
Improving the Thermostability of a Fungal GH11 Xylanase via Fusion of a Submodule (C2) from Hyperthermophilic CBM9_1-2

Abstract
:
1. Introduction
2. Results
2.1. The Fusion Gene and the Chimeric CDBFV Variants
2.2. Enzyme Properties
2.2.1. Effects of the C2 Submodule on Enzyme Activity and Kinetic Characterization
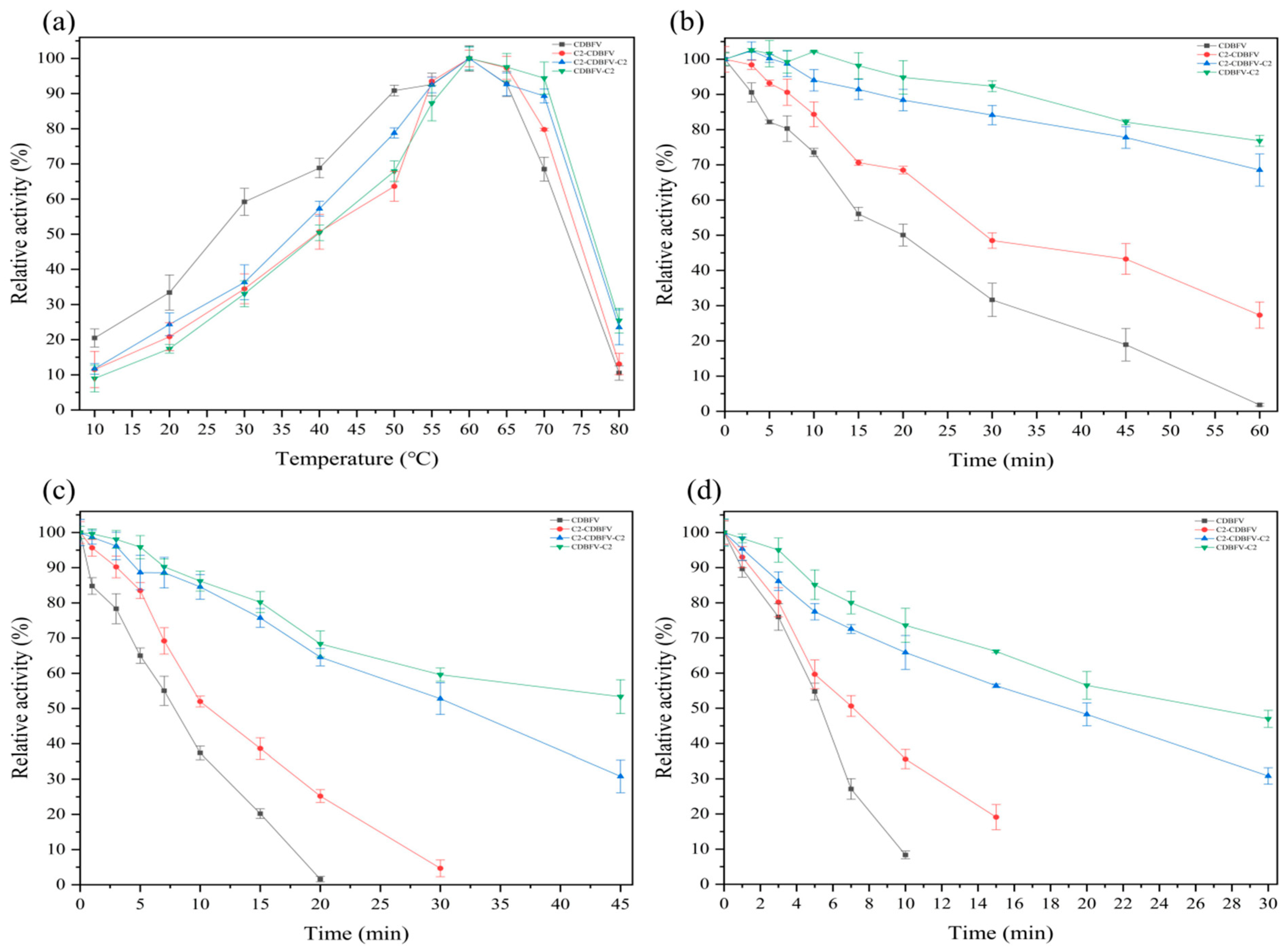
2.2.2. Effects of the C2 Submodule on pH and Temperature Characterization
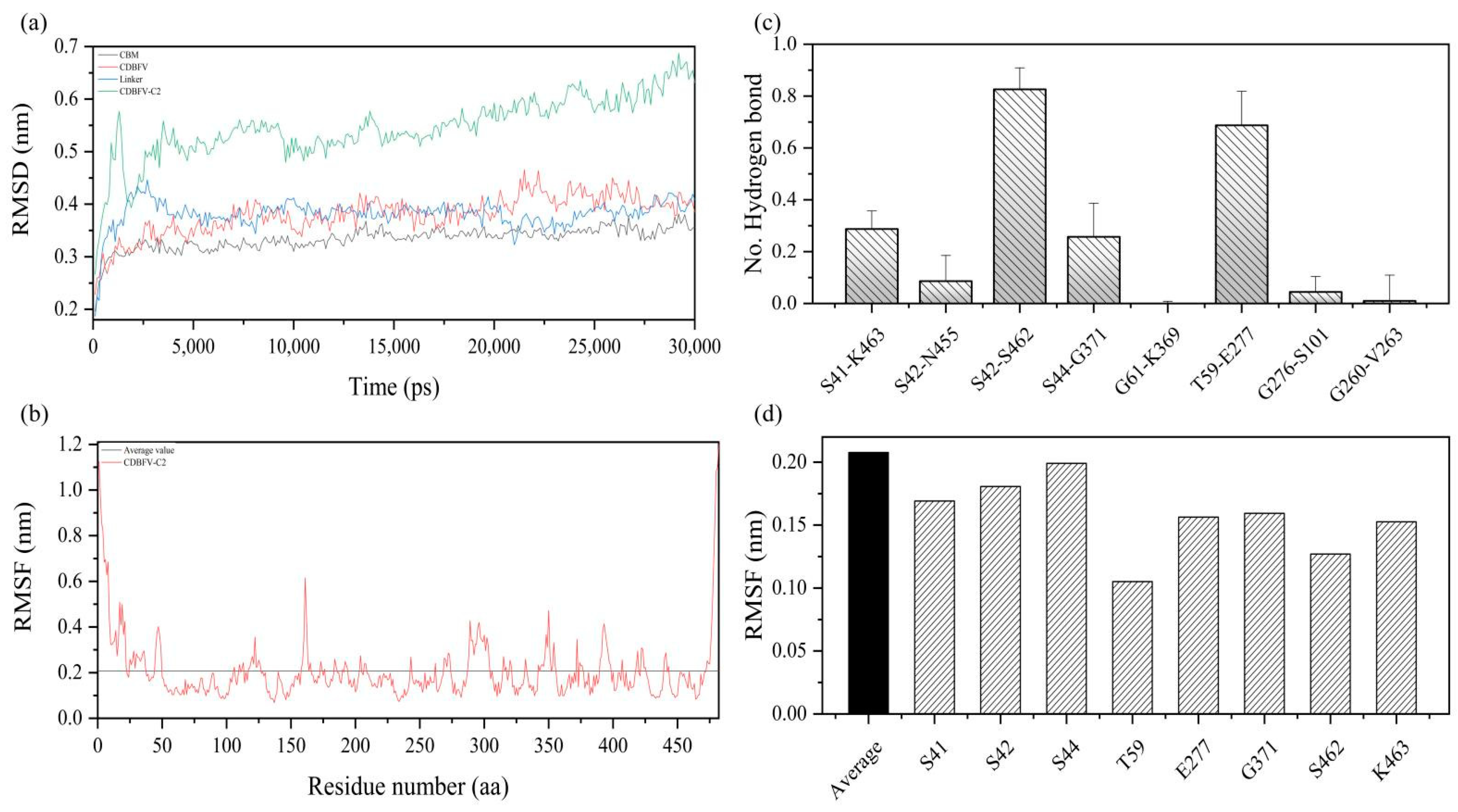
2.3. Molecular Modeling and Structural Analysis
2.4. MD Simulation Details of the Chimeric CDBFV-C2
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Vectors, and Reagents
4.2. Construction, Expression, and Purification of the Chimeric CDBFV Variants
4.3. Expression and Purification of the Chimeric CDBFV Variants
4.4. Assay Properties of the Chimeric CDBFV Variants
4.5. Molecular Modeling and Structural Analysis
4.6. MD Simulation Details of the Chimeric CDBFV-C2
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, L.W.; Zeng, L.Y.; Wang, S.Y.; Cheng, J.; Li, X.Q.; Song, A.D.; Wu, K.; Chen, H.G. Activity and thermostability increase of xylanase following transplantation with modules sub-divided from hyper-thermophilic CBM9_1-2. Process. Biochem. 2012, 47, 853–857. [Google Scholar] [CrossRef]
- Collins, T.; Charles, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Yu, B.; Zhang, K.Y.; Ding, X.M.; Chen, D.W. Thermostable carbohydrate binding module increases the thermostability and substrate-binding capacity of Trichoderma reesei xylanase 2. New Biotechnol. 2009, 26, 53–59. [Google Scholar] [CrossRef]
- Yang, H.Q.; Liu, L.; Li, J.H.; Du, G.C. Rational design to improve protein thermostability: Recent advances and prospects. ChemBioEng Rev. 2015, 2, 87–94. [Google Scholar] [CrossRef]
- Sunna, A.; Bergquist, P.L. A gene encoding a novel extremely thermosTable 1,4-beta-xylanase isolated directly from an environmental DNA sample. Extremophiles 2003, 7, 63–70. [Google Scholar] [CrossRef]
- Flint, J.E.; Lewis, R.J.; Lakey, J.H.; Morland, C.; Luginbühl, P.; Healey, S.; Todaro, T.; DeSantis, G.; Sun, M.; Parra-Gessert, L.; et al. Engineering hyperthermostability into a GH11 xylanase is mediated by subtle changes to protein structure. J. Biol. Chem. 2008, 283, 22557–22564. [Google Scholar] [CrossRef] [Green Version]
- Berrin, J.G.; Juge, N. Factors affecting xylanase functionality in the degradation of arabinoxylans. Biotechnol. Lett. 2008, 30, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Polizeli, M.L.T.M.; Rizzatti, A.C.S.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biot. 2005, 67, 577–591. [Google Scholar] [CrossRef]
- Pas, G.; Berrin, J.G.; Beaugrand, J. GH11 xylanases: Structure/function/properties relationships and applications. Biotechnol. Adv. 2011, 30, 564–592. [Google Scholar] [CrossRef]
- Wang, X.Y.; Luo, H.Y.; Yu, W.N.; Ma, R.; You, S.A.; Liu, W.N.; Hou, L.Y.; Zheng, F.; Xie, X.M.; Yao, B. A thermostable gloeophyllum trabeum xylanase with potential for the brewing industry. Food. Chem. 2016, 199, 516–523. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The carbohydrate-active enzymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids. Res. 2009, 37, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Mamo, G.; Hatti-Kaul, R.; Mattiasson, B. Fusion of carbohydrate binding modules from Thermotoga neapolitana with a family 10 xylanase from Bacillus halodurans S7. Extremophiles 2007, 11, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, M.; Khan, M.I.M.; Zafar, R.; Ahmad, S.; Niazi, U.H.K.; Akhtar, M.W. Influence of positioning of carbohydrate binding module on the activity of endoglucanase CelA of Clostridium thermocellum. J. Biotechnol. 2012, 161, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, M.; Khan, M.I.M.; Akbar, N.S.; Ahmad, S.; Ali, I.; Akhtar, M.W. Enhanced expression and activity yields of Clostridium thermocellum xylanases without non-catalytic domains. J. Biotechnol. 2010, 145, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.D.; Ying, Y.; Chen, X.H.; Lu, M.; Ning, K.; Wang, L.S.; Li, F.L. Distinct roles for carbohydrate-binding modules of glycoside hydrolase 10 (GH10) and GH11 xylanases from Caldicellulosiruptor sp. strain F32 in thermostability and catalytic efficiency. Appl. Environ. Microbiol. 2015, 81, 2006–2014. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.W.; Cheng, J.; Chen, H.G.; Li, X.Q.; Wang, S.Y.; Song, A.; Wang, M.D.; Wang, B.; Shen, J.W. Directed evolution of a mesophilic fungal xylanase by fusion of a thermophilic bacterial carbohydrate-binding module. Process. Biochem. 2011, 46, 395–398. [Google Scholar] [CrossRef]
- Sunna, A.; Gibbs, M.D.; Bergquist, P.L. The thermostabilizing domain, XynA, of Caldibacillus cellulovorans xylanase is a xylan binding domain. Biochem. J. 2000, 346, 583–586. [Google Scholar] [CrossRef]
- Han, N.Y.; Miao, H.B.; Ding, J.M.; Li, J.J.; Mu, Y.L.; Zhou, J.P.; Huang, Z.X. Improving the thermostability of a fungal GH11 xylanase via site-directed mutagenesis guided by sequence and structural analysis. Biotechnol. Biofuels. 2017, 10, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.E.; Lowe, S.E.; Henrissat, B.; Zeikus, J.G. Characterization of the active site and thermostability regions of endoxylanase from Thermoanaerobacterium saccharolyticum B6A-RI. J. Bacteriol. 1993, 175, 5890–5898. [Google Scholar] [CrossRef] [Green Version]
- Fontes, C.M.G.A.; Hazlewood, G.P.; Morag, E.; Hall, J.; Hirst, B.H.; Gilbert, H.J. Evidence for a general role for non-catalytic thermostabilizing domains in xylanases from thermophilic bacteria. Biochem. J. 1995, 307, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Takagi, K.I.; Fukumura, M.; Kimura, T.; Karita, S.; Sakka, K.; Ohmiya, K. Sequence of xynC and properties of XynC, a major component of the Clostridium thermocellum cellulosome. J. Bacteriol. 1997, 179, 4246–4253. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Qin, Z.; Chen, Q.M.; Fan, L.Q.; Zhou, J.C.; Zhao, L.M. Efficient immobilization of bacterial GH family 46 chitosanase by carbohydrate-binding module fusion for the controllable preparation of chitooligosaccharides. J. Agric. Food. Chem. 2019, 67, 6847–6855. [Google Scholar] [CrossRef]
- Kim, D.R.; Lim, H.K.; Lee, K.I.; Hwang, I.T. Identification of a novel cellulose-binding domain within the endo-β-1,4-xylanase KRICT PX-3 from Paenibacillus terrae HPL-003. Enzyme. Microb. Technol. 2016, 93, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein. Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Notenboom, V.; Boraston, A.B.; Kilburn, D.G.; Rose, D.R. Crystal structures of the family 9 carbohydrate-binding module from Thermotoga maritima xylanase 10A in native and ligand-bound forms. Biochemistry 2001, 40, 6248–6256. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.S.; Chen, C.C.; Huang, C.H.; Ko, T.P.; Luo, W.H.; Huang, J.W.; Liu, J.R.; Guo, R.T. Structural analysis of a glycoside hydrolase family 11 xylanase from Neocallimastix patriciarum. J. Biol. Chem. 2014, 289, 11020–11028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, P.; Feng, M.G. Bifunctional enhancement of a beta-glucanase-xylanase fusion enzyme by optimization of peptide linkers. Appl. Microbiol. Biotechnol. 2008, 79, 579–587. [Google Scholar] [CrossRef]
- An, J.M.; Kim, Y.K.; Lim, W.J.; Hong, S.Y.; An, C.L.; Shin, E.C.; Cho, K.M.; Choi, B.R.; Kang, J.M.; Lee, S.M.; et al. Evaluation of a novel bifunctional xylanase-cellulase constructed by gene fusion. Enzyme. Microb. Technol. 2005, 36, 989–995. [Google Scholar] [CrossRef]
- Lu, P.; Feng, M.G.; Li, W.F.; Hu, C.X. Construction and characterization of a bifunctional fusion enzyme of Bacillus-sourced β-glucanase and xylanase expressed in Escherichia coli. FEMS Microbiol. Lett. 2006, 261, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pell, G.; Szabo, L.; Charnock, S.J.; Xie, H.F.; Gloster, T.M.; Davies, G.J.; Gilbert, H.J. Structural and biochemical analysis of Cellvibrio japonicus xylanase 10C: How variation in substrate-binding cleft influences the catalytic profile of family GH-10 xylanases. J. Biol. Chem. 2004, 279, 11777–11788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, D.K.Y.; Withers, S.G.; Mcintosh, L.P. Direct demonstration of the flexibility of the glycosylated prolinethreonine linker in the Cellulomonas fimi xylanase cex through NMR spectroscopic analysis. J. Biol. Chem. 2007, 82, 2091–2100. [Google Scholar] [CrossRef] [Green Version]
- Tajwar, R.; Shahid, S.; Zafar, R.; Akhtar, M.W. Impact of orientation of carbohydrate binding modules family 22 and 6 on the catalytic activity of Thermotoga maritima xylanase XynB. Enzyme. Microb. Technol. 2017, 106, 295–300. [Google Scholar] [CrossRef]
- Suma, K.; Podile, A.R. Chitinase A from Stenotrophomonas maltophilia shows transglycosylation and antifungal activities. Bioresour. Technol. 2013, 133, 213–220. [Google Scholar] [CrossRef]
- Yang, J.Y.; Anishchenko, I.; Park, H.; Peng, Z.; Ovchinnikov, S.; Baker, D. Improved protein structure prediction using predicted interresidue orientations. Proc. Natl. Acad. Sci. USA 2020, 117, 14677–14685. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- Willard, L.; Ranjan, A.; Zhang, H.Y.; Monzavi, H.; Boyko, R.F.; Sykes, B.D.; Wishart, D.S. VADAR: A web server for quantitative evaluation of protein structure quality. Nucleic Acids. Res. 2003, 31, 3316–3319. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. The PyMOL Molecular Graphics System, Version 2.0. Schrodinger, LLC. Available online: http://www.pymol.org (accessed on 1 November 2021).
- Hess, B.; Kutzner, C.; van der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory. Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornak, V.; Abel, R.; Okur, A.; Strockbine, B.; Roitberg, A.; Simmerling, C. Comparison of multiple amber force fields and development of improved protein backbone parameters. Proteins 2006, 65, 712–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald-an n.Log (N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Xylanase | Number of Residues | Mol. Mass a (kDa) | Vmaxb (μmol/min/mg) | KMb (mg/mL) | t1/2 (60 °C/ 65 °C/70 °C) c (min) |
|---|---|---|---|---|---|
| CDBFV | 224 | 29.4/24.3 | 1960 ± 100 | 8.0 ± 0.7 | 24.8/8.7/5.3 |
| C2-CDBFV | 435 | 53.0/47.9 | 2210 ± 80 | 7.2 ± 0.5 | 37.5/13.7/8.1 |
| CDBFV-C2 | 435 | 53.0/47.9 | 2220 ± 160 | 7.1 ± 0.9 | 122.2/44.2/25.3 |
| C2-CDBFV-C2 | 646 | 76.5/71.4 | 2340 ± 120 | 6.9 ± 0.7 | 93.1/31.6/19.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, H.; Ma, Y.; Zhe, Y.; Tang, X.; Wu, Q.; Huang, Z.; Han, N. Improving the Thermostability of a Fungal GH11 Xylanase via Fusion of a Submodule (C2) from Hyperthermophilic CBM9_1-2. Int. J. Mol. Sci. 2022, 23, 463. https://doi.org/10.3390/ijms23010463
Miao H, Ma Y, Zhe Y, Tang X, Wu Q, Huang Z, Han N. Improving the Thermostability of a Fungal GH11 Xylanase via Fusion of a Submodule (C2) from Hyperthermophilic CBM9_1-2. International Journal of Molecular Sciences. 2022; 23(1):463. https://doi.org/10.3390/ijms23010463
Chicago/Turabian StyleMiao, Huabiao, Yu Ma, Yuanyuan Zhe, Xianghua Tang, Qian Wu, Zunxi Huang, and Nanyu Han. 2022. "Improving the Thermostability of a Fungal GH11 Xylanase via Fusion of a Submodule (C2) from Hyperthermophilic CBM9_1-2" International Journal of Molecular Sciences 23, no. 1: 463. https://doi.org/10.3390/ijms23010463
APA StyleMiao, H., Ma, Y., Zhe, Y., Tang, X., Wu, Q., Huang, Z., & Han, N. (2022). Improving the Thermostability of a Fungal GH11 Xylanase via Fusion of a Submodule (C2) from Hyperthermophilic CBM9_1-2. International Journal of Molecular Sciences, 23(1), 463. https://doi.org/10.3390/ijms23010463