
The Protective Effects of Lactoferrin on Aflatoxin M1-Induced Compromised Intestinal Integrity



Abstract
:1. Introduction
2. Results
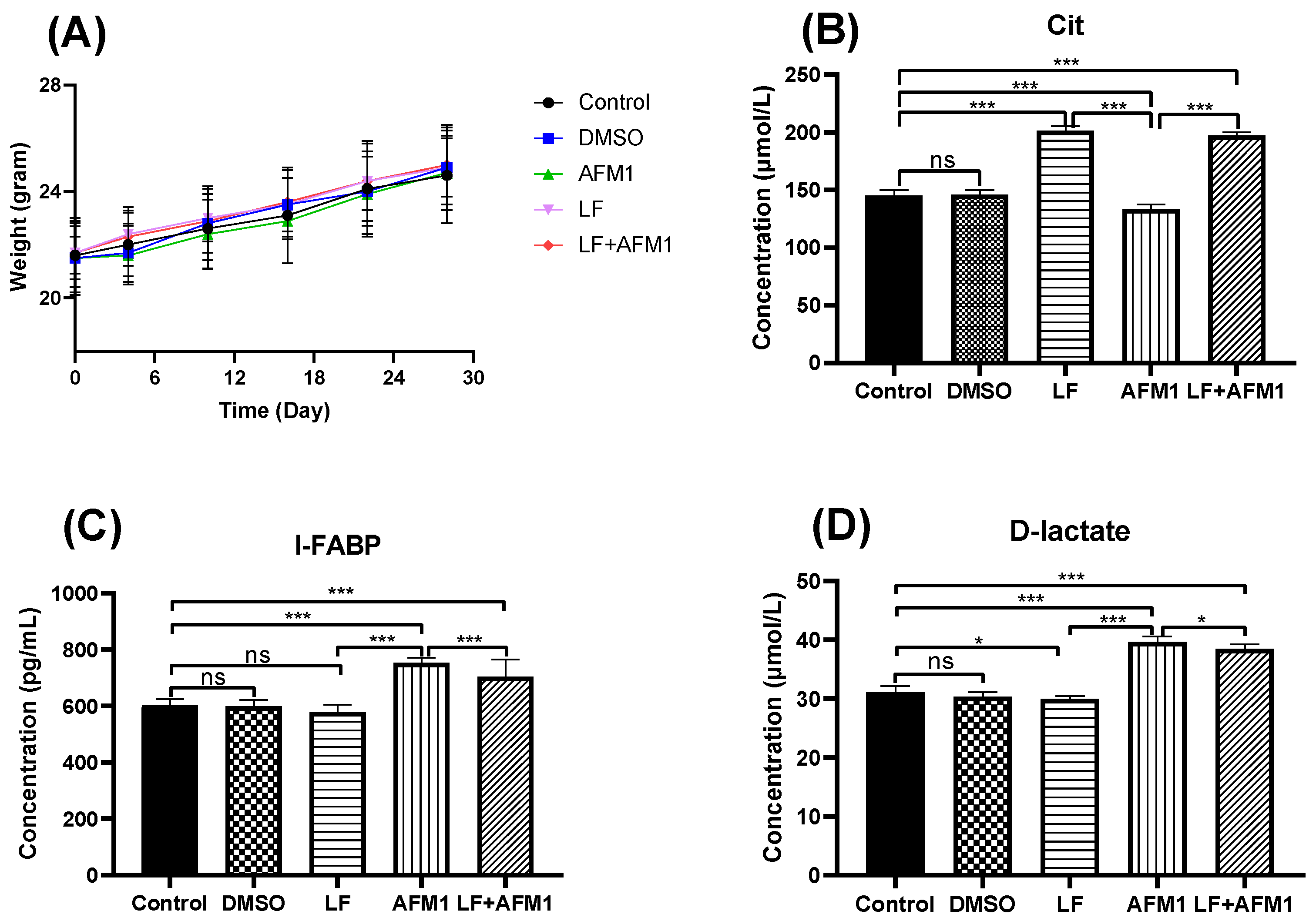
2.1. LF Inhibited AFM1-Induced Compromised Intestinal Barrier In Vivo
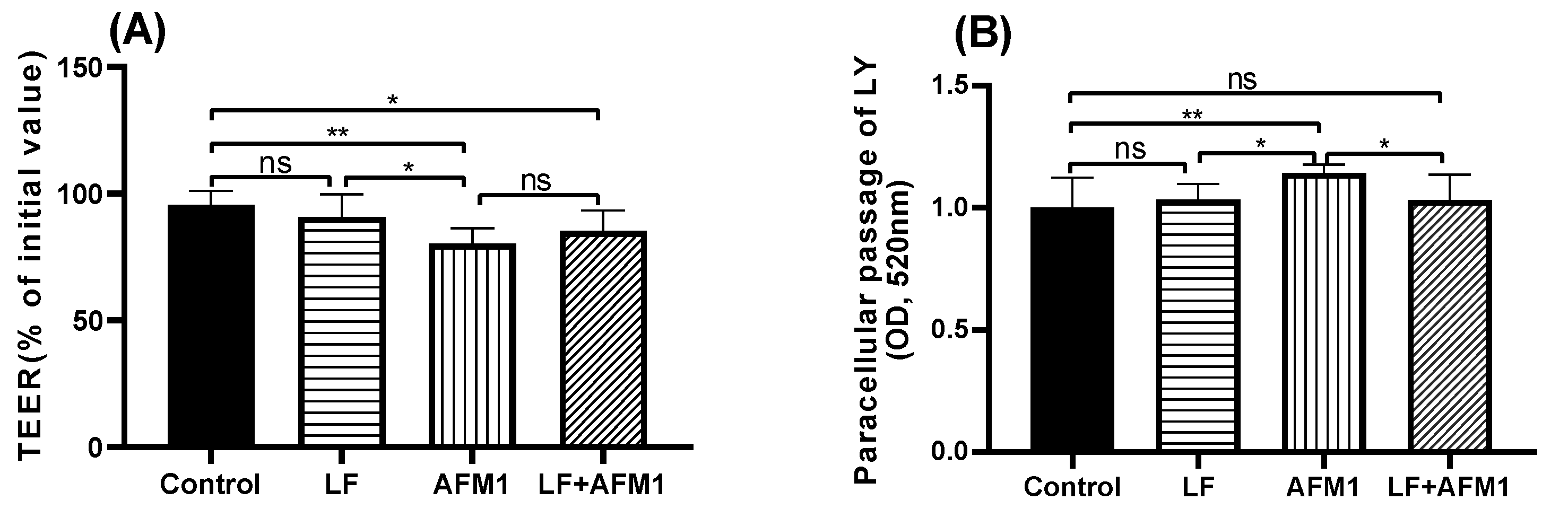
2.2. Effect of LF on Injured Differentiated Caco-2 Cells Induced by AFM1
2.3. Effect of LF on the Gene Expression in AFM1 Treatment In Vitro
2.4. Effect of LF on the Protein Expression in AFM1 Treatment In Vitro
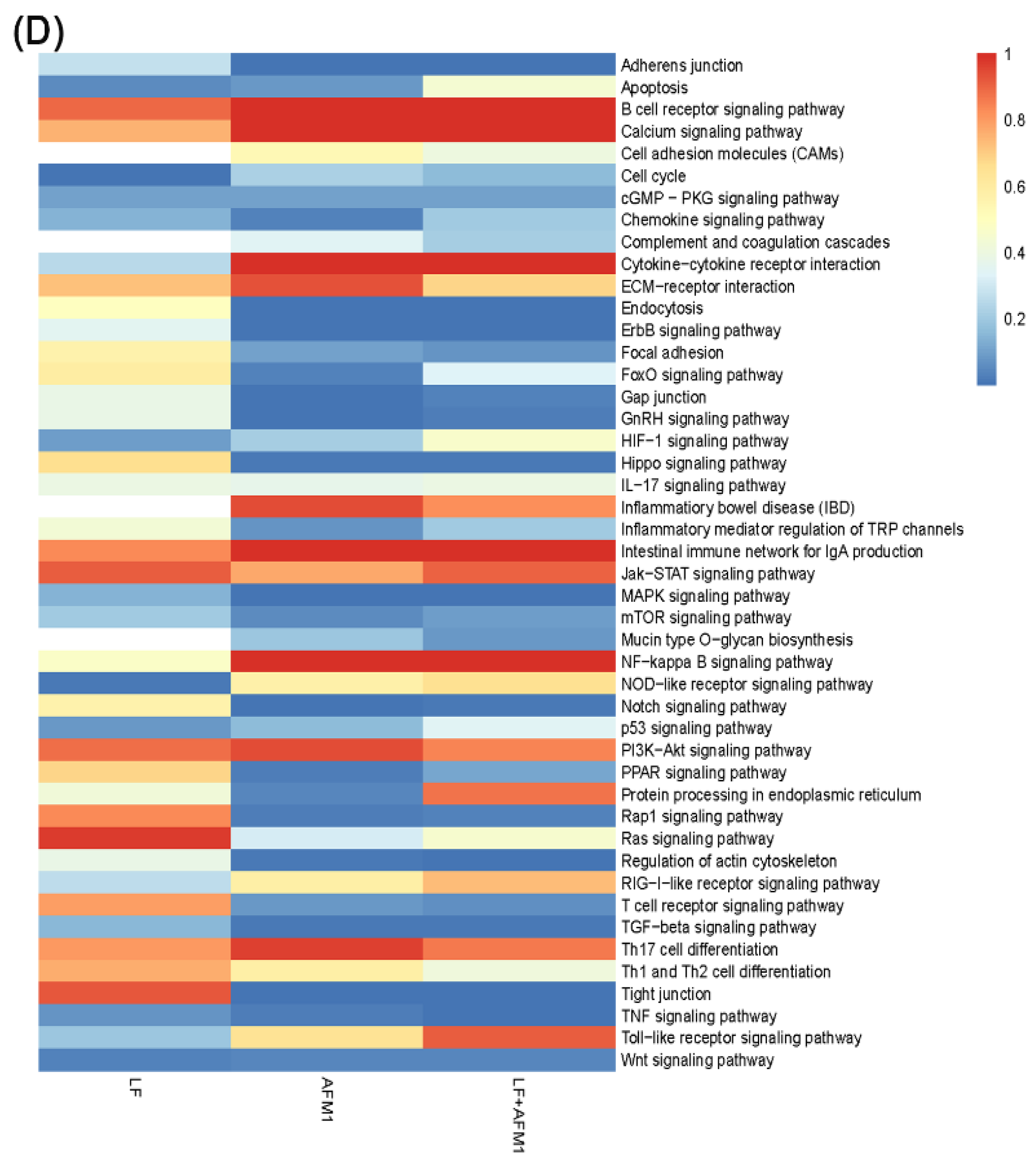
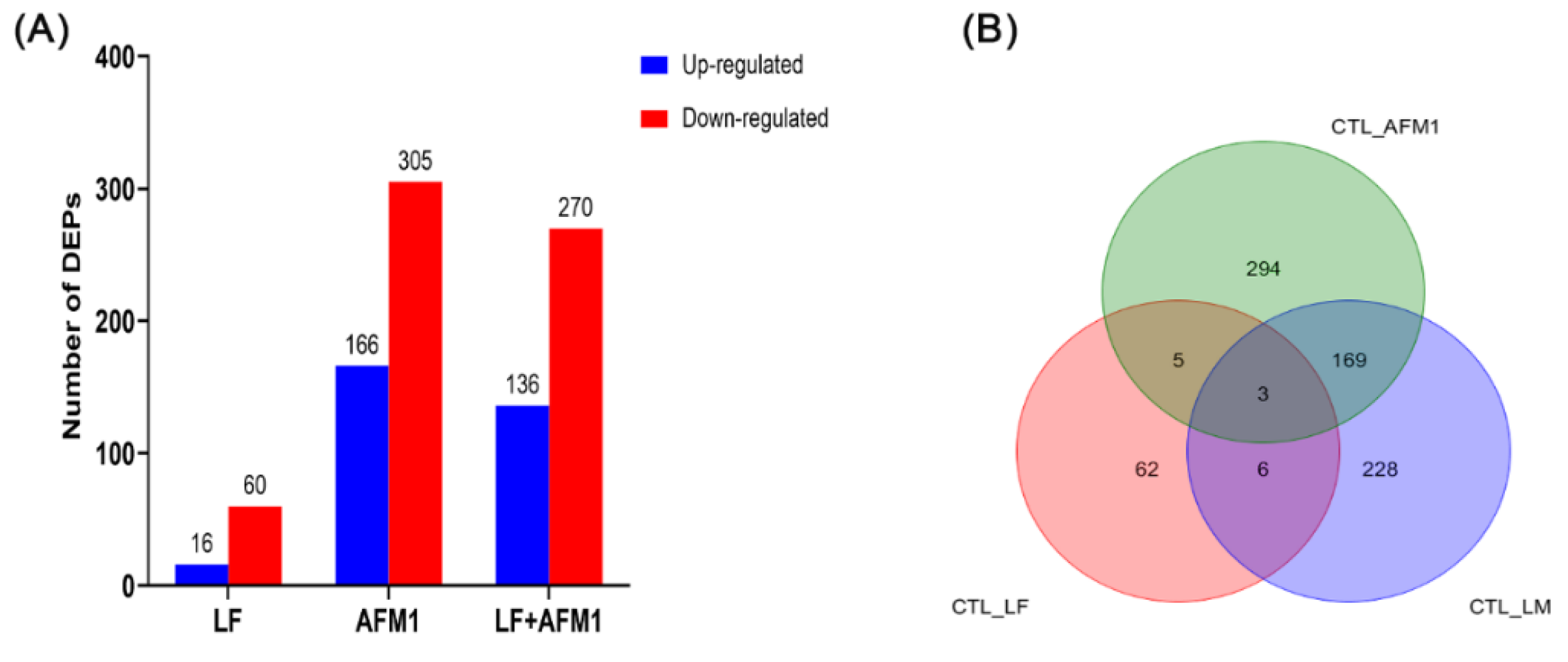
2.5. Omics-Analysis of the Transcriptome and Proteome
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Serum Biochemical Indicators Determination
4.4. Histological Assessment of the Ileum in Mice
4.5. Immunofluorescence Analysis of the Ileum in Mice
4.6. Cells Culture, TEER Values and Permeability Measurement
4.7. Transcriptomics Studies
4.8. Data-Independent Acquisition-Based (DIA) Proteomics Studies
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nongonierma, A.B.; FitzGerald, R.J. The scientific evidence for the role of milk protein-derived bioactive peptides in humans: A Review. J. Funct. Foods 2015, 17, 640–656. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.Z.; Jinap, S.; Pirouz, A.A.; Ahmad Faizal, A.R. Aflatoxin M1 in milk and dairy products, occurrence and recent challenges: A review. Trends Food Sci. Technol. 2015, 46, 110–119. [Google Scholar] [CrossRef]
- Zulueta, A.; Maurizi, A.; Frígola, A.; Esteve, M.J.; Coli, R.; Burini, G. Antioxidant capacity of cow milk, whey and deproteinized milk. Int. Dairy J. 2009, 19, 380–385. [Google Scholar] [CrossRef]
- Min, L.; Li, D.; Tong, X.; Sun, H.; Chen, W.; Wang, G.; Zheng, N.; Wang, J. The challenges of global occurrence of aflatoxin M1 contamination and the reduction of aflatoxin M1 in milk over the past decade. Food Control. 2020, 117, 107352. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Gao, Y.; Li, S.; Bao, X.; Luo, C.; Yang, H.; Wang, J.; Zhao, S.; Zheng, N. Transcriptional and Proteomic Analysis Revealed a Synergistic Effect of Aflatoxin M1 and Ochratoxin A Mycotoxins on the Intestinal Epithelial Integrity of Differentiated Human Caco-2 Cells. J. Proteome Res. 2018, 17, 3128–3142. [Google Scholar] [CrossRef]
- Gao, Y.; Li, S.; Wang, J.; Luo, C.; Zhao, S.; Zheng, N. Modulation of Intestinal Epithelial Permeability in Differentiated Caco-2 Cells Exposed to Aflatoxin M1 and Ochratoxin A Individually or Collectively. Toxins 2017, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Ye, Q.; Bao, X.; Huang, X.; Wang, J.; Zheng, N. Transcriptomic and proteomic profiling reveals the intestinal immunotoxicity induced by aflatoxin M1 and ochratoxin A. Toxicon 2020, 180, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, Y.; Li, S.; Wu, C.; Wang, J.; Zheng, N. Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins 2019, 11, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Gao, Y.; Li, S.; Huang, X.; Bao, X.; Wang, J.; Zheng, N. Modulation of intestinal epithelial permeability and mucin mRNA (MUC2, MUC5AC, and MUC5B) expression and protein secretion in Caco-2/HT29-MTX co-cultures exposed to aflatoxin M1, ochratoxin A, and zearalenone individually or collectively. Toxicol. Lett. 2019, 309, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Cutone, A.; Lepanto, M.S.; Paesano, R.; Valenti, P. Lactoferrin: A Natural Glycoprotein Involved in Iron and Inflammatory Homeostasis. Int. J. Mol. Sci. 2017, 18, 1985. [Google Scholar] [CrossRef]
- Garcia-Montoya, I.A.; Cendon, T.S.; Arevalo-Gallegos, S.; Rascon-Cruz, Q. Lactoferrin a multiple bioactive protein: An overview. Biochim. Biophys. Acta 2012, 1820, 226–236. [Google Scholar] [CrossRef]
- Chen, K.; Chai, L.; Li, H.; Zhang, Y.; Xie, H.M.; Shang, J.; Tian, W.; Yang, P.; Jiang, A.C. Effect of bovine lactoferrin from iron-fortified formulas on diarrhea and respiratory tract infections of weaned infants in a randomized controlled trial. Nutrition 2016, 32, 222–227. [Google Scholar] [CrossRef]
- Comstock, S.S.; Reznikov, E.A.; Contractor, N.; Donovan, S.M. Dietary bovine lactoferrin alters mucosal and systemic immune cell responses in neonatal piglets. J. Nutr. 2014, 144, 525–532. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, X.X.; Liu, Y.; Xi, E.Z.; An, J.J.; Tabys, D.; Liu, N. The In Vitro Protective Role of Bovine Lactoferrin on Intestinal Epithelial Barrier. Molecules 2019, 24, 148. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Gunasekaran, S.; Saleh, D.M.; Alexander, W.T.; Alexander, D.B.; Ohara, H.; Tsuda, H. Effects of oral bovine lactoferrin on a mouse model of inflammation associated colon cancer. Biochem. Cell Biol. 2021, 99, 159–165. [Google Scholar] [CrossRef]
- Alexander, D.B.; Iigo, M.; Abdelgied, M.; Ozeki, K.; Tanida, S.; Joh, T.; Takahashi, S.; Tsuda, H. Bovine lactoferrin and Crohn’s disease: A case study. Biochem. Cell Biol. 2017, 95, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, D.R.; Newburg, D.S. Clinical applications of bioactive milk components. Nutr. Rev. 2015, 73, 463–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertuccini, L.; Costanzo, M.; Iosi, F.; Tinari, A.; Terruzzi, F.; Stronati, L.; Aloi, M.; Cucchiara, S.; Superti, F. Lactoferrin prevents invasion and inflammatory response following E. coli strain LF82 infection in experimental model of Crohn’s disease. Digest. Liver. Dis. 2014, 46, 496–504. [Google Scholar] [CrossRef]
- Zheng, N.; Zhang, H.; Li, S.L.; Wang, J.Q.; Liu, J.; Ren, H.; Gao, Y.N. Lactoferrin inhibits aflatoxin B1- and aflatoxin M1-induced cytotoxicity and DNA damage in Caco-2, HEK, Hep-G2, and SK-N-SH cells. Toxicon 2018, 150, 77–85. [Google Scholar] [CrossRef]
- Drolia, R.; Bhunia, A.K. Crossing the Intestinal Barrier via Listeria Adhesion Protein and Internalin A. Trends Microbiol. 2019, 27, 408–425. [Google Scholar] [CrossRef] [PubMed]
- Lonnerdal, B. Infant formula and infant nutrition: Bioactive proteins of human milk and implications for composition of infant formulas. Am. J. Clin. Nutr. 2014, 99, 712S–717S. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Samadi, N.; Klems, M.; Untersmayr, E. The role of gastrointestinal permeability in food allergy. Ann. Allergy Asthma. Immunol. 2018, 121, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Arnott, I.D.; Kingstone, K.; Ghosh, S. Abnormal Intestinal Permeability Predicts Relapse in Inactive Crohn Disease. Scandinavian J. Gastroenterol. 2000, 35, 1163–1169. [Google Scholar] [CrossRef]
- Gu, A.; Yang, L.; Wang, J.; Li, J.; Shan, A. Protective effect of glutamine and alanyl-glutamine against zearalenone-induced intestinal epithelial barrier dysfunction in IPEC-J2 cells. Res. Vet. Sci. 2021, 137, 48–55. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Dong, R.; Zhang, Z.; Jia, F.; Yu, H.; Wang, Y.; Zhang, Z. Protective effect of selenomethionine on intestinal injury induced by T- 2 toxin. Res. Vet. Sci. 2020, 132, 439–447. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Zhang, G. A proteomic study on the protective effect of kaempferol pretreatment against deoxynivalenol-induced intestinal barrier dysfunction in a Caco-2 cell model. Food Funct. 2020, 11, 7266–7279. [Google Scholar] [CrossRef]
- Xiao, K.; Liu, C.; Qin, Q.; Zhang, Y.; Wang, X.; Zhang, J.; Odle, J.; Lin, X.; Hu, C.A.; Liu, Y. EPA and DHA attenuate deoxynivalenol-induced intestinal porcine epithelial cell injury and protect barrier function integrity by inhibiting necroptosis signaling pathway. FASEB J. 2020, 34, 2483–2496. [Google Scholar] [CrossRef] [PubMed]
- Crenn, P.; Messing, B.; Cynober, L. Citrulline as a biomarker of intestinal failure due to enterocyte mass reduction. Clin. Nutr. 2008, 27, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Gajda, A.M.; Storch, J. Enterocyte fatty acid-binding proteins (FABPs): Different functions of liver and intestinal FABPs in the intestine. Prostaglandins Leukot. Essent. Fat. Acids 2015, 93, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Eng, R.H.; Campos, J.M.; Chmel, H. D-lactic acid measurements in the diagnosis of bacterial infections. J. Clin. Microbiol. 1989, 27, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Naehrer, K.; Applegate, T.J. Interactive effects of dietary protein concentration and aflatoxin B1 on performance, nutrient digestibility, and gut health in broiler chicks. Poult. Sci. 2016, 95, 1312–1325. [Google Scholar] [CrossRef]
- Haiwen, Z.; Rui, H.; Bingxi, Z.; Qingfeng, G.; Jifeng, Z.; Xuemei, W.; Beibei, W. Oral Administration of Bovine Lactoferrin-Derived Lactoferricin (Lfcin) B Could Attenuate Enterohemorrhagic Escherichia coli O157:H7 Induced Intestinal Disease through Improving Intestinal Barrier Function and Microbiota. J. Agric. Food Chem. 2019, 67, 3932–3945. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhao, F.; Zhu, W.; Wang, J. Effects of early-life lactoferrin intervention on growth performance, small intestinal function and gut microbiota in suckling piglets. Food Funct. 2019, 10, 5361–5373. [Google Scholar] [CrossRef]
- Wei, Y.L.; Xu, J.Y.; Zhang, R.; Zhang, Z.; Zhao, L.; Qin, L.Q. Effects of lactoferrin on X-ray-induced intestinal injury in Balb/C mice. Appl. Radiat. Isot. 2019, 146, 72–77. [Google Scholar] [CrossRef]
- Hering, N.A.; Luettig, J.; Krug, S.M.; Wiegand, S.; Gross, G.; van Tol, E.A.; Schulzke, J.D.; Rosenthal, R. Lactoferrin protects against intestinal inflammation and bacteria-induced barrier dysfunction in vitro. Ann. N. Y. Acad. Sci. 2017, 1405, 177–188. [Google Scholar] [CrossRef]
- Liu, J.; Li, B.; Lee, C.; Zhu, H.; Zheng, S.; Pierro, A. Protective effects of lactoferrin on injured intestinal epithelial cells. J. Pediatr. Surg. 2019, 54, 2509–2513. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yang, M.; Guo, J.; Feng, Z. Effects of Bovine Lactoferrin on Rat Intestinal Epithelial Cells. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 645–651. [Google Scholar] [CrossRef]
- Liu, L.; Jiang, R.; Liu, J.; Lonnerdal, B. The bovine Lactoferrin-Osteopontin complex increases proliferation of human intestinal epithelial cells by activating the PI3K/Akt signaling pathway. Food Chem. 2020, 310, 125919. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Bao, X.; Meng, L.; Liu, H.; Wang, J.; Zheng, N. Aflatoxin B1 and Aflatoxin M1 Induce Compromised Intestinal Integrity through Clathrin-Mediated Endocytosis. Toxins 2021, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Bertiaux-Vandaele, N.; Youmba, S.B.; Belmonte, L.; Lecleire, S.; Antonietti, M.; Gourcerol, G.; Leroi, A.M.; Dechelotte, P.; Menard, J.F.; Ducrotte, P.; et al. The expression and the cellular distribution of the tight junction proteins are altered in irritable bowel syndrome patients with differences according to the disease subtype. Am. J. Gastroenterol. 2011, 106, 2165–2173. [Google Scholar] [CrossRef]
- Hering, N.A.; Fromm, M.; Schulzke, J.D. Determinants of colonic barrier function in inflammatory bowel disease and potential therapeutics. J. Physiol. 2012, 590, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Brandt, A.; Nier, A.; Jin, C.J.; Baumann, A.; Jung, F.; Ribas, V.; Garcia-Ruiz, C.; Fernandez-Checa, J.C.; Bergheim, I. Consumption of decaffeinated coffee protects against the development of early non-alcoholic steatohepatitis: Role of intestinal barrier function. Redox Biol. 2019, 21, 101092. [Google Scholar] [CrossRef]
- Qin, T.; Yang, J.; Huang, D.; Zhang, Z.; Huang, Y.; Chen, H.; Xu, G. DOCK4 stimulates MUC2 production through its effect on goblet cell differentiation. J. Cell Physiol. 2021, 236, 6507–6519. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Symbol | Log2FC (AFM1/Control) | Log2FC (LM/Control) | p Value (AFM1/Control) | p Value (LM/Control) | KEGG Pathway | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Protein | mRNA | Protein | mRNA | Protein | mRNA | Protein | mRNA | |||
| clusterin | CLU | 0.67 | 1.32 | 0.77 | 1.14 | 0.0001 | 8.68 × 10−53 | 8.13 × 10−5 | 3.47 × 10−33 | Complement and coagulation cascades |
| G protein subunit alpha i1 | GNAI1 | 1.19 | 0.85 | 1.65 | 0.99 | 0.97 | 0.0001 | 0.65 | 0.0003 | Rap1 signaling pathway, Chemokine signaling pathway |
| agrin | AGRN | −1.15 | −0.88 | −0.96 | −0.82 | 1.03 × 10−12 | 9.12 × 10−14 | 9.06 × 10−8 | 7.15 × 10−13 | ECM-receptor interaction |
| ribonucleotide reductase regulatory subunit M2 | RRM2 | −1.53 | −0.99 | −1.81 | −1.39 | 8.86 × 10−10 | 0.01 | 5.59 × 10−7 | 0.0002 | p53 signaling pathway |
| hepatocyte nuclear factor 4 alpha | HNF4A | −1.09 | −1.57 | −1.11 | −1.44 | 1.27 × 10−9 | 7.36 × 10−37 | 2.98 × 10−8 | 1.45 × 10−19 | AMPK signaling pathway |
| PVR cell adhesion molecule | PVR | −0.88 | −0.91 | −0.71 | −0.96 | 3.14 × 10−5 | 9.28 × 10−12 | 0.001 | 3.77 × 10−14 | Cell adhesion molecules (CAMs) |
| JunD proto-oncogene | JUND | −1.74 | −1.71 | −0.77 | −1.12 | 0.18 | 1.34 × 10−43 | 0.017 | 5.00 × 10−21 | MAPK signaling pathway, IL-17 signaling pathway |
| phosphoenolpyruvate carboxykinase 1 | PCK1 | −1.94 | −3.14 | −0.84 | −0.68 | 0.003 | 1.05 × 10−16 | 0.02 | 2.67 × 10−37 | FoxO signaling pathway, PI3K-Akt signaling pathway |
| erb-b2 receptor tyrosine kinase 3 | ERBB3 | −1.59 | −0.90 | −0.83 | −0.68 | 0.007 | 1.74 × 10−14 | 0.03 | 1.50 × 10−9 | ErbB signaling pathway, Calcium signaling pathway |
| insulin receptor | INSR | −1.29 | −1.69 | −1.23 | −1.15 | 0.10 | 1.76 × 10−42 | 0.07 | 1.78 × 10−18 | Adherens junction, HIF-1 signaling pathway |
| Amphiregulin | AREG | −1.08 | −1.26 | −1.89 | −1.02 | 0.08 | 9.20 × 10−18 | 0.13 | 6.49 × 10−12 | PI3K-Akt signaling pathway, MAPK signaling pathway |
| exocyst complex component 2 | EXOC2 | −0.85 | −1.38 | −1.19 | −1.11 | 0.37 | 2.72 × 10−23 | 0.15 | 5.13 × 10−14 | Ras signaling pathway |
| E1A binding protein p300 | EP300 | −1.11 | −0.97 | −0.93 | −0.76 | 0.25 | 7.10 × 10−11 | 0.33 | 1.55 × 10−6 | HIF-1 signaling pathway, Wnt signaling pathway |
| cytoplasmic FMR1 interacting protein 2 | CYFIP2 | −1.41 | −1.64 | −1.16 | −1.31 | 0.02 | 4.49 × 10−22 | 0.39 | 9.66 × 10−14 | Regulation of actin cytoskeleton |
| nuclear receptor corepressor 2 | NCOR2 | −1.31 | −1.78 | −2.94 | −1.46 | 0.41 | 4.33 × 10−33 | 0.61 | 4.30 × 10−18 | Notch signaling pathway |
| dedicator of cytokinesis 1 | DOCK1 | −1.28 | −2.01 | −1.19 | −2.12 | 0.16 | 1.98 × 10−41 | 0.92 | 7.24 × 10−61 | Focal adhesion, Regulation of actin cytoskeleton |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.-N.; Li, S.-L.; Yang, X.; Wang, J.-Q.; Zheng, N. The Protective Effects of Lactoferrin on Aflatoxin M1-Induced Compromised Intestinal Integrity. Int. J. Mol. Sci. 2022, 23, 289. https://doi.org/10.3390/ijms23010289
Gao Y-N, Li S-L, Yang X, Wang J-Q, Zheng N. The Protective Effects of Lactoferrin on Aflatoxin M1-Induced Compromised Intestinal Integrity. International Journal of Molecular Sciences. 2022; 23(1):289. https://doi.org/10.3390/ijms23010289
Chicago/Turabian StyleGao, Ya-Nan, Song-Li Li, Xue Yang, Jia-Qi Wang, and Nan Zheng. 2022. "The Protective Effects of Lactoferrin on Aflatoxin M1-Induced Compromised Intestinal Integrity" International Journal of Molecular Sciences 23, no. 1: 289. https://doi.org/10.3390/ijms23010289
APA StyleGao, Y. -N., Li, S. -L., Yang, X., Wang, J. -Q., & Zheng, N. (2022). The Protective Effects of Lactoferrin on Aflatoxin M1-Induced Compromised Intestinal Integrity. International Journal of Molecular Sciences, 23(1), 289. https://doi.org/10.3390/ijms23010289