
OMICS in Chronic Kidney Disease: Focus on Prognosis and Prediction

 ,
,  ,
, 
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Variability in Prognosis and Response to Treatments in CKD Patients
3. Omics: New Frontiers of Research
4. Prognostic Omics in CKD
5. Predictive Omics in CKD
6. Omics and Personalized Medicine in Nephrology: Future Perspectives and Conclusions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Provenzano, M.; Coppolino, G.; Faga, T.; Garofalo, C.; Serra, R.; Andreucci, M. Epidemiology of cardiovascular risk in chronic kidney disease patients: The real silent killer. Rev. Cardiovasc. Med. 2019, 20, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Andreucci, M.; Provenzano, M.; Faga, T.; Michael, A.; Patella, G.; Mastroroberto, P.; Serraino, G.F.; Bracale, U.M.; Ielapi, N.; Serra, R. Aortic Aneurysms, Chronic Kidney Disease and Metalloproteinases. Biomolecules 2021, 11, 194. [Google Scholar] [CrossRef]
- Hu, J.R.; Coresh, J. The public health dimension of chronic kidney disease: What we have learnt over the past decade. Nephrol. Dial. Transplant. 2017, 32 (Suppl. S2), ii113–ii120. [Google Scholar] [CrossRef] [Green Version]
- Tonelli, M.; Muntner, P.; Lloyd, A.; Manns, B.J.; Klarenbach, S.; Pannu, N.; James, M.T.; Hemmelgarn, B.R. Alberta Kidney Disease Network. Risk of coronary events in people with chronic kidney disease compared with those with diabetes: A population-level cohort study. Lancet 2012, 380, 807–814. [Google Scholar] [CrossRef]
- De Nicola, L.; Donfrancesco, C.; Minutolo, R.; Lo Noce, C.; Palmieri, L.; De Curtis, A.; Iacoviello, L.; Zoccali, C.; Gesualdo, L.; Conte, G.; et al. Prevalence and cardiovascular risk profile of chronic kidney disease in Italy: Results of the 2008–2012 National Health Examination Survey. Nephrol. Dial. Transplant. 2015, 30, 806–814. [Google Scholar] [CrossRef] [PubMed]
- GBD Chronic Kidney Disease Collaboration. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Provenzano, M.; Andreucci, M.; Garofalo, C.; Minutolo, R.; Serra, R.; De Nicola, L. Selective endothelin A receptor antagonism in patients with proteinuric chronic kidney disease. Expert Opin. Investig. Drugs 2021, 30, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Borrelli, S.; Liberti, M.E.; Andreucci, M.; Conte, G.; Minutolo, R.; Provenzano, M.; De Nicola, L. SGLT2 Inhibitors: Nephroprotective Efficacy and Side Effects. Medicina 2019, 55, 268. [Google Scholar] [CrossRef] [Green Version]
- Heerspink, H.J.L.; Parving, H.-H.; Andress, D.L.; Bakris, G.; Correa-Rotter, R.; Hou, F.-F.; Kitzman, D.W.; Kohan, D.; Makino, H.; McMurray, J.J.V.; et al. Atrasentan and renal events in patients with type 2 diabetes and chronic kidney disease (SONAR): A double-blind, randomised, placebo-controlled trial. Lancet 2019, 393, 1937–1947. [Google Scholar] [CrossRef]
- Bakris, G.L.; Agarwal, R.; Anker, S.D.; Pitt, B.; Ruilope, L.M.; Rossing, P.; Kolkhof, P.; Nowack, C.; Schloemer, P.; Joseph, A.; et al. Effect of Finerenone on Chronic Kidney Disease Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2020, 383, 2219–2229. [Google Scholar] [CrossRef]
- Provenzano, M.; Pelle, M.C.; Zaffina, I.; Tassone, B.; Pujia, R.; Ricchio, M.; Serra, R.; Sciacqua, A.; Michael, A.; Andreucci, M.; et al. Sodium-Glucose Co-transporter-2 Inhibitors and Nephroprotection in Diabetic Patients: More Than a Challenge. Front. Med. 2021, 8, 654557. [Google Scholar] [CrossRef]
- Provenzano, M.; De Nicola, L.; Pena, M.J.; Capitoli, G.; Garofalo, C.; Borrelli, S.; Gagliardi, I.; Antolini, L.; Andreucci, M. Precision Nephrology Is a Non-Negligible State of Mind in Clinical Research: Remember the Past to Face the Future. Nephron 2020, 144, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.; Mariani, L.H.; Kretzler, M. Integrated multi-omics approaches to improve classification of chronic kidney disease. Nat. Rev. Nephrol. 2020, 16, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Perco, P.; Pena, M.; Heerspink, H.J.L.; Mayer, G. Multimarker panels in diabetic kidney disease: The way to improved clinical trial design and clinical practice? Kidney Int. Rep. 2018, 4, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Pena, M.J.; Stenvinkel, P.; Kretzler, M.; Adu, D.; Agarwal, S.K.; Coresh, J.; Feldman, H.I.; Fogo, A.B.; Gansevoort, R.T.; Harris, D.C.; et al. Strategies to improve monitoring disease progression, assessing cardiovascular risk, and defining prognostic biomarkers in chronic kidney disease. Kidney Int. Suppl. 2017, 7, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Velde, M.; the Chronic Kidney Disease Prognosis Consortium; Matsushita, K.; Coresh, J.; Astor, B.C.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Gansevoort, R.T. Lower estimated glomerular filtration rate and higher albuminuria are associated with all-cause and cardiovascular mortality. A collaborative meta-analysis of high-risk population cohorts. Kidney Int. 2011, 79, 1341–1352. [Google Scholar] [CrossRef] [Green Version]
- Astor, B.C.; the Chronic Kidney Disease Prognosis Consortium; Matsushita, K.; Gansevoort, R.T.; van der Velde, M.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Coresh, J. Lower estimated glomerular filtration rate and higher albuminuria are associated with mortality and end-stage renal disease. A collaborative meta-analysis of kidney disease population cohorts. Kidney Int. 2011, 79, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Gansevoort, R.T.; Matsushita, K.; van der Velde, M.; Astor, B.C.; Woodward, M.; Levey, A.S.; de Jong, P.E.; Coresh, J.; Chronic Kidney Disease Prognosis Consortium. Lower estimated GFR and higher albuminuria are associated with adverse kidney outcomes. A collaborative meta-analysis of general and high-risk population cohorts. Kidney Int. 2011, 80, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nicola, L.; Provenzano, M.; Chiodini, P.; Borrelli, S.; Russo, L.; Bellasi, A.; Santoro, D.; Conte, G.; Minutolo, R. Epidemiology of low-proteinuric chronic kidney disease in renal clinics. PLoS ONE 2017, 12, e0172241. [Google Scholar] [CrossRef]
- Provenzano, M.; Minutolo, R.; Chiodini, P.; Bellizzi, V.; Nappi, F.; Russo, D.; Borrelli, S.; Garofalo, C.; Iodice, C.; De Stefano, T.; et al. Competing-Risk Analysis of Death and End Stage Kidney Disease by Hyperkalaemia Status in Non-Dialysis Chronic Kidney Disease Patients Receiving Stable Nephrology Care. J. Clin. Med. 2018, 7, 499. [Google Scholar] [CrossRef] [Green Version]
- De Nicola, L.; Minutolo, R.; Chiodini, P.; Zamboli, P.; Cianciaruso, B.; Nappi, F.; Signoriello, S.; Conte, G.; Zoccali, C.; SIN-TABLE CDK Study Group. Prevalence and prognosis of mild anemia in non-dialysis chronic kidney disease: A prospective cohort study in outpatient renal clinics. Am. J. Nephrol. 2010, 32, 533–540. [Google Scholar] [CrossRef]
- Minutolo, R.; Gabbai, F.B.; Chiodini, P.; Provenzano, M.; Borrelli, S.; Garofalo, C.; Bellizzi, V.; Russo, D.; Conte, G.; De Nicola, L. Collaborative Study Group on the Conservative Treatment of CKD of the Italian Society of Nephrology. Sex Differences in the Progression of CKD Among Older Patients: Pooled Analysis of 4 Cohort Studies. Am. J. Kidney Dis. 2020, 75, 30–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, R. Ambulatory blood pressure and cardiovascular events in chronic kidney disease. Semin. Nephrol. 2007, 27, 538–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, A.D.; Agarwal, R. The complex relationship between CKD and ambulatory blood pressure patterns. Adv. Chronic Kidney Dis. 2015, 22, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Brück, K.; Jager, K.J.; Zoccali, C.; Bello, A.K.; Minutolo, R.; Ioannou, K.; Verbeke, F.; Völzke, H.; Arnlöv, J.; Leonardis, D.; et al. Different rates of progression and mortality in patients with chronic kidney disease at outpatient nephrology clinics across Europe. Kidney Int. 2018, 93, 1432–1441. [Google Scholar] [CrossRef]
- Borrelli, S.; Leonardis, D.; Minutolo, R.; Chiodini, P.; De Nicola, L.; Esposito, C.; Mallamaci, F.; Zoccali, C.; Conte, G. Epidemiology of CKD Regression in Patients under Nephrology Care. PLoS ONE 2015, 10, e0140138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, E.J.; Hunsicker, L.G.; Clarke, W.R.; Berl, T.; Pohl, M.A.; Lewis, J.B.; Ritz, E.; Atkins, R.C.; Rohde, R.; Raz, I. Renoprotective effect of the angiotensin-receptor antagonist irbesartan in patients with nephropathy due to type 2 diabetes. N. Engl. J. Med. 2001, 345, 851–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrykiv, S.I.; de Zeeuw, D.; Persson, F.; Rossing, P.; Gansevoort, R.T.; Laverman, G.D.; Heerspink, H.J.L. Variability in response to albuminuria-lowering drugs: True or random? Br. J. Clin. Pharmacol. 2017, 83, 1197–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, M.; Chiodini, P.; Minutolo, R.; Zoccali, C.; Bellizzi, V.; Conte, G.; Locatelli, F.; Tripepi, G.; Del Vecchio, L.; Mallamaci, F.; et al. Reclassification of chronic kidney disease patients for end-stage renal disease risk by proteinuria indexed to estimated glomerular filtration rate: Multicentre prospective study in nephrology clinics. Nephrol. Dial. Transplant. 2020, 35, 138–147. [Google Scholar] [CrossRef]
- Schievink, B.; de Zeeuw, D.; Parving, H.H.; Rossing, P.; Lambers Heerspink, H.J. The renal protective effect of angiotensin receptor blockers depends on intra-individual response variation in multiple risk markers. Br. J. Clin. Pharmacol. 2015, 80, 678–686. [Google Scholar] [CrossRef] [Green Version]
- Petrykiv, S.I.; Laverman, G.D.; de Zeeuw, D.; Heerspink, H.J.L. The albuminuria-lowering response to dapagliflozin is variable and reproducible among individual patients. Diabetes Obes. Metab. 2017, 19, 1363–1370. [Google Scholar] [CrossRef]
- Heerspink, H.J.; Makino, H.; Andress, D.; Brennan, J.J.; Correa-Rotter, R.; Coll, B.; Davis, J.W.; Idler, K.; Kohan, D.E.; Liu, M.; et al. Comparison of exposure response relationship of atrasentan between North American and Asian populations. Diabetes Obes. Metab. 2016, 19, 545–552. [Google Scholar] [CrossRef]
- Minutolo, R.; Gabbai, F.B.; Provenzano, M.; Chiodini, P.; Borrelli, S.; Garofalo, C.; Sasso, F.C.; Santoro, D.; Bellizzi, V.; Conte, G.; et al. Cardiorenal prognosis by residual proteinuria level in diabetic chronic kidney disease: Pooled analysis of four cohort studies. Nephrol. Dial. Transplant. 2018, 33, 1942–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quantitative gene expression analysis in renal biopsies: A novel protocol for a high- throughput multicenter application. Kidney Int. 2002, 61, 133–140. [CrossRef] [PubMed] [Green Version]
- Park, J.; Shrestha, R.; Qiu, C.; Kondo, A.; Huang, S.; Werth, M.; Li, M.; Barasch, J.; Suszták, K. Single-cell transcriptomics of the mouse kidney reveals potential cellular targets of kidney disease. Science 2018, 360, 758–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinschen, M.M.; Limbutara, K.; Knepper, M.A.; Payne, D.M.; Pisitkun, T. From molecules to mechanisms: Functional proteomics and its application to renal tubule physiology. Physiol. Rev. 2018, 98, 2571–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalim, S.; Rhee, E.P. Metabolomics and kidney precision medicine. Clin. J. Am. Soc. Nephrol. 2017, 12, 1726–1727. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, M.; Garofalo, C.; Chiodini, P.; Mancuso, C.; Barbato, E.; De Nicola, L.; Andreucci, M. Ruolo della proteinuria nella ricerca clinica: Per ogni vecchia risposta, una nuova domanda [Role of proteinuria in clinical research: For each old-answer, a new key-question.]. Recenti. Prog. Med. 2020, 111, 74–81. [Google Scholar] [CrossRef]
- Lambers Heerspink, H.J.; Gansevoort, R.T. Albuminuria Is an Appropriate Therapeutic Target in Patients with CKD: The Pro View. Clin. J. Am. Soc. Nephrol. 2015, 10, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.; Hao, L.; Ricke, W.A.; Li, L. Biomarker discovery in mass spectrometry-based urinary proteomics. Proteom. Clin. Appl. 2015, 10, 358–370. [Google Scholar] [CrossRef]
- Di Meo, A.; Pasic, M.D.; Yousef, G.M. Proteomics and peptidomics: Moving toward precision medicine in urological malignancies. Oncotarget 2016, 7, 52460–52474. [Google Scholar] [CrossRef] [Green Version]
- Groopman, E.E.; Marasa, M.; Cameron-Christie, S.; Petrovski, S.; Aggarwal, V.S.; Rasouly, H.M.; Li, Y.; Zhang, J.; Nestor, J.; Krithivasan, P.; et al. Diagnostic Utility of Exome Sequencing for Kidney Disease. N. Engl. J. Med. 2019, 380, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Renkema, K.Y.; Stokman, M.F.; Giles, R.H.; Knoers, N.V. Next-generation sequencing for research and diagnostics in kidney disease. Nat. Rev. Nephrol. 2014, 10, 433–444. [Google Scholar] [CrossRef]
- Stokman, M.F.; Renkema, K.Y.; Giles, R.H.; Schaefer, F.; Knoers, N.V.; van Eerde, A.M. The expanding phenotypic spectra of kidney diseases: Insights from genetic studies. Nat. Rev. Nephrol. 2016, 12, 472–483. [Google Scholar] [CrossRef]
- Ju, W.; Smith, S.; Kretzler, M. Genomic biomarkers for chronic kidney disease. Transl. Res. 2012, 159, 290–302. [Google Scholar] [CrossRef] [Green Version]
- Connaughton, D.M.; Kennedy, C.; Shril, S.; Mann, N.; Murray, S.L.; Williams, P.A.; Conlon, E.; Nakayama, M.; van der Ven, A.T.; Ityel, H.; et al. Monogenic causes of chronic kidney disease in adults. Kidney Int. 2019, 95, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, E.; Nestor, J.G.; Gharavi, A.G. Clinical Genetic Screening in Adult Patients with Kidney Disease. Clin. J. Am. Soc. Nephrol. 2020, 15, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Lata, S.; Marasa, M.; Li, Y.; Fasel, D.A.; Groopman, E.; Jobanputra, V.; Rasouly, H.; Mitrotti, A.; Westland, R.; Verbitsky, M.; et al. Whole-exome sequencing in adults with chronic kidney disease: A pilot study. Ann. Intern. Med. 2018, 168, 100–109. [Google Scholar] [CrossRef]
- Wuttke, M.; Lifelines Cohort Study; Li, Y.; Li, M.; Sieber, K.B.; Feitosa, M.F.; Gorski, M.; Tin, A.; Wang, L.; Chu, A.Y.; et al. A catalog of genetic loci associated with kidney function from analyses of a million individuals. Nat. Genet. 2019, 51, 957–972. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Connelly, J.; Lindon, J.C.; Holmes, E. Metabonomics: A platform for studying drug toxicity and gene function. Nat. Rev. Drug Discov. 2002, 1, 153–161. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics--the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Emerging applications of metabolomics in drug discovery and precision medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, K.; Zhang, X. Next-generation metabolomics in lung cancer diagnosis, treatment and precision medicine: Mini review. Oncotarget 2017, 8, 115774–115786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubin, R.F.; Rhee, E.P. Proteomics and Metabolomics in Kidney Disease, including Insights into Etiology, Treatment, and Prevention. Clin. J. Am. Soc. Nephrol. 2019, 15, 404–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.Y. Metabolomics in chronic kidney disease. Clin. Chim. Acta 2013, 422, 59–69. [Google Scholar] [CrossRef]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Ryan, D.; Brennan, L.; Tenori, L.; Luchinat, C.; Gao, X.; Zeri, A.C.; Gowda, G.A.N.; et al. Recommendations and Standardization of Biomarker Quantification Using NMR-Based Metabolomics with Particular Focus on Urinary Analysis. J. Proteome Res. 2016, 15, 360–373. [Google Scholar] [CrossRef] [Green Version]
- Hajduk, J.; Matysiak, J.; Kokot, Z.J. Challenges in biomarker discovery with MALDI-TOF MS. Clin. Chim. Acta 2016, 458, 84–98. [Google Scholar] [CrossRef]
- Davies, R. The metabolomic quest for a biomarker in chronic kidney disease. Clin. Kidney J. 2018, 11, 694–703. [Google Scholar] [CrossRef]
- Rysz, J.; Gluba-Brzózka, A.; Franczyk, B.; Jabłonowski, Z.; Ciałkowska-Rysz, A. Novel Biomarkers in the Diagnosis of Chronic Kidney Disease and the Prediction of Its Outcome. Int. J. Mol. Sci. 2017, 18, 1702. [Google Scholar] [CrossRef]
- Bryant, S.; Manning, D.L. Isolation of messenger RNA. Methods Mol. Biol. 1998, 86, 61–64. [Google Scholar] [CrossRef]
- Hobby, G.P.; Karaduta, O.; Dusio, G.F.; Singh, M.; Zybailov, B.L.; Arthur, J.M. Chronic kidney disease and the gut microbiome. Am. J. Physiol. Physiol. 2019, 316, F1211–F1217. [Google Scholar] [CrossRef]
- Roy, J.; Shou, H.; Xie, D.; Hsu, J.Y.; Yang, W.; Anderson, A.H.; Landis, J.R.; Jepson, C.; He, J.; Liu, K.D.; et al. Chronic Renal Insufficiency Cohort (CRIC) Study Investigators. Statistical Methods for Cohort Studies of CKD: Prediction Modeling. Clin. J. Am. Soc. Nephrol. 2016, 12, 1010–1017. [Google Scholar] [CrossRef] [Green Version]
- Tibshirani, R. Regression shrinkage and selection via the lasso. J. R. Stat. Soc. Ser. B Stat. Methodol. 1994, 58, 267–288. [Google Scholar] [CrossRef]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Mariani, M.C.; Tweneboah, O.K.; Bhuiyan, M.A.M. Supervised machine learning models applied to disease diagnosis and prognosis. AIMS Public Health 2019, 6, 405–423. [Google Scholar] [CrossRef]
- Tangri, N.; Grams, M.E.; Levey, A.S.; Coresh, J.; Appel, L.J.; Astor, B.C.; Chodick, G.; Collins, A.J.; Djurdjev, O.; Elley, C.R.; et al. CKD Prognosis Consortium.Multinational assessment of accuracy of equations for predicting risk of kidney failure: A meta-analysis. JAMA 2016, 315, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, K.; Coresh, J.; Sang, Y.; Chalmers, J.; Fox, C.; Guallar, E.; Jafar, T.; Jassal, S.K.; Landman, G.W.; Muntner, P.; et al. Estimated glomerular filtration rate and albuminuria for prediction of cardiovascular outcomes: A collaborative meta-analysis of individual participant data. Lancet Diabetes Endocrinol. 2015, 3, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Kadatz, M.J.; Lee, E.S.; Levin, A. Predicting Progression in CKD: Perspectives and Precautions. Am. J. Kidney Dis. 2016, 67, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Simeoni, M.; Citraro, M.L.; Cerantonio, A.; Deodato, F.; Provenzano, M.; Cianfrone, P.; Capria, M.; Corrado, S.; Libri, E.; Comi, A.; et al. An open-label, randomized, placebo-controlled study on the effectiveness of a novel probiotics administration protocol (ProbiotiCKD) in patients with mild renal insufficiency (stage 3a of CKD). Eur. J. Nutr. 2018, 58, 2145–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haubitz, M.; Wittke, S.; Weissinger, E.M.; Walden, M.; Rupprecht, H.D.; Floege, J.; Haller, H.; Mischak, H. Urine protein patterns can serve as diagnostic tools in patients with IgA nephropathy. Kidney Int. 2005, 67, 2313–2320. [Google Scholar] [CrossRef] [Green Version]
- Weissinger, E.M.; Wittke, S.; Kaiser, T.; Haller, H.; Bartel, S.; Krebs, R.; Golovko, I.; Rupprecht, H.D.; Haubitz, M.; Hecker, H.; et al. Proteomic patterns established with capillary electrophoresis and mass spectrometry for diagnostic purposes. Kidney Int. 2004, 65, 2426–2434. [Google Scholar] [CrossRef] [Green Version]
- Fliser, D.; Novak, J.; Thongboonkerd, V.; Argilés, A.; Jankowski, V.; Girolami, M.A.; Jankowski, J.; Mischak, H. Advances in urinary proteome analysis and biomarker discovery. J. Am. Soc. Nephrol. 2007, 18, 1057–1071. [Google Scholar] [CrossRef] [Green Version]
- Mischak, H.; Kaiser, T.; Walden, M.; Hillmann, M.; Wittke, S.; Herrmann, A.; Knueppel, S.; Haller, H.; Fliser, D. Proteomic analysis for the assessment of diabetic renal damage in humans. Clin. Sci. 2004, 107, 485–495. [Google Scholar] [CrossRef]
- Siwy, J.; Zürbig, P.; Argiles, A.; Beige, J.; Haubitz, M.; Jankowski, J.; Julian, B.A.; Linde, P.G.; Marx, D.; Mischak, H.; et al. Noninvasive diagnosis of chronic kidney diseases using urinary proteome analysis. Nephrol. Dial. Transplant. 2016, 32, 2079–2089. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.; Staplin, N.; Emberson, J.; Herrington, W.G.; Tomson, C.; Agodoa, L.; Tesar, V.; Levin, A.; Lewis, D.; Reith, C.; et al. Evaluating the Contribution of the Cause of Kidney Disease to Prognosis in CKD: Results from the Study of Heart and Renal Protection (SHARP). Am. J. Kidney Dis. 2014, 64, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, D.M.; Zürbig, P.; Argilés, A.; Bauer, H.W.; Behrens, G.; Coon, J.J.; Dakna, M.; Decramer, S.; Delles, C.; Dominiczak, A.F.; et al. Naturally occurring human urinary peptides for use in diagnosis of chronic kidney disease. Mol. Cell. Proteom. 2010, 9, 2424–2437. [Google Scholar] [CrossRef] [Green Version]
- Argilés, Á.; Siwy, J.; Duranton, F.; Gayrard, N.; Dakna, M.; Lundin, U.; Osaba, L.; Delles, C.; Mourad, G.; Weinberger, K.M.; et al. CKD273, a new proteomics classifier assessing CKD and its prognosis. PLoS ONE 2013, 8, e62837. [Google Scholar] [CrossRef] [Green Version]
- Schanstra, J.P.; Zürbig, P.; Alkhalaf, A.; Argiles, A.; Bakker, S.J.; Beige, J.; Bilo, H.J.; Chatzikyrkou, C.; Dakna, M.; Dawson, J.; et al. Diagnosis and Prediction of CKD Progression by Assessment of Urinary Peptides. J. Am. Soc. Nephrol. 2015, 26, 1999–2010. [Google Scholar] [CrossRef]
- Verbeke, F.; Siwy, J.; Van Biesen, W.; Mischak, H.; Pletinck, A.; Schepers, E.; Neirynck, N.; Magalhães, P.; Pejchinovski, M.; Pontillo, C.; et al. The urinary proteomics classifier chronic kidney disease 273 predicts cardiovascular outcome in patients with chronic kidney disease. Nephrol. Dial. Transplant. 2019, 36, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, J.M.; Thongboonkerd, V.; Scherzer, J.A.; Cai, J.; Pierce, W.M.; Klein, J.B. Differential expression of proteins in renal cortex and medulla: A proteomic approach. Kidney Int. 2002, 62, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.J.; Shyr, Y.; Liang, X.; Ma, L.J.; Donnert, E.M.; Roberts, J.D.; Zhang, X.; Kon, V.; Brown, N.J.; Caprioli, R.M.; et al. Proteomic patterns and prediction of glomerulosclerosis and its mechanisms. J. Am. Soc. Nephrol. 2005, 16, 2967–2975. [Google Scholar] [CrossRef] [Green Version]
- Kislinger, T.; Cox, B.; Kannan, A.; Chung, C.; Hu, P.; Ignatchenko, A.; Scott, M.S.; Gramolini, A.O.; Morris, Q.; Hallett, M.T.; et al. Global survey of organ and organelle protein expression in mouse: Combined proteomic and transcriptomic profiling. Cell 2006, 125, 173–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mischak, H. Pro: Urine proteomics as a liquid kidney biopsy: No more kidney punctures! Nephrol. Dial. Transplant. 2015, 30, 532–537. [Google Scholar] [CrossRef] [Green Version]
- Hellwege, J.N.; Velez Edwards, D.R.; Giri, A.; Qiu, C.; Park, J.; Torstenson, E.S.; Keaton, J.M.; Wilson, O.D.; Robinson-Cohen, C.; Chung, C.P.; et al. Mapping eGFR loci to the renal transcriptome and phenome in the VA Million Veteran Program. Nat. Commun. 2019, 10, 3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köttgen, A.; Albrecht, E.; Teumer, A.; Vitart, V.; Krumsiek, J.; Hundertmark, C.; Pistis, G.; Ruggiero, D.; O’Seaghdha, C.M.; Haller, T.; et al. Genome-wide association analyses identify 18 new loci associated with serum urate concentrations. Nat. Genet. 2012, 45, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Teumer, A.; Tin, A.; Sorice, R.; Gorski, M.; Yeo, N.C.; Chu, A.Y.; Li, M.; Li, Y.; Mijatovic, V.; Ko, Y.-A.; et al. Genome-wide Association Studies Identify Genetic Loci Associated with Albuminuria in Diabetes. Diabetes 2015, 65, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; De Stefano, T.; Vita, C.; Vinci, G.; Balia, F.; Nettuno, F.; Scarpati, L.; Sguazzo, A.; Sagliocchi, A.; Pacilio, M.; et al. Hyperuricaemia and Chronic Kidney Disease. G Italy Nefrol. 2018, 35, 2018-vol1. [Google Scholar]
- Mooyaart, A.L.; Valk, E.J.; van Es, L.A.; Bruijn, J.A. Genetic associations in diabetic nephropathy: A meta-analysis. Diabetologia 2011, 54, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.C.; Cooper, M.E. Genetics of diabetic kidney disease-from the worst of nightmares to the light of dawn? J. Am. Soc. Nephrol. 2017, 28, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Gillies, C.E.; Putler, R.; Menon, R.; Otto, E.; Yasutake, K.; Nair, V.; Hoover, P.; Lieb, D.; Li, S.; Eddy, S.; et al. An eQTL Landscape of Kidney Tissue in Human Nephrotic Syndrome. Am. J. Hum. Genet. 2018, 103, 232–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, C.; Huang, S.; Park, J.; Park, Y.; Ko, Y.A.; Seasock, M.J.; Bryer, J.S.; Xu, X.X.; Song, W.C.; Palmer, M.; et al. Renal compartment-specic genetic variation analyses identify new pathways in chronic kidney disease. Nat. Med. 2018, 24, 1721–1731. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, S.K.; Sedor, J.R.; Freedman, B.I.; Kao, W.H.; Kretzler, M.; Keller, B.J.; Abboud, H.E.; Adler, S.G.; Best, L.G.; Bowden, D.W.; et al. Family Investigation of Nephropathy and Diabetes (FIND). Genome-Wide Association and Trans-ethnic Meta-Analysis for Advanced Diabetic Kidney Disease: Family Investigation of Nephropathy and Diabetes (FIND). PLoS Genet. 2015, 11, e1005352. [Google Scholar] [CrossRef] [PubMed]
- Satirapoj, B.; Tasanavipas, P.; Supasyndh, O. Role of TCF7L2 and PPARG2 Gene Polymorphisms in Renal and Cardiovascular Complications among Patients with Type 2 Diabetes: A Cohort Study. Kidney Dis. 2019, 5, 220–227. [Google Scholar] [CrossRef]
- Valdivielso, J.M.; Bozic, M.; Galimudi, R.K.; Bermudez-López, M.; Navarro-González, J.F.; Fernández, E.; Betriu, À. Association of the rs495392 Klotho polymorphism with atheromatosis progression in patients with chronic kidney disease. Nephrol. Dial. Transplant. 2018, 34, 2079–2088. [Google Scholar] [CrossRef]
- Ferguson, J.F.; Matthews, G.J.; Townsend, R.R.; Raj, D.S.; Kanetsky, P.A.; Budoff, M.; Fischer, M.J.; Rosas, S.E.; Kanthety, R.; Rahman, M.; et al. CRIC Study Principal Investigators. Candidate gene association study of coronary artery calcification in chronic kidney disease: Findings from the CRIC study (Chronic Renal Insufficiency Cohort). J. Am. Coll. Cardiol. 2013, 62, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Dudley, A.J.; Bleasby, K.; Brown, C.D. The organic cation transporter OCT2 mediates the uptake of β-adrenoceptor antagonists across the apical membrane of renal LLC-PK1 cell monolayers. Br. J. Pharm. 2000, 131, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Filipski, K.K.; Mathijssen, R.H.; Mikkelsen, T.S.; Schinkel, A.H.; Sparreboom, A. Contribution of organic cation transporter 2 (OCT2) to cisplatin-induced nephrotoxicity. Clin. Pharmacol. Ther. 2009, 86, 396–402. [Google Scholar] [CrossRef]
- Motohashi, H.; Inui, K. Organic cation transporter OCTs (SLC22) and MATEs (SLC47) in the human kidney. AAPS J. 2013, 15, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Kalim, S.; Rhee, E.P. An overview of renal metabolomics. Kidney Int. 2016, 91, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Rhee, E.P.; Clish, C.B.; Ghorbani, A.; Larson, M.G.; Elmariah, S.; McCabe, E.; Yang, Q.; Cheng, S.; Pierce, K.; Deik, A.; et al. A combined epidemiologic and metabolomic approach improves CKD prediction. J. Am. Soc. Nephrol. 2013, 24, 1330–1338. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Simonavicius, N.; Wu, X.; Swaminath, G.; Reagan, J.; Tian, H.; Ling, L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. J. Biol. Chem. 2006, 281, 22021–22028. [Google Scholar] [CrossRef] [Green Version]
- Rhee, E.P.; Clish, C.B.; Wenger, J.; Roy, J.; Elmariah, S.; Pierce, K.A.; Bullock, K.; Anderson, A.H.; Gerszten, R.E.; Feldman, H.I. Metabolomics of Chronic Kidney Disease Progression: A Case-Control Analysis in the Chronic Renal Insufficiency Cohort Study. Am. J. Nephrol. 2016, 3, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular Risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, J.R.; House, J.A.; Ocque, A.J.; Zhang, S.; Johnson, C.; Kimber, C.; Schmidt, K.; Gupta, A.; Wetmore, J.B.; Nolin, T.D.; et al. Serum Trimethylamine-N-Oxide is Elevated in CKD and Correlates with Coronary Atherosclerosis Burden. J. Am. Soc. Nephrol. 2016, 27, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missailidis, C.; Hällqvist, J.; Qureshi, A.R.; Barany, P.; Heimbürger, O.; Lindholm, B.; Stenvinkel, P.; Bergman, P. Serum Trimethylamine-N-Oxide Is Strongly Related to Renal Function and Predicts Outcome in Chronic Kidney Disease. PLoS ONE 2016, 11, e0141738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.; Karl, B.; Mathew, A.; Gangoiti, J.A.; Wassel, C.L.; Saito, R.; Pu, M.; Sharma, S.; You, Y.-H.; Wang, L.; et al. Metabolomics Reveals Signature of Mitochondrial Dysfunction in Diabetic Kidney Disease. J. Am. Soc. Nephrol. 2013, 24, 1901–1912. [Google Scholar] [CrossRef]
- You, Y.-H.; Quach, T.; Saito, R.; Pham, J.; Sharma, K. Metabolomics Reveals a Key Role for Fumarate in Mediating the Effects of NADPH Oxidase 4 in Diabetic Kidney Disease. J. Am. Soc. Nephrol. 2015, 27, 466–481. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.; Ranches, G.; Schafferer, S.; Lukasser, M.; Rudnicki, M.; Mayer, G.; Hüttenhofer, A. Identification of urinary exosomal noncoding RNAs as novel biomarkers in chronic kidney disease. RNA 2016, 23, 142–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Arce, L.; Natarajan, R. MicroRNAs and their role in progressive kidney diseases. Clin. J. Am. Soc. Nephrol. 2009, 4, 1255–1266. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Srinivasan, L.; Calado, D.P.; Patterson, H.C.; Zhang, B.; Wang, J.; Henderson, J.M.; Kutok, J.L.; Rajewsky, K. Lymphoproliferative disease and autoimmunity in mice with increased miR-17–92 expression in lymphocytes. Nat. Immunol. 2008, 9, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Yu, L.; Chiu, C.; Sun, Y.; Chen, J.; Khitrov, G.; Merkenschlager, M.; Holzman, L.B.; Zhang, W.; Mundel, P.; et al. Podocyte-selective deletion of dicer induces proteinuria and glomerulosclerosis. J. Am. Soc. Nephrol. 2008, 19, 2159–2169. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Ng, K.H.; Rosen, S.; Dostal, A.; Gregory, R.I.; Kreidberg, J.A. Podocyte-specific loss of functional microRNAs leads to rapid glomerular and tubular injury. J. Am. Soc. Nephrol. 2008, 19, 2069–2075. [Google Scholar] [CrossRef] [Green Version]
- Taïbi, F.; Metzinger-Le Meuth, V.; M’Baya-Moutoula, E.; Djelouat, M.S.; Louvet, L.; Bugnicourt, J.M.; Poirot, S.; Bengrine, A.; Chillon, J.M.; Massy, Z.A.; et al. Possible involvement of microRNAs in vascular damage in experimental chronic kidney disease. Biochim. Biophys. Acta 2014, 1842, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol. Res. 2018, 2018, 1–16. [Google Scholar] [CrossRef]
- Al Khodor, S.; Shatat, I.F. Gut microbiome and kidney disease: A bidirectional relationship. Pediatr. Nephrol. 2016, 32, 921–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, M.; Rotundo, S.; Chiodini, P.; Gagliardi, I.; Michael, A.; Angotti, E.; Borrelli, S.; Serra, R.; Foti, D.; De Sarro, G.; et al. Contribution of Predictive and Prognostic Biomarkers to Clinical Research on Chronic Kidney Disease. Int. J. Mol. Sci. 2020, 21, 5846. [Google Scholar] [CrossRef] [PubMed]
- Rossing, K.; Mischak, H.; Parving, H.H.; Christensen, P.K.; Walden, M.; Hillmann, M.; Kaiser, T. Impact of diabetic nephropathy and angiotensin II receptor blockade on urinary polypeptide patterns. Kidney Int. 2005, 68, 193–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tofte, N.; Lindhardt, M.; Adamova, K.; Bakker, S.J.L.; Beige, J.; Beulens, J.W.J.; Birkenfeld, A.L.; Currie, G.; Delles, C.; Dimos, I.; et al. Early detection of diabetic kidney disease by urinary proteomics and subsequent intervention with spironolactone to delay progression (PRIORITY): A prospective observational study and embedded randomised placebo-controlled trial. Lancet Diabetes Endocrinol. 2020, 8, 301–312. [Google Scholar] [CrossRef]
- Andersen, S.; Mischak, H.; Zurbig, P.; Parving, H.H.; Rossing, P. Urinary proteome analysis enables assessment of renoprotective treatment in type 2 diabetic patients with microalbuminuria. BMC Nephrol. 2010, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindhart, M.; Persson, F.; Oxlund, C.; Jacobsen, A.; Zurbig, P.; Mischak, H.; Rossing, P.; Heerspink, H.J.L. Predicting albuminuria response to spironolactone treatment with urinary proteomics in patients with type 2 diabetes and hypertension. Nephrol. Dial. Transplant. 2018, 33, 296–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siwy, J.; Klein, T.; Rosler, M.; von Eynatten, M. Urinary proteomics as a tool to identify kidney responders to dipeptidyl peptidase-4 inhibition: A hypothesis-generating analysis from the MARLINA-T2D trial. Proteom. Clin. Appl. 2019, 13, 1800144. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, M.T.; Centra, M.; Papale, M.; Bortone, G.; Palermo, C.; Centonze, D.; Renieri, E.; Di Paolo, S.; Gesualdo, L. Urine protein profile of IgA nephropathy patients may predict the response to ACE-inhibitor therapy. Proteomics 2008, 8, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Perna, A.; Ruggenenti, P.; Testa, A.; Spoto, B.; Benini, R.; Misefari, V.; Remuzzi, G.; Zoccali, C.; Gruppo Italiano di Studi Epidemiologici in Nefrologia. ACE genotype and ACE inhibitors induced enoprotection in chronic proteinuric nephropathies. Kidney Int. 2000, 57, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Parving, H.H.; de Zeeuw, D.; Cooper, M.E.; Remuzzi, G.; Liu, N.; Lunceford, J.; Shahinfar, S.; Wong, P.H.; Lyle, P.A.; Rossing, P.; et al. ACE gene polymorphism and losartan treatment in type 2 diabetic patients with nephropathy. J. Am. Soc. Nephrol. 2008, 19, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzkova, H.; Pechandova, K.; Danzig, V.; Vařeka, T.; Perlík, F.; Žák, A.; Slanař, O. Lipid-lowering effect of fluvastatin in relation to cytochrome P450 2C9 variant alleles frequently distributed in the Czech population. Med. Sci. Monit. 2012, 18, CR512–CR517. [Google Scholar] [CrossRef] [Green Version]
- Luvai, A.; Mbagaya, W.; Hall, A.S.; Barth, J.H. Rosuvastatin: A review of the pharmacology and clinical effectiveness in cardiovascular disease. Clin. Med. Insights Cardiol. 2012, 6, 17–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwinyi, J.; Johne, A.; Bauer, S.; Roots, I.; Gerloff, T. Evidence for inverse effects of OATP-C (SLC21A6) 5 and 1b haplotypes on pravastatin kinetics. Clin. Pharmacol. Ther. 2004, 75, 415–421. [Google Scholar] [CrossRef]
- Hoeben, E.; De Winter, W.; Neyens, M.; Devineni, D.; Vermeulen, A.; Dunne, A. Population Pharmacokinetic Modeling of Canagliflozin in Healthy Volunteers and Patients with Type 2 Diabetes Mellitus. Clin. Pharm. 2016, 55, 209–223. [Google Scholar] [CrossRef]
- Francke, S.; Mamidi, R.N.V.S.; Ms, B.S.; Scheers, E.; Jadwin, A.; Favis, R.; Devineni, D. In vitro metabolism of canagliflozin in human liver, kidney, intestine microsomes, and recombinant uridine diphosphate glucuronosyltransferases (UGT) and the effect of genetic variability of UGT enzymes on the pharmacokinetics of canagliflozin in humans. J. Clin. Pharmacol. 2015, 55, 1061–1072. [Google Scholar] [CrossRef]
- Jin, T. Current Understanding on Role of the Wnt Signaling Pathway Effector TCF7L2 in Glucose Homeostasis. Endocr. Rev. 2016, 37, 254–277. [Google Scholar] [CrossRef] [Green Version]
- Tran, M.T.; Zsengeller, Z.K.; Berg, A.H.; Khankin, E.V.; Bhasin, M.K.; Kim, W.; Clish, C.B.; Stillman, I.E.; Karumanchi, S.A.; Rhee, E.P.; et al. PGC1α drives NAD biosynthesis linking oxidative metabolism to renal protection. Nature 2016, 531, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Denby, L.; Baker, A.H. Targeting non-coding RNA for the therapy of renal disease. Curr. Opin. Pharmacol. 2016, 27, 70–77. [Google Scholar] [CrossRef]
- Gomez, I.G.; MacKenna, D.A.; Johnson, B.G.; Kaimal, V.; Roach, A.M.; Ren, S.; Nakagawa, N.; Xin, C.; Newitt, R.; Pandya, S.; et al. Anti-microRNA-21 oligonucleotides prevent Alport nephropathy progression by stimulating metabolic pathways. J. Clin. Investig. 2015, 125, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.D.; Putta, S.; Wang, M.; Lai, J.Y.; Bitzer, M.; Nelson, R.G.; Lanting, L.L.; Kato, M.; Natarajan, R. Trans-forming growth factor-β-induced cross talk between p53 and a microRNA in the pathogenesis of diabetic nephropathy. Diabetes 2013, 62, 3151–3162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krupa, A.; Jenkins, R.; Luo, D.D.; Lewis, A.; Phillips, A.; Fraser, D. Loss of MicroRNA-192 promotes fibrogenesis in diabetic nephropathy. J. Am. Soc. Nephrol. 2010, 21, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Su, P.Y.; Lee, Y.H.; Kuo, L.N.; Chen, Y.C.; Chen, C.; Kang, Y.N.; Chang, E.H. Efficacy of AST-120 for Patients with Chronic Kidney Disease: A Network Meta-Analysis of Randomized Controlled Trials. Front. Pharmacol. 2021, 12, 676345. [Google Scholar] [CrossRef] [PubMed]
- Rossello, X.; Dorresteijn, J.A.; Janssen, A.; Lambrinou, E.; Scherrenberg, M.; Bonnefoy-Cudraz, E.; Cobain, M.; Piepoli, M.F.; Visseren, F.L.; Dendale, P. Risk prediction tools in cardiovascular disease prevention: A report from the ESC Prevention of CVD Programme led by the European Association of Preventive Cardiology (EAPC) in collaboration with the Acute Cardiovascular Care Association (ACCA) and the Association of Cardiovascular Nursing and Allied Professions (ACNAP). Eur. Heart J. Acute Cardiovasc. Care 2019, 18, 534–544. [Google Scholar]
- Weiner, D.E.; Tighiouart, H.; Elsayed, E.F.; Griffith, J.L.; Salem, D.N.; Levey, A.S.; Sarnak, M.J. The Framingham predictive instrument in chronic kidney disease. J. Am. Coll. Cardiol. 2007, 50, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Ielapi, N.; Andreucci, M.; Licastro, N.; Faga, T.; Grande, R.; Buffone, G.; Mellace, S.; Sapienza, P.; Serra, R. Precision Medicine and Precision Nursing: The Era of Biomarkers and Precision Health. Int. J. Gen. Med. 2020, 13, 1705–1711. [Google Scholar] [CrossRef]
- Busceti, M.T.; Grande, R.; Amato, B.; Gasbarro, V.; Buffone, G.; Amato, M.; Gallelli, L.; Serra, R.; de Franciscis, S. Pulmonary embolism, metalloproteinases and neutrophil gelatinase associated lipocalin. Acta Phlebol. 2013, 14, 115–121. [Google Scholar]
- Russo, D.; Morrone, L.F.; Errichiello, C.; De Gregorio, M.G.; Imbriaco, M.; Battaglia, Y.; Russo, L.; Andreucci, M.; Di Iorio, B.R. Impact of BMI on cardiovascular events, renal function, and coronary artery calcification. Blood Purif. 2014, 38, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Battaglia, Y. Clinical Significance of FGF-23 in Patients with CKD. Int. J. Nephrol. 2011, 2011, 364890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, D.; Corrao, S.; Battaglia, Y.; Andreucci, M.; Caiazza, A.; Carlomagno, A.; Lamberti, M.; Pezone, N.; Pota, A.; Russo, L.; et al. Progression of coronary artery calcification and cardiac events in patients with chronic renal disease not receiving dialysis. Kidney Int. 2011, 80, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provenzano, M.; Serra, R.; Michael, A.; Bolignano, D.; Coppolino, G.; Ielapi, N.; Serraino, G.F.; Mastroroberto, P.; Locatelli, F.; De Nicola, L.; et al. Smoking habit as a risk amplifier in chronic kidney disease patients. Sci. Rep. 2021, 11, 14778. [Google Scholar] [CrossRef] [PubMed]
- Minutolo, R.; Lapi, F.; Chiodini, P.; Simonetti, M.; Bianchini, E.; Pecchioli, S.; Cricelli, I.; Cricelli, C.; Piccinocchi, G.; Conte, G.; et al. Risk of ESRD and death in patients with CKD not referred to a nephrologist: A 7-year prospective study. Clin. J. Am. Soc. Nephrol. 2014, 9, 1586–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minutolo, R.; De Nicola, L.; Mazzaglia, G.; Postorino, M.; Cricelli, C.; Mantovani, L.G.; Conte, G.; Cianciaruso, B. Detection and awareness of moderate to advanced CKD by primary care practitioners: A cross-sectional study from Italy. Am. J. Kidney Dis. 2008, 52, 444–453. [Google Scholar] [CrossRef]
- Ju, W.; Nair, V.; Smith, S.; Zhu, L.; Shedden, K.; Song, P.X.; Mariani, L.H.; Eichinger, F.H.; Berthier, C.C.; Randolph, A.; et al. Tissue transcriptome- driven identificationof epidermal growth factor as a chronic kidney diseasebiomarker. Sci. Transl. Med. 2015, 7, 316ra193. [Google Scholar] [CrossRef] [Green Version]
- Martini, S.; Nair, V.; Keller, B.J.; Eichinger, F.; Hawkins, J.J.; Randolph, A.; Böger, C.A.; Gadegbeku, C.A.; Fox, C.S.; Cohen, C.D.; et al. European Renal cDNA Bank; C-PROBE Cohort; CKDGen Consortium. Integrative biology identifies shared transcriptional networks in CKD. J. Am. Soc. Nephrol. 2014, 25, 2559–2572. [Google Scholar] [CrossRef] [Green Version]
- Glassock, R.J. Con: Kidney biopsy: An irreplaceable tool for patient management in nephrology. Nephrol. Dial Transplant. 2015, 30, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Ahn, S.H.; Choi, P.; Ko, Y.A.; Han, S.H.; Chinga, F.; Park, A.S.; Tao, J.; Sharma, K.; Pullman, J.; et al. Defective fatty acid oxidation in renal tubular epithelial cells has a key role in kidney fibrosis development. Nat. Med. 2015, 21, 37–46. [Google Scholar] [CrossRef]


{kind=link}
| Name | Types | Source | Role | Outcome | Disease | Note/Comments |
|---|---|---|---|---|---|---|
| Human studies | ||||||
| CDK273 [78,79] | Peptide | Urine | Prognostic | ESKD onset or eGFR decline; CV events | CKD | High CKD273 score was associated with an increased individual risk for CKD progression. |
| PKD1 & NOS3 [85] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | Decrease in renal function (eGFR) | Renal function | Mutations of PKD1, encoding polycystin-1 and NOS3, involving nitric oxide production, have been associated with reduced renal function. |
| TRIM46, INHBB, SFMBT1, TMEM171, VEGFA, BAZ1B [86,87] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | CV events | CKD | SNPs in these genes influence serum uric acid levels and this association partially explains the increased CV risk in CKD. |
| C9orf3 and variant rs334 of HBB encoding beta-globin [86,87] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | CV events | CKD | These genetic variants influence urine albumin excretion and mediate the association between CKD and CV events. |
| ZFHX3, PMF1-BGLAP, USP38, and TTBK1 [87] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | Cerebrovascular accidents | CKD | SNPs in these genes influence serum uric acid levels and this association partially explains the increased risk for cerebrovascular accidents in CKD patients. |
| UMOD [91,92] | Gene | Uromodulin | Prognostic | Disorders in diabetic nephropathy | Diabetic nephropathy | The variants rs77924615 and rs111285796 were found to predict the risk of nephrotic syndrome. |
| PPARG2 [94] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | Cerebrovascular events | CKD patients | Variant Pro12Ala was able to predict cerebrovascular events in CKD patients. |
| Klotho [95] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | Progression of endothelial dysfunction and arterial stiffness, CV events | CKD | Genetic variants of Klotho gene influence CV risk and progression of atherosclerosis in CKD patients. |
| Chr9p21, COL4A1, ATP2B1, and HNF4A [96] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | CV events | CKD | These genes are involved in regulation of blood pressure, vascular tone, and calcium homeostasis and their variants predict coronary artery disease in CKD patients. |
| MATE1 [99] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | Severity of CKD | CKD | MATE1 secretes drugs from cells into the lumen of proximal tubules. Several genetic polymorphisms such as the variant Ala465Val of SLC47A1 may affect the function of this transporter with impaired secretion of toxins and drugs, which reflect on the severity of CKD. |
| Trimethylamine-N-oxide (TMAO) [105] | Amine oxides | Serum | Prognostic | Stroke, CV acute events, and mortality over time | CKD | Trimethylamine-N-oxide (TMAO) plasma levels are strictly associated with the incidence of stroke, CV acute events, and mortality over time. TMAO levels increase with the progression of kidney damage. Therefore, this marker could perform even better in CKD patients. |
| NOX4 [109] | Gene | Single nucleotide polymorphisms (SNPs) | Prognostic | Severity of CKD | CKD | NOX4 expression increases fumarate levels, which are linked to glomerular dysfunction. Therefore, fumarate is a key link connecting metabolic pathways to diabetic nephropathy. |
| miR-222-3p, miR-27a-3p, miR-27b-3p, miR-877-3p, miR-31-5p, miR-3687, let-7c-5p, miR-6769b-5p miR-296-5p miR-133a, miR-133b, miR-15a-5p, miR-181a-5p, miR-34a-5p, miR-181c-5p miR1-2 [111] | miRNAs | Non-coding RNA fragments | Prognostic | Severity of CKD | CKD | These miRNAs are differentially expressed in CKD patients. miRNAs associated with CKD impair the degree of fibrosis, ECM deposition and proteinuria and accelerate CKD progression. |
| p-cresyl-sulphate indoxyl sulphate [117] | Protein-bound uremic toxins | Microbiomics | Prognostic | Severity of CV damage | CKD | Overexpression of uremic toxins accelerate CKD progression. |
| CKD273 [120,121,122,123] | Peptide | Urine | Predictive | Response to RAASi/DPP-4 | Diabetic nephropathy | CKD273 panel is not only a prognostic but also a predictive tool. There is a close relationship between a high CKD273 score and response to RAASi or Linagliptin therapy. In high-risk patients undergoing therapy with the latter, the CKD273 score had a significant decrease compared with healthy subjects. |
| Urine kininogen [75,124] | Peptide | Urine | Predictive | Response to RAASi | CKD | Urine kininogen could predict the response to therapy with RAASi. However, further studies are needed. |
| Angiotensin-converting enzyme gene polymorphisms [125,126] | Gene | Single nucleotide polymorphisms (SNPs) | Predictive | Response to RAASi | CKD | Polymorphisms (insertion or deletion) for the gene encoding the angiotensin-converting enzyme may predict the response to RAASi. In one study, the D/D variant, followed by the I/D variant, resulted in a greater reduction in proteinuria, and better renal function over time. In contrast, the I/I variant predicted poor response and less benefit from RAASi therapy. |
| SLCO1B1, ABCB1, ABCC2, ABCG2 and ABCB11 [127,128] | Gene | Single nucleotide polymorphisms (SNPs) | Predictive | Response to statins and consequent increased CV risk | CKD | Several polymorphisms (SLCO1B1, ABCB1, ABCC2, ABCG2, and ABCB11) for the gene encoding cytochrome P450 could affect the response to statins, which play a central role in reducing CV risk among CKD patients. |
| UGT1A9 [129,130,131] | Gene | Single nucleotide polymorphisms (SNPs) | Predictive | Pharmacokinetics of SGLT2i | CKD and diabetes | UGT1A9 gene translates for an enzyme involved in the pharmacokinetics of SGLT2i. Carriers of the variants UGT1A9*3 and UGT2B4*2 have higher plasma levels of drugs, which are associated with greater benefits. |
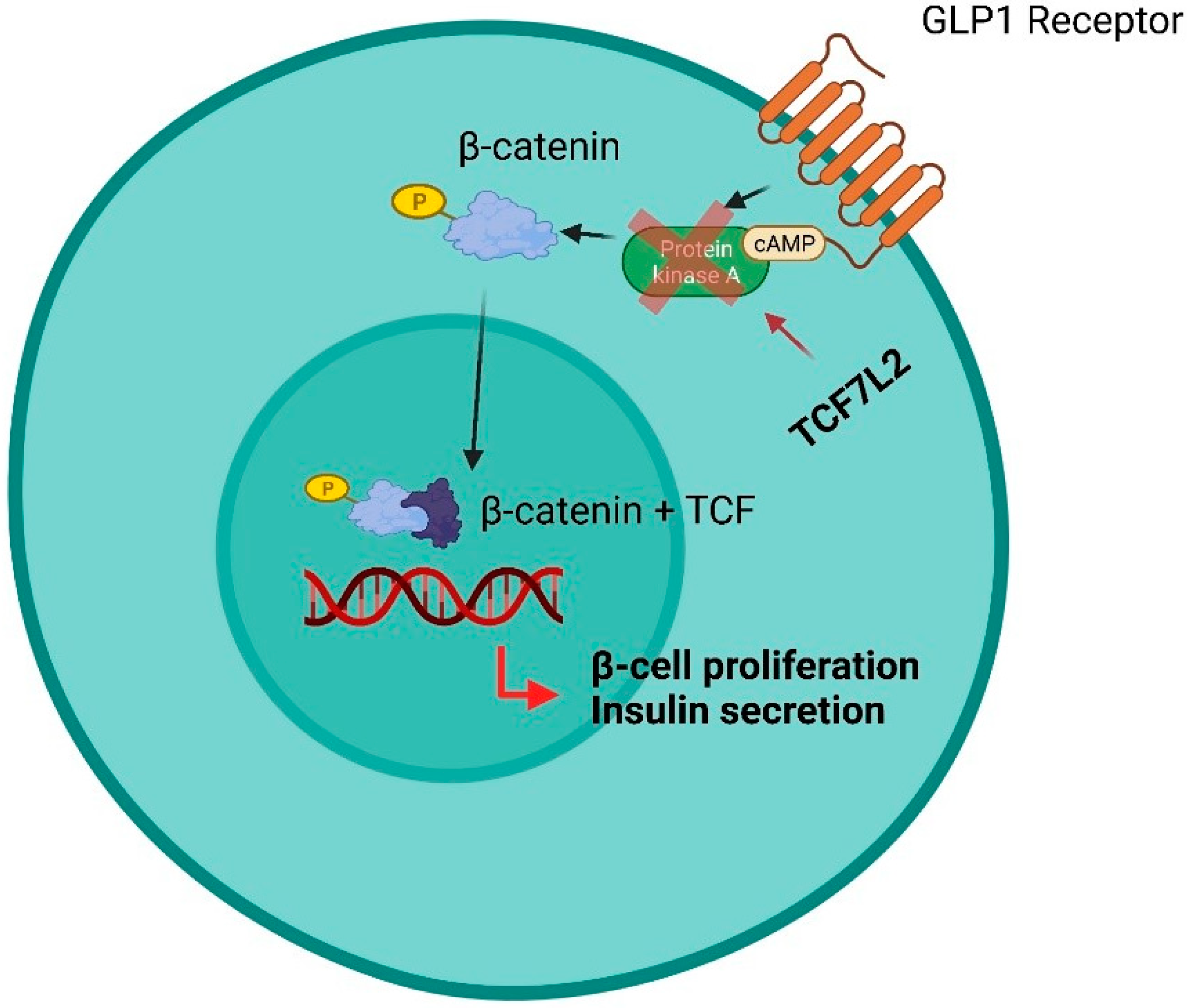
| TCF7L2 [132] | Gene | Single nucleotide polymorphisms (SNPs) | Predictive | Pancreatic response to incretins | CKD | Some variants of the TCF7L2 gene cause a lower pancreatic response to incretins. Therefore, patients carrying these variants are expected to benefit less from therapy with GLP-1 agonists or DDP-4 inhibitors. |
| miR-192 [137] | miRNAs | Non-coding RNA fragments | Predictive | Onset of CKD | CKD | Inhibition of miR-192 reduces the renal complications of diabetes. |
| 3-methyl-indole indicant [70] | Protein-bound uremic toxins | Microbiomics | Predictive | Severity of inflammation | CKD | Reduction of urinary levels of both markers after treatment is associated with reduction in inflammatory patterns in CKD patients. |
| Animal studies | ||||||
| Thymosin β4 [81,82] | Protein | Renal parenchyma | Prognostic | Sclerosis progression | Segmental glomerulosclerosis (FSGS) | Thymosin β4 was associated with sclerosis progression in animal models of FSGS. |
| miR-143 miR-145 [115] | miRNAs | Non-coding RNA fragments | Prognostic | Severity of CV damage | CKD | They are associated with higher severity and less stability of atherosclerotic plaque in CKD. |
| miR-21 [135] | miRNAs | Non-coding RNA fragments | Predictive | Severity of CKD | CKD | Inhibition of miR-21 reduces renal fibrosis in Alport nephropathy. |
| miR-145 [60] | miRNAs | Non-coding RNA fragments | Predictive | Severity of CV damage | CKD | Inhibition of miR-145 allows stabilization of the atherosclerotic plaque and the onset of CV events. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Provenzano, M.; Serra, R.; Garofalo, C.; Michael, A.; Crugliano, G.; Battaglia, Y.; Ielapi, N.; Bracale, U.M.; Faga, T.; Capitoli, G.; et al. OMICS in Chronic Kidney Disease: Focus on Prognosis and Prediction. Int. J. Mol. Sci. 2022, 23, 336. https://doi.org/10.3390/ijms23010336
Provenzano M, Serra R, Garofalo C, Michael A, Crugliano G, Battaglia Y, Ielapi N, Bracale UM, Faga T, Capitoli G, et al. OMICS in Chronic Kidney Disease: Focus on Prognosis and Prediction. International Journal of Molecular Sciences. 2022; 23(1):336. https://doi.org/10.3390/ijms23010336
Chicago/Turabian StyleProvenzano, Michele, Raffaele Serra, Carlo Garofalo, Ashour Michael, Giuseppina Crugliano, Yuri Battaglia, Nicola Ielapi, Umberto Marcello Bracale, Teresa Faga, Giulia Capitoli, and et al. 2022. "OMICS in Chronic Kidney Disease: Focus on Prognosis and Prediction" International Journal of Molecular Sciences 23, no. 1: 336. https://doi.org/10.3390/ijms23010336
APA StyleProvenzano, M., Serra, R., Garofalo, C., Michael, A., Crugliano, G., Battaglia, Y., Ielapi, N., Bracale, U. M., Faga, T., Capitoli, G., Galimberti, S., & Andreucci, M. (2022). OMICS in Chronic Kidney Disease: Focus on Prognosis and Prediction. International Journal of Molecular Sciences, 23(1), 336. https://doi.org/10.3390/ijms23010336