
Nutraceuticals Synergistically Promote Osteogenesis in Cultured 7F2 Osteoblasts and Mitigate Inhibition of Differentiation and Maturation in Simulated Microgravity



Abstract
:1. Introduction
2. Results
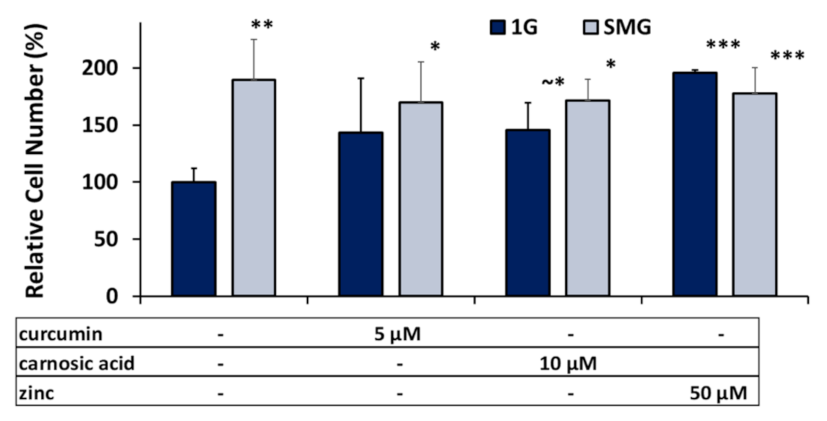
2.1. Nutraceuticals Affect Cell Viability and Increase Cell Numbers:
2.2. Nutraceuticals Modulate Short-Term ALP Activity
2.3. Nutraceuticals Induce Osteogenic Marker Gene Expression in Non-Osteogenic Medium
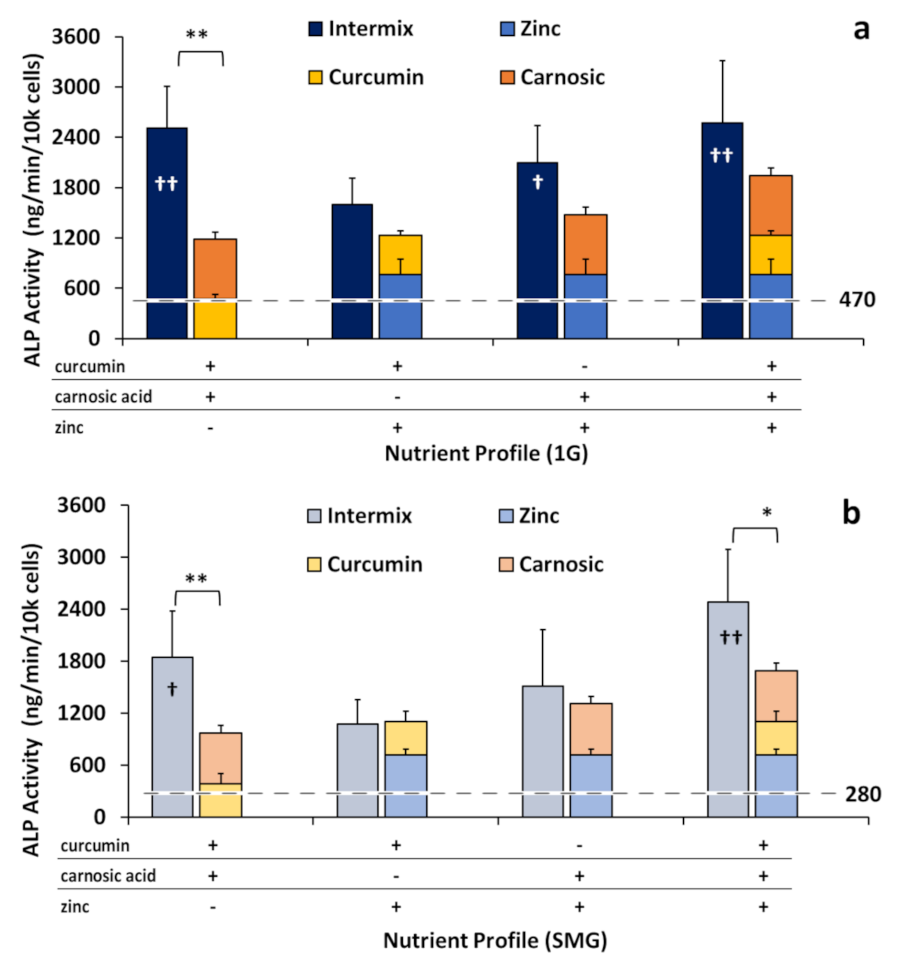
2.4. Phytonutrients Display Synergistic Effects in Intermixtures Independent of Zinc
2.5. Long-Term Application of Nutraceuticals Partly Mitigated Inhibition of ALP Activity and Mineralization by SMG
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture Techniques
4.3. Random Positioning Machine
4.4. Short Term Cytotoxicity/Cell Viability Assay
4.5. PICO Green Assay for Long-Term Cell Proliferation
4.6. Alkaline Phosphatase Activity Assay
4.7. Alizarin Red and Dissolved Calcium TECO Assay for Mineralization
4.8. RNA Extraction and Real-Time PCR for Osteogenic Marker Gene Expression (ALPL, RUN, ON)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase, enzyme |
| ALPl | Alkaline phosphatase, gene |
| BSA | Bovine serum albumin |
| CA | carnosic acid |
| Cur | curcumin |
| DMSO | Dimethyl sulfoxide |
| G | gravity, Earth normal (1G) |
| ON | Osteonectin |
| RPM | Random Positioning Machine, 3D clinostat |
| RUN | Runx2, gene |
| SMG | simulated microgravity (10−3 G) typically via RPM or clinostat |
| Zn | zinc |
| α-MEM | α-Minimal essential medium |
References
- Smith, S.M.; Heer, M.; Shackelford, L.C.; Sibonga, J.D.; Spatz, J.; Pietrzyk, R.A.; Hudson, E.K.; Zwart, S.R. Bone metabolism and renal stone risk during International Space Station missions. Bone 2015, 81, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; LeBlanc, A.; Evans, H.; Lu, Y.; Genant, H.; Yu, A. Cortical and trabecular bone mineral loss from the spine and hip in long-duration spaceflight. J. Bone Miner. Res. 2004, 19, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Zerath, E.; Holy, X.; Roberts, S.G.; Andre, C.; Renault, S.; Hott, M.; Marie, P.J. Spaceflight inhibits bone formation independent of corticosteroid status in growing rats. J. Bone and Miner. Res. 2000, 15, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Stavnichuk, M.; Mikolajewicz, N.; Corlett, T.; Morris, M.; Komarova, S.V. A systematic review and meta-analysis of bone loss in space travelers. NPJ Microgravity 2020, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Orwoll, E.S.; Adler, R.A.; Amin, S.; Binkley, N.; Lewiecki, E.M.; Petak, S.M.; Shapses, S.A.; Sinaki, M.; Watts, N.B.; Sibonga, J.D. Skeletal health in long-duration astronauts: Nature, assessment, and management recommendations from the NASA Bone Summit. J. Bone Miner. Res. 2013, 28, 1243–1255. [Google Scholar] [CrossRef]
- Nelson, E.S.; Jules, K. The microgravity environment for experiments on the International Space Station. J. Gravit. Physiol. 2004, 11, 1–10. [Google Scholar]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. Biomed. Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Ma, X.; Yang, C.; Su, P.; Yin, C.; Qian, A.R. The Impact of Oxidative Stress on the Bone System in Response to the Space Special Environment. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinon, F. Signaling by ROS drives inflammasome activation. Eur. J. Immunol. 2010, 40, 616–619. [Google Scholar] [CrossRef]
- Goldfarb, A.H. Nutritional antioxidants as therapeutic and preventive modalities in exercise-induced muscle damage. Can. J. Appl. Physiol. 1999, 24, 249–266. [Google Scholar] [CrossRef]
- Morabito, C.; Guarnieri, S.; Cucina, A.; Bizzarri, M.; Mariggiò, M.A. Antioxidant Strategy to Prevent Simulated Microgravity-Induced Effects on Bone Osteoblasts. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Chilibeck, P.D. Nutritional interventions during bed rest and spaceflight: Prevention of muscle mass and strength loss, bone resorption, glucose intolerance, and cardiovascular problems. Nutr. Res. 2020, 82, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Mortreux, M.; Riveros, D.; Bouxsein, M.L.; Rutkove, S.B. A Moderate Daily Dose of Resveratrol Mitigates Muscle Deconditioning in a Martian Gravity Analog. Front. Physiol. 2019, 10, 899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, C.C.; Leon, L.L. Biological activities of Curcuma longa L. Mem. Inst. Oswaldo. Cruz. 2001, 96, 723–728. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Misra, J.; Chanda, D.; Kim, D.K.; Li, T.; Koo, S.H.; Back, S.H.; Chiang, J.Y.L.; Choi, H.S. Curcumin differentially regulates endoplasmic reticulum stress through transcriptional corepressor SMILE (small heterodimer partner-interacting leucine zipper protein)-mediated inhibition of CREBH (cAMP responsive element-binding protein H). J. Biol Chem. 2011, 286, 41972–41984. [Google Scholar] [CrossRef] [Green Version]
- Shishodia, S.; Chaturvedi, M.M.; Aggarwal, B.B. Role of curcumin in cancer therapy. Curr. Probl. Cancer 2007, 31, 243–305. [Google Scholar] [CrossRef]
- Ravindran, J.; Prasad, S.; Aggarwal, B.B. Curcumin and cancer cells: How many ways can curry kill tumor cells selectively? AAPS J. 2009, 11, 495–510. [Google Scholar] [CrossRef]
- Son, H.E.; Kim, E.J.; Jang, W.G. Curcumin induces osteoblast differentiation through mild-endoplasmic reticulum stress-mediated such as BMP2 on osteoblast cells. Life Sci. 2018, 193, 34–39. [Google Scholar] [CrossRef]
- Jain, S.; Krishna Meka, S.R.; Chatterjee, K. Curcumin eluting nanofibers augment osteogenesis toward phytochemical based bone tissue engineering. Biomed. Mater. 2016, 11, 055007. [Google Scholar] [CrossRef]
- Birtić, S.; Dussort, P.; Pierre, F.X.; Bily, A.C.; Roller, M. Carnosic acid. Phytochemistry 2015, 115, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Loussouarn, M.; Krieger-Liszkay, A.; Svilar, L.; Bily, A.; Birtić, S.; Havaux, M. Carnosic Acid and Carnosol, Two Major Antioxidants of Rosemary, Act through Different Mechanisms. Plant. Physiol. 2017, 175, 1381–1394. [Google Scholar] [CrossRef] [Green Version]
- Xia, G.; Wang, X.; Sun, H.; Qin, Y.; Fu, M. Carnosic acid (CA) attenuates collagen-induced arthritis in db/db mice via inflammation suppression by regulating ROS-dependent p38 pathway. Free Radic. Biol. Med. 2017, 108, 418–432. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, X.; Zhou, L.; Liu, Z.; Yuan, J.; Cheng, J.; Zhao, J.; Wu, L.; Li, H.; Qiu, H.; et al. Carnosic acid inhibits inflammation response and joint destruction on osteoclasts, fibroblast-like synoviocytes, and collagen-induced arthritis rats. J. Cell Physiol. 2018, 233, 6291–6303. [Google Scholar] [CrossRef]
- Hagiwara, H.; Basnet, R.; Wiyasihati, S.I.; Nakata, K.; Hagiwara, K.; Miyazaki, H.; Yoshida, K. Carnosic acid inhibits the formation of osteoclasts through attenuation of expression of RANKL. PharmaNutrition 2015, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Oner, G.; Bhaumick, B.; Bala, R.M. Effect of zinc deficiency on serum somatomedin levels and skeletal growth in young rats. Endocrinology 1984, 114, 1860–1863. [Google Scholar] [CrossRef]
- Yamaguchi, M. Role of nutritional zinc in the prevention of osteoporosis. Mol. Cell Biochem. 2010, 338, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, M.N.; Akdas, S.; Yazihan, N. Is Zinc an Important Trace Element on Bone-Related Diseases and Complications? A Meta-analysis and Systematic Review from Serum Level, Dietary Intake, and Supplementation Aspects. Biol. Trace Elem. Res. 2021, 199, 535–549. [Google Scholar] [CrossRef]
- Meyers, V.E.; Zayzafoon, M.; Douglas, J.T.; McDonald, J.M. RhoA and cytoskeletal disruption mediate reduced osteoblastogenesis and enhanced adipogenesis of human mesenchymal stem cells in modeled microgravity. J. Bone Miner. Res. 2005, 20, 1858–1866. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Segawa, Y.; Yamaguchi, M. Histomorphological confirmation of the preventive effect of beta-alanyl-L-histidinato zinc on bone loss in ovariectomized rats. Biol. Pharm. Bull. 1994, 17, 862–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Ehara, Y. Zinc decrease and bone metabolism in the femoral-metaphyseal tissues of rats with skeletal unloading. Calcif. Tissue Int. 1995, 57, 218–223. [Google Scholar] [CrossRef]
- Higashi, A.; Nakamura, T.; Nishiyama, S.; Matsukura, M.; Tomoeda, S.; Futagoishi, Y.; Shinohara, M.; Matsuda, I. Zinc kinetics in patients with bone demineralization due to physical immobilization. J. Am. Coll Nutr. 1993, 12, 61–65. [Google Scholar] [CrossRef]
- Patel, M.J.; Chang, K.H.; Sykes, M.C.; Talish, R.; Rubin, C.; Jo, H. Low magnitude and high frequency mechanical loading prevents decreased bone formation responses of 2T3 preosteoblasts. J. Cell Biochem. 2009, 106, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Pardo, S.J.; Patel, M.J.; Sykes, M.C.; Platt, M.O.; Boyd, N.L.; Sorescu, G.P.; Xu, M.; van Loon, J.J.; Wang, M.D.; Jo, H. Simulated microgravity using the Random Positioning Machine inhibits differentiation and alters gene expression profiles of 2T3 preosteoblasts. Am. J. Physiol. Cell Physiol. 2005, 288, C1211–C1221. [Google Scholar] [CrossRef] [Green Version]
- Braveboy-Wagner, J.; Lelkes, P.I. Reduced Gravity Impairs Proliferation, Maturation, and Mineralization of 7F2 Osteoblasts Cultured in a Random Positioning Machine. Nat. Microgravity 2022. Manuscript in review after revision. [Google Scholar]
- Karadas, O.; Mese, G.; Ozcivici, E. Cytotoxic Tolerance of Healthy and Cancerous Bone Cells to Anti-microbial Phenolic Compounds Depend on Culture Conditions. Appl Biochem Biotechnol 2019, 188, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Yang, M.; Guo, B.; Cao, J.; Yang, L.; Guo, X. Zinc upregulates the expression of osteoprotegerin in mouse osteoblasts MC3T3-E1 through PKC/MAPK pathways. Biol Trace Elem. Res. 2012, 146, 340–348. [Google Scholar] [CrossRef]
- Trachtenberg, A.; Muduli, S.; Sidoryk, K.; Cybulski, M.; Danilenko, M. Synergistic Cytotoxicity of Methyl 4-Hydroxycinnamate and Carnosic Acid to Acute Myeloid Leukemia Cells. Front. Pharmacol 2019, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Pesakhov, S.; Khanin, M.; Studzinski, G.P.; Danilenko, M. Distinct combinatorial effects of the plant polyphenols curcumin, carnosic acid, and silibinin on proliferation and apoptosis in acute myeloid leukemia cells. Nutr Cancer 2010, 62, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Pesakhov, S.; Nachliely, M.; Barvish, Z.; Aqaqe, N.; Schwartzman, B.; Voronov, E.; Sharoni, Y.; Studzinski, G.P.; Fishman, D.; Danilenko, M. Cancer-selective cytotoxic Ca2+ overload in acute myeloid leukemia cells and attenuation of disease progression in mice by synergistically acting polyphenols curcumin and carnosic acid. Oncotarget 2016, 7, 31847–31861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buravkova, L.B.; Gershovich, P.M.; Gershovich, J.G.; Grigor’ev, A.I. Mechanisms of gravitational sensitivity of osteogenic precursor cells. Acta Naturae 2010, 2, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, Y.; Suzuki, K.; Koshikawa, M.; Ando, M.; Iida, J. Necessity of enzymatic activity of alkaline phosphatase for mineralization of osteoblastic cells. Jpn J. Pharmacol 2002, 88, 262–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, F.U.; Rehman, M.S.; Khan, M.S.; Ali, M.A.; Javed, A.; Nawaz, A.; Yang, C. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, S.; Scheyer, T.; Romano, C.S.; Vojnov, A.A. Antioxidant and antimicrobial activities of rosemary extracts linked to their polyphenol composition. Free Radic Res. 2006, 40, 223–231. [Google Scholar] [CrossRef]
- Satoh, T.; McKercher, S.R.; Lipton, S.A. Reprint of: Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic Biol Med. 2014, 66, 45–57. [Google Scholar] [CrossRef]
- de Oliveira, M.R. The Dietary Components Carnosic Acid and Carnosol as Neuroprotective Agents: A Mechanistic View. Mol. Neurobiol. 2016, 53, 6155–6168. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.R. Carnosic Acid as a Promising Agent in Protecting Mitochondria of Brain Cells. Mol. Neurobiol 2018, 55, 6687–6699. [Google Scholar] [CrossRef]
- Chen, D.; Waite, L.C.; Pierce, W.M. In vitro effects of zinc on markers of bone formation. Biol Trace Elem Res 1999, 68, 225–234. [Google Scholar] [CrossRef]
- Montazerolghaem, M.; Ning, Y.; Engqvist, H.; Karlsson Ott, M.; Tenje, M.; Mestres, G. Simvastatin and zinc synergistically enhance osteoblasts activity and decrease the acute response of inflammatory cells. J. Mater. Sci Mater. Med. 2016, 27, 23. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Oishi, H.; Suketa, Y. Stimulatory effect of zinc on bone formation in tissue culture. Biochem Pharmacol 1987, 36, 4007–4012. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Matsui, T. Stimulatory effect of zinc-chelating dipeptide on deoxyribonucleic acid synthesis in osteoblastic MC3T3-E1 cells. Peptides 1996, 17, 1207–1211. [Google Scholar] [CrossRef]
- Lutz, W.; Burritt, M.F.; Nixon, D.E.; Kao, P.C.; Kumar, R. Zinc increases the activity of vitamin D-dependent promoters in osteoblasts. Biochem Biophys Res. Commun 2000, 271, 1–7. [Google Scholar] [CrossRef]
- Alcantara, E.H.; Lomeda, R.A.; Feldmann, J.; Nixon, G.F.; Beattie, J.H.; Kwun, I.S. Zinc deprivation inhibits extracellular matrix calcification through decreased synthesis of matrix proteins in osteoblasts. Mol. Nutr Food Res. 2011, 55, 1552–1560. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Ohtaki, J. Effect of beta-alanyl-L-histidinato zinc on osteoblastic MC3T3-E1 cells: Increases in alkaline phosphatase and proliferation. Pharmacology 1991, 43, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Ozaki, K.; Hoshi, T. Beta-alanyl-L-histidinato zinc prevents skeletal unloading-induced disorder of bone metabolism in rats. Res. Exp. Med. (Berl) 1990, 190, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Notoya, M.; Nishimura, H.; Woo, J.T.; Nagai, K.; Ishihara, Y.; Hagiwara, H. Curcumin inhibits the proliferation and mineralization of cultured osteoblasts. Eur J. Pharmacol 2006, 534, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Braveboy-Wagner, J.; Lelkes, P.I. Altered cell signaling, nuclear morphology and cytoskeletal assembly of 7F2 osteoblasts cultured in simulated microgravity in vitro. 2022; Manuscript in preparation. [Google Scholar]
- Anto, R.J.; Mukhopadhyay, A.; Denning, K.; Aggarwal, B.B. Curcumin (diferuloylmethane) induces apoptosis through activation of caspase-8, BID cleavage and cytochrome c release: Its suppression by ectopic expression of Bcl-2 and Bcl-xl. Carcinogenesis 2002, 23, 143–150. [Google Scholar] [CrossRef]
- Bush, J.A.; Cheung, K.J.; Li, G. Curcumin induces apoptosis in human melanoma cells through a Fas receptor/caspase-8 pathway independent of p53. Exp. Cell Res. 2001, 271, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, S.; Yamaguchi, M. Genistein and zinc synergistically enhance gene expression and mineralization in osteoblastic MC3T3-E1 cells. Int J. Mol. Med. 2007, 19, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Gaus, S.; Li, H.; Li, S.; Wang, Q.; Kottek, T.; Hahnel, S.; Liu, X.; Deng, Y.; Ziebolz, D.; Haak, R.; et al. Shared Genetic and Epigenetic Mechanisms between the Osteogenic Differentiation of Dental Pulp Stem Cells and Bone Marrow Stem Cells. Biomed. Res. Int 2021, 2021, 6697810. [Google Scholar] [CrossRef]
- Fernández, J.M.; Molinuevo, M.S.; McCarthy, A.D.; Cortizo, A.M. Strontium ranelate stimulates the activity of bone-specific alkaline phosphatase: Interaction with Zn(2+) and Mg (2+). Biometals 2014, 27, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Benavides Damm, T.; Walther, I.; Wüest, S.L.; Sekler, J.; Egli, M. Cell cultivation under different gravitational loads using a novel random positioning incubator. Biotechnol Bioeng 2014, 111, 1180–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, A.; Herranz, R.; den Toom, L.A.; Te Slaa, S.; Borst, G.; Visser, M.; Medina, F.J.; van Loon, J.J.W.A. Novel, Moon and Mars, partial gravity simulation paradigms and their effects on the balance between cell growth and cell proliferation during early plant development. NPJ Microgravity 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Kamal, K.Y.; Herranz, R.; van Loon, J.J.W.A.; Medina, F.J. Simulated microgravity, Mars gravity, and 2g hypergravity affect cell cycle regulation, ribosome biogenesis, and epigenetics in Arabidopsis cell cultures. Sci. Rep. 2018, 8, 6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.Y.; Lin, Y.J. In vitro effects of low frequency electromagnetic fields on osteoblast proliferation and maturation in an inflammatory environment. Bioelectromagnetics 2011, 32, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Montoya, C.; Orrego, S.; Wei, X.; Ling, J.; Lelkes, P.I.; Yang, M. Topographic cues of a novel bilayered scaffold modulate dental pulp stem cells differentiation by regulating YAP signalling through cytoskeleton adjustments. Cell Prolif 2019, 52, e12676. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Curcumin | Carnosic Acid | Zinc | |||
|---|---|---|---|---|---|
| 2.5 µM | 89.7 ± 2.4 * | 5 µM | 85.1 ± 1.2 | 50 µM | 88.2 ± 3.5 |
| 5 µM | 71.6 ± 2.5 *** | 10 µM | 82.3 ± 2.1 | 100 µM | 79.7 ± 2.5 * |
| 7.5 µM | 65.3 ± 4.5 *** | 25 µM | 65.8 ± 0.8 * | 250 µM | 71.7 ± 8.3 * |
| 15 µM | 42.9 ± 1.5 *** | 50 µM | 48.9 ± 13.2 ** | 500 µM | 67.3 ± 9.3 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braveboy-Wagner, J.; Sharoni, Y.; Lelkes, P.I. Nutraceuticals Synergistically Promote Osteogenesis in Cultured 7F2 Osteoblasts and Mitigate Inhibition of Differentiation and Maturation in Simulated Microgravity. Int. J. Mol. Sci. 2022, 23, 136. https://doi.org/10.3390/ijms23010136
Braveboy-Wagner J, Sharoni Y, Lelkes PI. Nutraceuticals Synergistically Promote Osteogenesis in Cultured 7F2 Osteoblasts and Mitigate Inhibition of Differentiation and Maturation in Simulated Microgravity. International Journal of Molecular Sciences. 2022; 23(1):136. https://doi.org/10.3390/ijms23010136
Chicago/Turabian StyleBraveboy-Wagner, Justin, Yoav Sharoni, and Peter I. Lelkes. 2022. "Nutraceuticals Synergistically Promote Osteogenesis in Cultured 7F2 Osteoblasts and Mitigate Inhibition of Differentiation and Maturation in Simulated Microgravity" International Journal of Molecular Sciences 23, no. 1: 136. https://doi.org/10.3390/ijms23010136
APA StyleBraveboy-Wagner, J., Sharoni, Y., & Lelkes, P. I. (2022). Nutraceuticals Synergistically Promote Osteogenesis in Cultured 7F2 Osteoblasts and Mitigate Inhibition of Differentiation and Maturation in Simulated Microgravity. International Journal of Molecular Sciences, 23(1), 136. https://doi.org/10.3390/ijms23010136