothalamus–pituitary–renal axis and related behavioural and neurobiological alterations. Psychoneuroendocrinology 2007, 32, 10–15. [Google Scholar] [CrossRef]
- Daskalakis, N.P.; Enthoven, L.; Schoonheere, E.; de Kloet, E.R.; Oitzl, M.S. Immediate Effects of Maternal Deprivation on the (Re)Activity of the HPA-Axis Differ in CD1 and C57Bl/6J Mouse Pups. Front. Endocrinol. 2014, 5, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Caudal, D.; Jay, T.M.; Godsil, B.P. Behavioral stress induces regionally-distinct shifts of brain mineralocorticoid and glucocorticoid receptor levels. Front. Behav. Neurosci. 2014, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridder, S.; Chourbaji, S.; Hellweg, R.; Urani, A.; Zacher, C.; Schmid, W.; Zink, M.; Hörtnagl, H.; Flor, H.; Henn, F.A.; et al. Mice with genetically altered glucocorticoid receptor expression show altered sensitivity for stress-induced depressive reactions. J. Neurosci. 2005, 25, 6243–6250. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. Glucocorticoid-sensitive hippocampal neurons are involved in terminating the adrenocortical stress response. Proc. Natl. Acad. Sci. USA 1984, 81, 6174–6177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maccari, S.; Piazza, P.V.; Kabbaj, M.; Barbazanges, A.; Simon, H.; Le Moal, M. Adoption reverses the long-term impairment in glucocorticoid feedback induced by prenatal stress. J. Neurosci. 1995, 15, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Koehl, M.; Darnaudéry, M.; Dulluc, J.; Van Reeth, O.; Le Moal, M.; Maccari, S. Prenatal stress alters circadian activity of hypothalamo-pituitary-adrenal axis and hippocampal corticosteroid receptors in adult rats of both gender. J. Neurobiol. 1999, 40, 302–315. [Google Scholar] [CrossRef]
- Kendler, K.S.; Kessler, R.C.; Walters, E.E.; MacLean, C.; Neale, M.C.; Heath, A.C.; Eaves, L.J. Stressful life events, genetic liability and onset of an episode of major depression in women. Am. J. Psychiatry 1995, 152, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Coplan, J.D.; Andrews, M.W.; Rosenblum, L.A.; Owens, M.J.; Friedman, S.; Gorman, J.M.; Nemeroff, C.B. Persistent elevations of cerebrospinal fluid concentrations of corticotrophin releasing factor in adult nonhuman primates exposed to early-life stressors: Implications for the pathophysiology of mood and anxiety disorders. Proc. Natl. Acad. Sci. USA 1996, 93, 1619–1623. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S. The Neurobiology and Neuroendocrinology of Stress. Implications for Post-traumatic Stress Disorder from a Basic Science Perspective. Psychiatr. Clin. N. Am. 2002, 25, 469–494. [Google Scholar] [CrossRef]
- Blair, R.J.; Leibenluft, E.; Pine, D.S. Conduct disorder and callous-unemotional traits in youth. N. Engl. J. Med. 2014, 371, 2207–2216. [Google Scholar] [CrossRef] [PubMed]
- Morley-Fletcher, S.; Rea, M.; Maccari, S.; Laviola, G. Environmental enrichment during adolescence reverses the effects of prenatal stress on play behaviour and HPA axis reactivity in rats. Eur. J. Neurosci. 2003, 18, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Crusio, W.E. Genetic dissection of mouse exploratory behaviour. Behav. Brain Res. 2001, 125, 127–132. [Google Scholar]
- Laviola, G.; Macrì, S.; Morley-Fletcher, S.; Adriani, W. Risk-taking Be-havior in Adolescent Mice: Psychobiological Determinants and Early Epigenetic Influence. Neurosci. Biobehav. Rev. 2003, 27, 19–31. [Google Scholar] [CrossRef]
- Kaiser, S.; Kruijver, F.P.M.; Straub, R.H.; Sachser, N.; Swaab, D.F. Early Social Stress in Male Guinea-pigs Changes Social Behaviour, and Autonomic and Neuroendocrine Functions. J. Neuroendocrinol. 2003, 15, 761–769. [Google Scholar] [CrossRef]
- Sandi, C.; Haller, J. Stress and the Social Brain: Behavioural Effects and Neurobiological Mechanisms. Nat. Rev. Neurosci. 2015, 16, 290–304. [Google Scholar] [CrossRef] [Green Version]
- Panksepp, J.; Jalowiec, J.; DeEskinazi, F.G.; Bishop, P. Opiates and play dominance in juvenile rats. Behav. Neurosci. 1985, 99, 441–453. [Google Scholar]
- Crusio, W.; van Abeelen, J. The genetic architecture of behavioural responses to novelty in mice. Heredity 1986, 56, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Kelley, A.E.; Schochet, T.; Landry, C.F. Risk taking and novelty seeking in adolescence: Introduction to part I. Ann. N.Y. Acad. Sci. 2004, 1021, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Sanci, L.; Webb, M.; Hocking, J. Risk-taking behaviour in adolescents. Aust. J. Gen. Pract. 2018, 47, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoratto, F.; Berry, A.; Anzidei, F.; Fiore, M.; Alleva, E.; Laviola, G.; Macrì, S. Effects of maternal L-tryptophan depletion and corticosterone administration on neurobehavioral adjustments in mouse dams and their adolescent and adult daughters. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1479–1492. [Google Scholar] [CrossRef]
- Lesse, A.; Rether, K.; Gröger, N.; Braun, K.; Bock, J. Chronic Postnatal Stress Induces Depressive-like Behavior in Male Mice and Programs second-Hit Stress-Induced Gene, Expression Patterns of OxtR and AvpR1a in Adulthood. Mol. Neurobiol. 2017, 54, 4813–4819. [Google Scholar] [CrossRef]
- Wigger, D.C.; Gröger, N.; Lesse, A.; Krause, S.; Merz, T.; Gündel, H.; Braun, K.; McCook, O.; Radermacher, P.; Bock, J.; et al. Maternal Separation Induces Long-Term Alterations in the Cardiac Oxytocin Receptor and Cystathionine γ-Lyase Expression in Mice. Oxid. Med. Cell Longev. 2020, 2020, 4309605–4309610. [Google Scholar]
- Maud, C.; Ryan, J.; McIntosh, J.E.; Olsson, C.A. The role of oxytocin receptor gene (OXTR) DNA methylation (DNAm) in human social and emotional functioning: A systematic narrative review. BMC Psychiatry 2018, 18, 154. [Google Scholar] [CrossRef] [Green Version]
- Kalueff, A.V.; Stewart, A.M.; Song, C.; Berridge, K.C.; Graybiel, A.M.; Fentress, J.C. Neurobiology of rodent self-grooming and its value for translational neuroscience. Nat. Rev. Neurosci. 2016, 17, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferkin, M.H.; Leonard, S.T. Self-grooming by rodents in social and sexual contexts. Acta Zool. Sin. 2005, 51, 772–779. [Google Scholar]
- Fernández-Teruel, A.; Estanislau, C. Meanings of self-grooming depend on an inverted U-shaped function with aversiveness. Nat. Rev. Neurosci. 2016, 17, 591. [Google Scholar] [CrossRef]
- Wang, S.C.; Lin, C.C.; Tzeng, N.S.; Tung, C.S.; Liu, Y.P. Effects of oxytocin on prosocial behavior and the associated profiles of oxytocinergic and corticotropin-releasing hormone receptors in a rodent model of posttraumatic stress disorder. J. Biomed. Sci. 2019, 26, 26. [Google Scholar] [CrossRef] [Green Version]
- Estanislau, C.; Veloso, A.W.N.; Filgueiras, G.B.; Maio, T.P.; Dal-Cól, M.L.C.; Cunha, D.C.; Klein, R.; Carmona, L.F.; Fernández-Teruel, A. Rat Self- Grooming and its Relationships with Anxiety, Dearousal and Perseveration: Evidence for a Self-Grooming Trait Physiology of Behaviour. Physiol. Behav. 2019, 209, 1–9. [Google Scholar] [CrossRef]
- Drago, F.; Pedersen, C.A.; Caldwell, J.D.; Prange, A.J. Oxytocin potently enhances novelty-induced grooming behavior in the rat. Brain Res. 1986, 368, 287–295. [Google Scholar] [CrossRef]
- Pedersen, C.A.; Caldwell, J.D.; Drago, F.; Noonan, L.R.; Peterson, G.; Hood, L.E.; Prange, A.J. Grooming Behavioral Effects of Oxytocin. Pharmacology, Ontogeny, and Comparisons with Other Nonapeptides. Ann. N.Y. Acad. Sci. 1988, 525, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Vestlund, J.; Kalafateli, A.L.; Studer, E.; Westberg, L.; Jerlhag, E. Neuromedin U induces self-grooming in socially-stimulated mice. Neuropharmacology 2020, 162, 107818. [Google Scholar] [CrossRef]
- Lu, Y.F.; Ren, B.; Ling, B.F.; Zhang, J.; Xu, C.; Li, Z. Social interaction with a cagemate in pain increases allogrooming and induces pain hypersensitivity in the observer rats. Neurosci. Lett. 2018, 662, 385–388. [Google Scholar]
- Sanathara, N.M.; Garau, C.; Alachkar, A.; Wang, L.; Wang, Z.; Nishimori, K.; Xu, X.; Civelli, O. Melanin concentrating hormone modulates oxytocin-mediated marble burying. Neuropharmacology 2018, 128, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Leckman, J.F.; Goodman, W.K.; North, W.G.; Chappell, P.B.; Price, L.H.; Pauls, D.L.; Anderson, G.M.; Riddle, M.A.; McDougle, C.J.; Barr, L.C. The role of central oxytocin in obsessive compulsive disorder and related normal behavior. Psychoneuroendocrinology 1994, 19, 723–749. [Google Scholar] [CrossRef]
- Hodges, A.; Davis, T.; Crandall, M.; Phipps, L.; Weston, R. Using Shaping to Increase Foods Consumed by Children with Autism. J. Autism Dev. Disord. 2017, 47, 2471–2479. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.S.; Park, J.H. Brain uptake and the analgesic effect of oxytocin--its usefulness as an analgesic agent. Arch. Pharm. Res. 2000, 23, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.; Bai, B.; Liu, W. The analgesic effects of oxytocin in the peripheral and central nervous system. Neurochem. Int. 2017, 103, 57–64. [Google Scholar]
- Cuffe, J.S.; Turton, E.L.; Akison, L.K.; Bielefeldt-Ohmann, H.; Moritz, K.M. Prenatal corticosterone exposure programs sex-specific adrenal adaptations in mouse offspring. J. Endocrinol. 2017, 232, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, G.S.; Sazma, M.A.; McCullough, A.M.; Yonelinas, A.P. The effects of acute stress on episodic memory: A meta-analysis and integrative review. Psychol. Bull. 2017, 143, 636–675. [Google Scholar] [CrossRef] [PubMed]
- Valentino, R.J.; Van Bockstaele, E. Endogenous opioids: The downside of opposing stress. Neurobiol. Stress 2015, 1, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.M.; Fan, K.M.; Qiao, Y.N.; Xu, J.H.; Qiu, L.J.; Li, X.; Liu, Y.; Qian, Z.Q.; Wei, C.L.; Han, J.; et al. Hippocampal µ-opioid receptors on GABAergic neurons mediate stress-induced impairment of memory retrieval. Mol. Psychiatry 2020, 25, 977–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panksepp, J.; Herman, B.; Conner, R.; Bishop, P.; Scott, J.P. The biology of social attachments: Opiates alleviate separation distress. Biol. Psychiatry 1978, 13, 607–618. [Google Scholar]
- Terranova, M.L.; Laviola, G. δ-opioid modulation of social interactions in juvenile mice weaned at different ages. Physiol. Behav. 2001, 73, 393–400. [Google Scholar] [CrossRef]
- Johnson, B.; Ulberg, S.; Shivale, S.; Donaldson, J.; Milczarski, B.; Faraone, S.V. Fibromyalgia, autism, and opioid addiction as natural and induced disorders of the endogenous opioid hormonal system. Discov. Med. 2014, 18, 209–220. [Google Scholar]
- McDonald, S.; Darke, S.; Kaye, S.; Torok, M. Deficits in social perception in opioid maintenance patients, abstinent opioid users and non-opioid users. Addiction 2013, 108, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.C.; Kennedy, S.E.; Smith, Y.R.; Stohler, C.S.; Zubieta, J.K. Interface of physical and emotional stress regulation through the endogenous opioid system and mu-opioid receptors. Prog Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 264–1280. [Google Scholar] [CrossRef] [PubMed]
- Harno, E.; Gali Ramamoorthy, T.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef]
- Yamamoto, M.; Komori, T.; Matsumoto, T.; Zhang, K.; Miyahara, S.; Shizuya, K.; Okazaki, Y. Effects of single and repeated prolonged stress on mu-opioid receptor mRNA expression in rat gross hypothalamic and midbrain homogenates. Brain Res. 2003, 980, 191–196. [Google Scholar] [CrossRef]
- Milner, T.A.; Burstein, S.R.; Marrone, G.F.; Khalid, S.; Gonzalez, A.D.; Williams, T.J.; Schierberl, K.C.; Torres-Reveron, A.; Gonzales, K.L.; McEwen, B.S.; et al. Stress differentially alters mu opioid receptor density and trafficking in parvalbumin-containing interneurons in the female and male rat hippocampus. Synapse 2013, 67, 757–772. [Google Scholar]
- Insel, T.R.; Kinsley, C.H.; Mann, P.E.; Bridges, R.S. Prenatal stress has long-term effects on brain opiate receptors. Brain Res. 1990, 511, 93–97. [Google Scholar] [CrossRef]
- Van’t Veer, A.; Carlezon, W.A. Role of kappa-opioid receptors in stress and anxiety-related behavior. Psychopharmacology 2013, 229, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Butelman, E.R.; McElroy, B.D.; Prisinzano, T.E.; Kreek, M.J. Impact of pharmacological manipulation of the κ-opioid receptor system on self-grooming and Anhedonic-like behaviors in male mice. J. Pharmacol. Exp. Ther. 2019, 370, 1–8. [Google Scholar] [CrossRef]
- Carden, S.E.; Barr, G.; Hofer, M.A. Differential effects of specific opioid receptor agonists on rat pup isolation calls. Dev. Brain Res. 1991, 62, 17–22. [Google Scholar] [CrossRef]
- Mukhin, V.N.; Abdurasulova, I.N.; Pavlov, K.I.; Kozlov, A.P.; Klimenko, V.M. Effects of Activation of κ-Opioid Receptors on Behavior during Postnatal Formation of the Stress Reactivity Systems. Neurosci. Behav. Physiol. 2016, 46, 6. [Google Scholar] [CrossRef]
- Neumann, I.D.; Maloumby, R.; Beiderbeck, D.I.; Lukas, M.; Landgraf, R. Increased brain and plasma oxytocin after nasal and peripheral adminis- tration in rats and mice. Psychoneuroendocrinology 2013, 38, 1985–1993. [Google Scholar] [CrossRef]
- Calcagnoli, F.; Kreutzmann, J.C.; de Boer, S.F.; Althaus, M.; Koolhaas, J.M. Acute and repeated intranasal oxytocin administration exerts anti-aggressive and pro-affiliative effects in male rats. Psychoneuroendocrinology 2015, 51, 112–121. [Google Scholar] [CrossRef]
- Quintana, D.S.; Smerud, K.T.; Andreassen, O.A.; Djupesland, P.G. Evidence for intranasal oxytocin delivery to the brain: Recent advances and future perspectives. Ther. Deliv. 2018, 9, 515–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, B.; Managò, F.; Tucci, V.; Kao, H.T.; Valtorta, F.; Benfenati, F. Autism-related behavioral abnormalities in synapsin knockout mice. Behav. Brain Res. 2013, 251, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Sufka, K.J.; Watson, G.S.; Nothdurft, R.E.; Mogil, J.S. Scoring the mouse formalin test: Validation study. Eur. J. Pain 1998, 2, 351–358. [Google Scholar] [CrossRef]
- Gioiosa, L.; Chen, X.; Watkins, R.; Klanfer, N.; Bryant, C.D.; Evans, C.J.; Arnold, A.P. Sex chromosome complement affects nociception in tests of acute and chronic exposure to morphine in mice. Horm. Behav. 2008, 53, 124–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosland, J.H.; Tjølsen, A.; Mæhle, B.; Hole, K. The formalin test in mice: Effect of formalin concentration. Pain 1990, 42, 235–242. [Google Scholar] [CrossRef]
- Laviola, G.; Terranova, M.L. The developmental psychobiology of behavioural plasticity in mice: The role of social experiences in the family unit. Neurosci. Biobehav. Rev. 1998, 23, 197–213. [Google Scholar] [CrossRef]
- Hartwig, W. Endokrynologia praktyczna (Practical Endocrinology); Państwowy Zakład Wydawnictw Lekarskich: Warszaw, Poland, 1989; pp. 538–576. [Google Scholar]
- Cochran, D.; Fallon, D.; Hill, M.; Frazier, J.A. “The role of oxytocin in psychiatric disorders: A review of biological and therapeutic research findings”. Harv. Rev. Psychiatry 2013, 21, 219–247. [Google Scholar] [CrossRef]
- Bernaerts, S.; Boets, B.; Steyaert, J.; Wenderoth, N.; Alaerts, K. Oxytocin treatment attenuates amygdala activity in autism: A treatment-mechanism study with long-term follow-up. Transl. Psychiatry 2020, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Domes, G.; Ower, N.; von Dawans, B.; Spengler, F.B.; Dziobek, I.; Bohus, M.; Matthies, S.; Philipsen, A.; Heinrichs, M. Effects of intranasal oxytocin administration on empathy and approach motivation in women with borderline personality disorder: A randomized controlled trial. Transl. Psychiatry 2019, 9, 328. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Perinatal Treatment | ||
|---|---|---|---|
| AFR | PreCORT | PostCORT | |
| Response to stress | = | ↓ * | ↓ |
| Sociability (with Social Stimulus) | ↑ | ↑ | ↑ |
| Sociability (with Novel Stimulus) | ↓ | ↓ | ↓ |
| Emotional Contagion (Paw-Licking In-Front) | = | ↑ | = |
| Emotional Contagion (Paw-Licking In-Back) | = | ↑ * | ↑ |
| Self-Grooming | ↑ | ↑ | = |
| Hole Probe Behaviour | ↓ | ↓ | = |
| Formalin Test (Paw-Licking in Non-Social Context) | ↑ | ↑ | ↑ |
| Receptor | Brain Area | Perinatal Treatment | |
|---|---|---|---|
| PreCORT | PostCORT | ||
| GR | Prefrontal Cortex | = | ↓ |
| Hypothalamus | ↓ * | ↓ * | |
| Hippocampus | ↑ * | = | |
| MR | Prefrontal Cortex | ↑ | ↑ |
| Hypothalamus | ↑ * | ↑ | |
| Hippocampus | ↓ * | ↓ | |
| MOR | Prefrontal Cortex | = | ↑ |
| Hypothalamus | ↑ * | ↑ * | |
| Hippocampus | ↓ * | ↓ | |
| KOR | Prefrontal Cortex | ↑ * | = |
| Hypothalamus | ↓ | = | |
| OXTR | Prefrontal Cortex | ↑ | ↑ * |
| Hypothalamus | = | ↑ * | |
| Parameter | Perinatal Treatment | |
|---|---|---|
| PreCORT | PostCORT | |
| Response to stress | ↑ | ↑ |
| Sociability (with Social Stimulus) | ↓ | = |
| Sociability (with Novel Stimulus) | ↑ | ↑ |
| Emotional Contagion (Paw-Licking In-Front) | ↑ | ↑ |
| Emotional Contagion (Paw-Licking In-Back) | ↑ | ↑ |
| Self-Grooming | ↓ | ↑ |
| Hole Probe Behaviour | ↑ | ↓ |
| Formalin Test (Paw-Licking in Non-Social Context) | ↓ | ↓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laviola, G.; Busdraghi, L.M.; Meschino, N.; Petrella, C.; Fiore, M. Aberrant Early in Life Stimulation of the Stress-Response System Affects Emotional Contagion and Oxytocin Regulation in Adult Male Mice. Int. J. Mol. Sci. 2021, 22, 5039. https://doi.org/10.3390/ijms22095039
Laviola G, Busdraghi LM, Meschino N, Petrella C, Fiore M. Aberrant Early in Life Stimulation of the Stress-Response System Affects Emotional Contagion and Oxytocin Regulation in Adult Male Mice. International Journal of Molecular Sciences. 2021; 22(9):5039. https://doi.org/10.3390/ijms22095039
Chicago/Turabian StyleLaviola, Giovanni, Ludovica Maria Busdraghi, Noemi Meschino, Carla Petrella, and Marco Fiore. 2021. "Aberrant Early in Life Stimulation of the Stress-Response System Affects Emotional Contagion and Oxytocin Regulation in Adult Male Mice" International Journal of Molecular Sciences 22, no. 9: 5039. https://doi.org/10.3390/ijms22095039
APA StyleLaviola, G., Busdraghi, L. M., Meschino, N., Petrella, C., & Fiore, M. (2021). Aberrant Early in Life Stimulation of the Stress-Response System Affects Emotional Contagion and Oxytocin Regulation in Adult Male Mice. International Journal of Molecular Sciences, 22(9), 5039. https://doi.org/10.3390/ijms22095039