
Molecular Basis to Integrate Microgravity Signals into the Photoperiodic Flowering Pathway in Arabidopsis thaliana under Spaceflight Condition

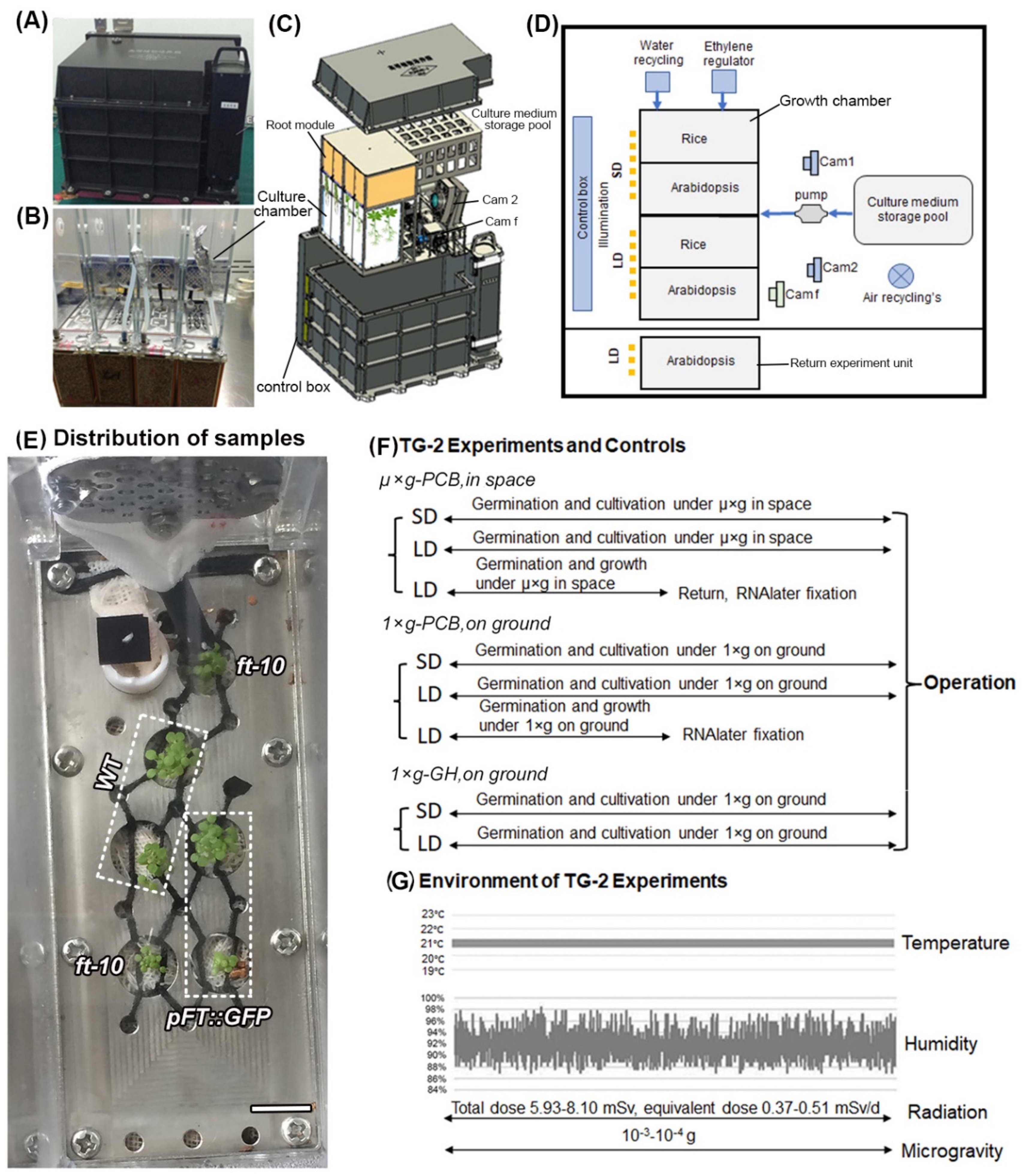{kind=link}
{kind=link}
{kind=link}
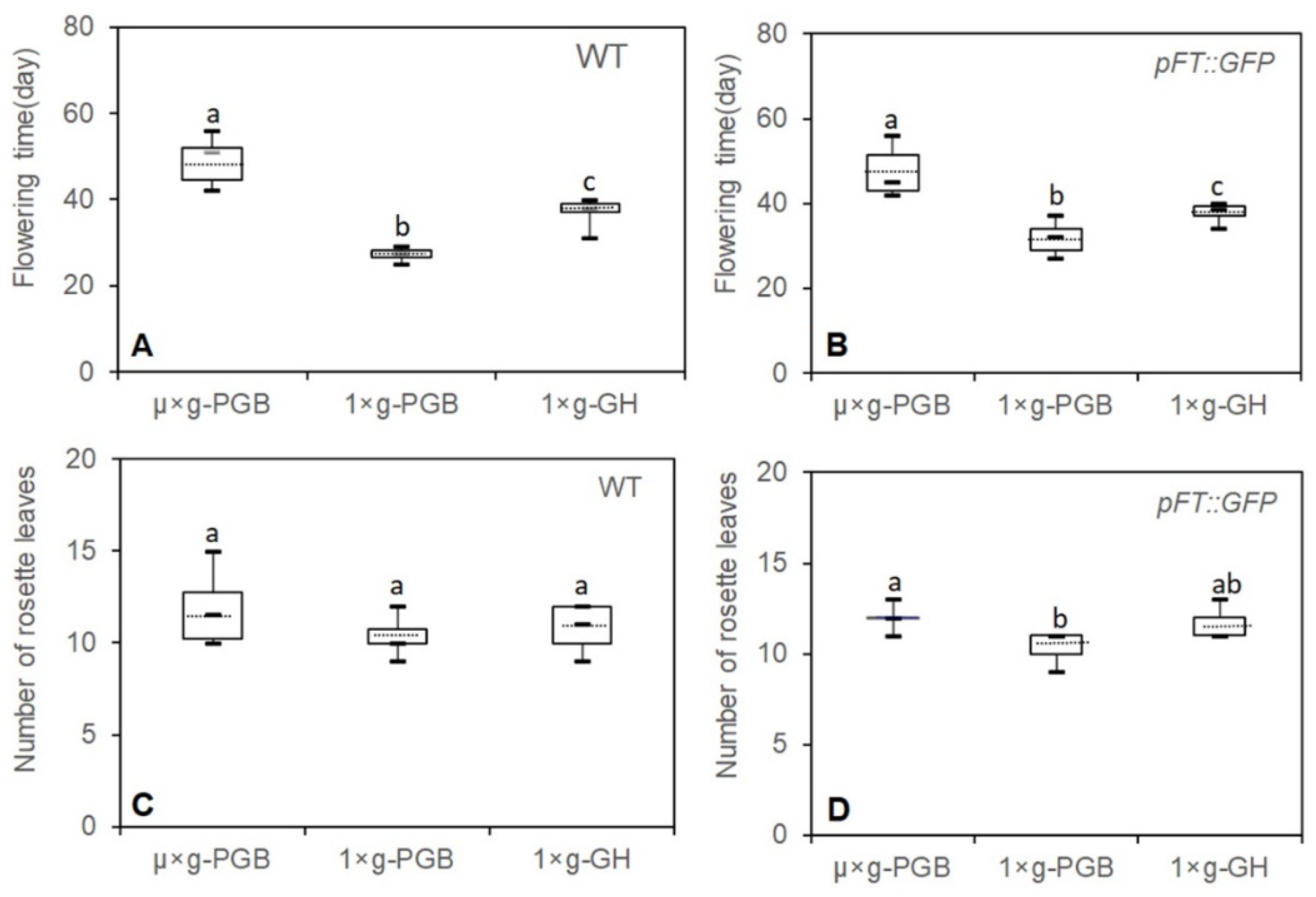{kind=link}
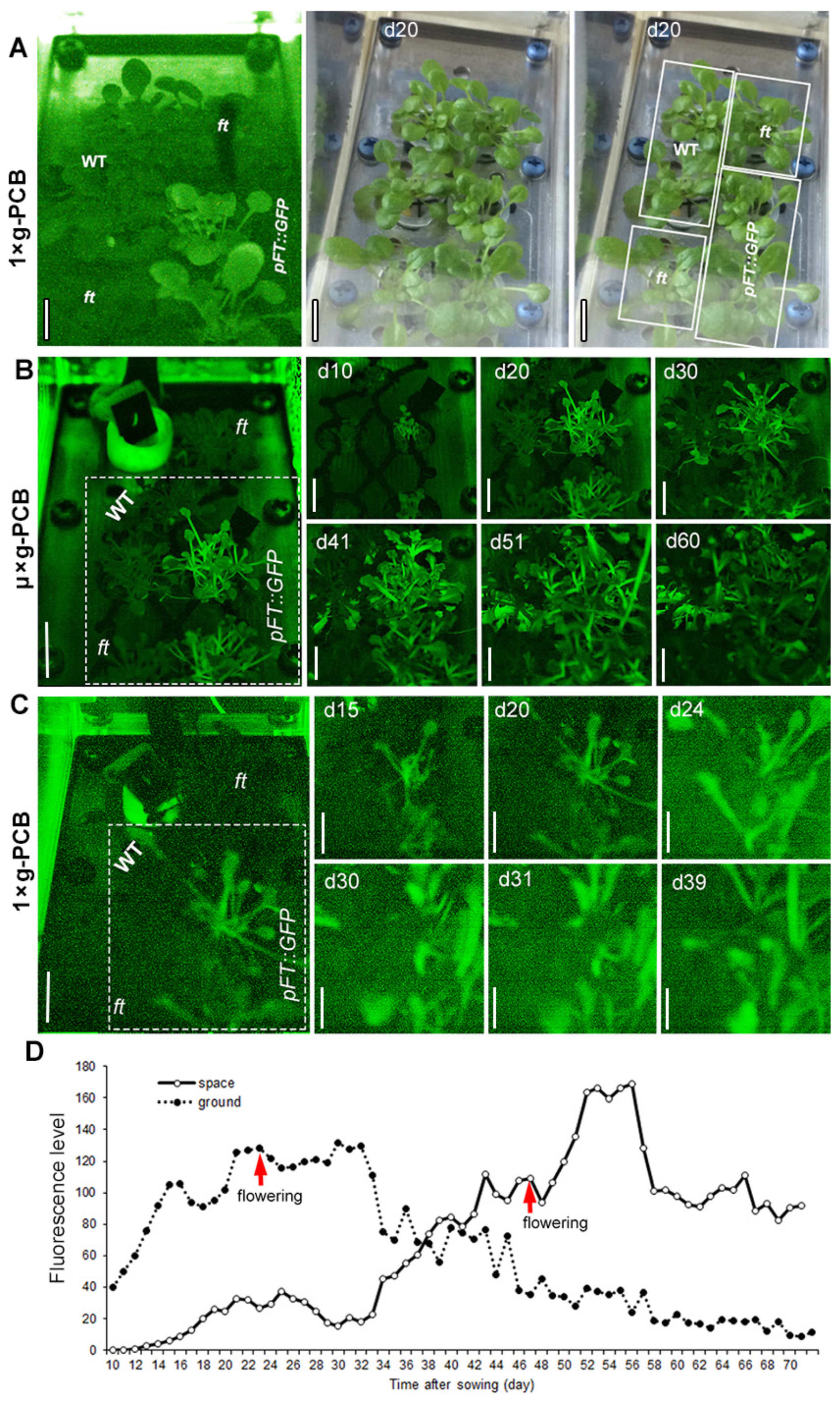{kind=link}
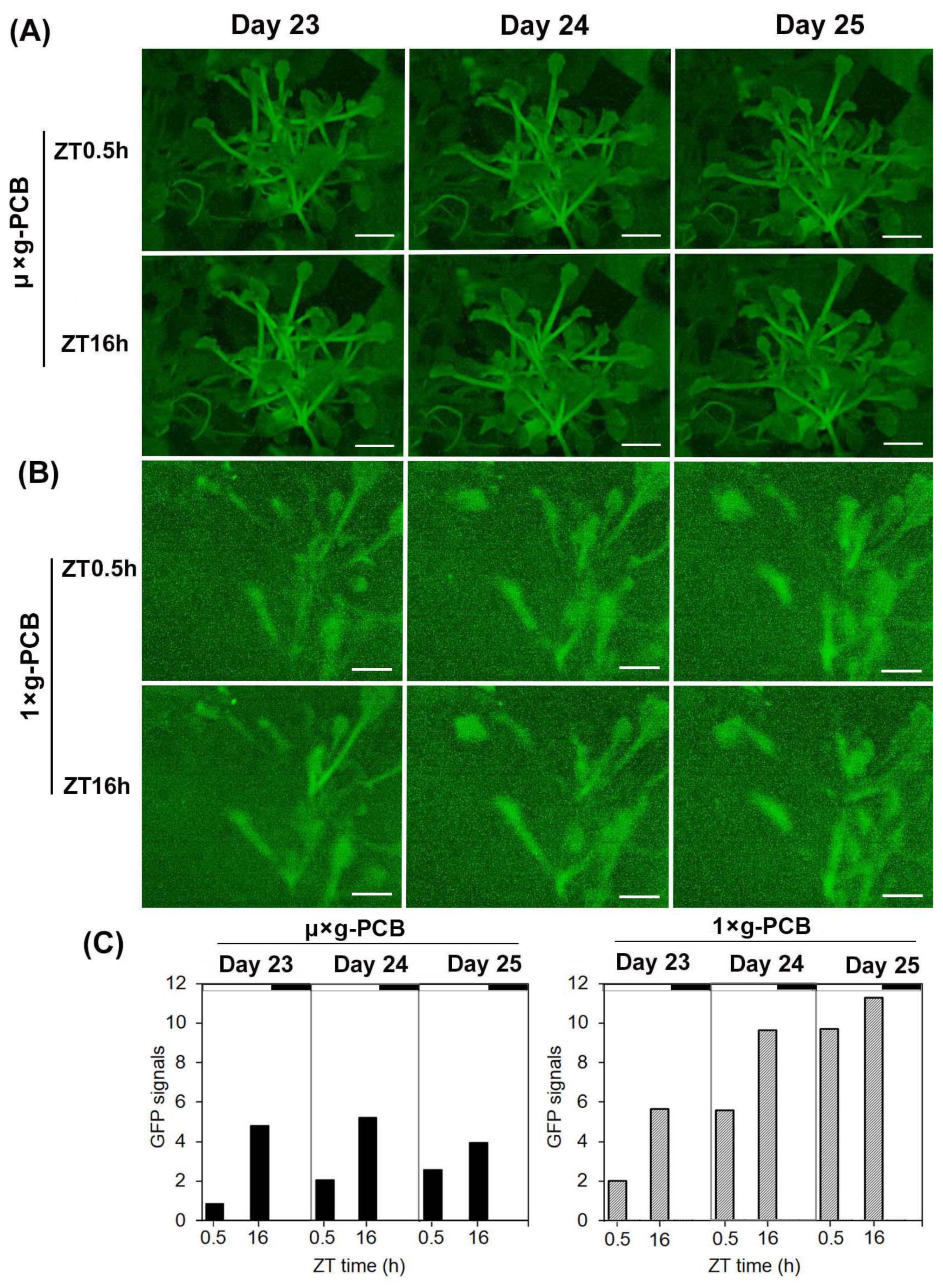{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Microgravity on Flowering of Plants under Different Photoperiodic Conditions
2.2. Application of GFP Technique for Detection of FT Expression in Space
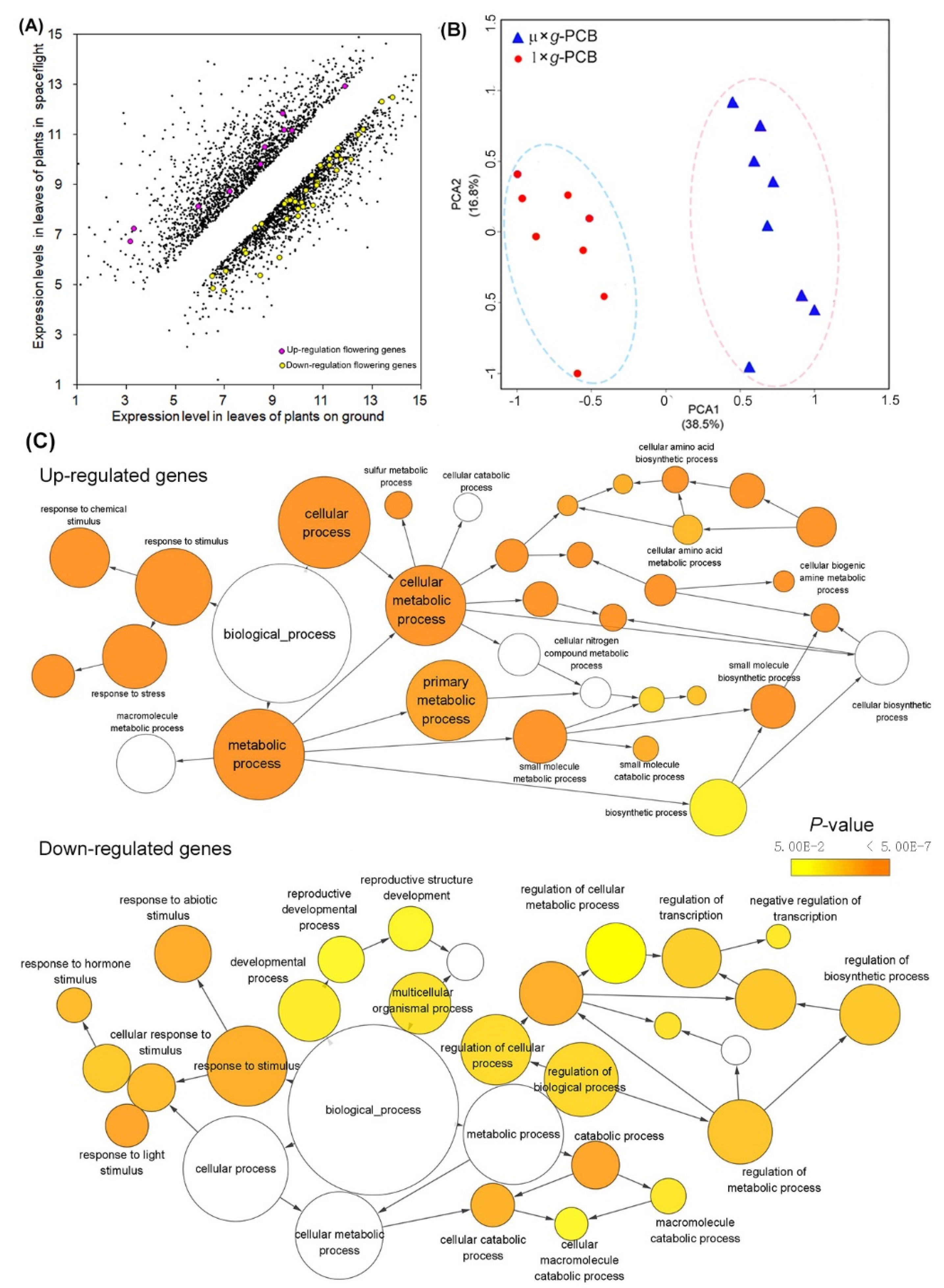
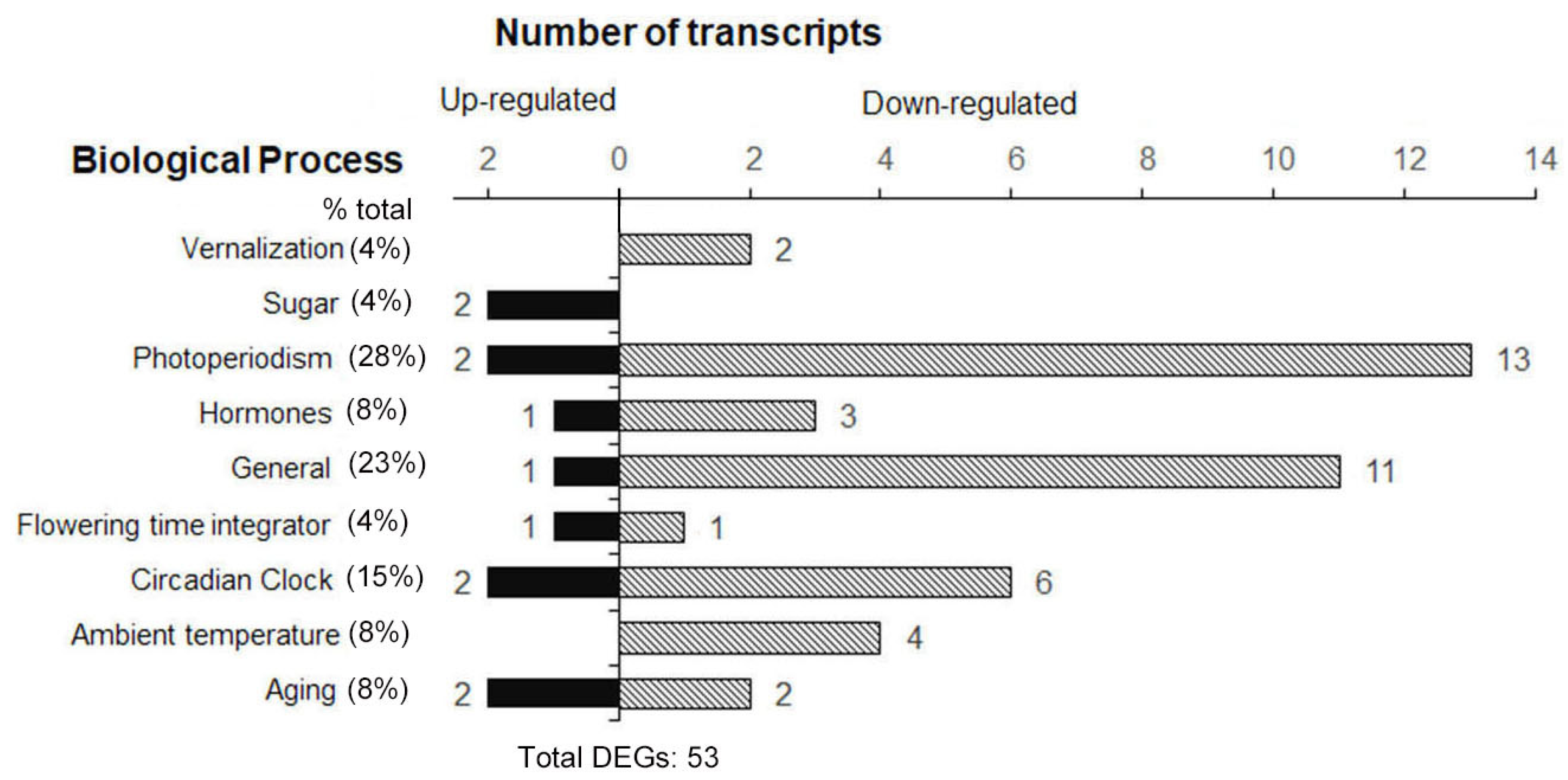
2.3. Altered Expression of Genes in Arabidopsis Leaves at Flowering Stage in Space
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Spaceflight Procedures
4.3. Measurement of Flowering Time and Fluorescence Intensity
4.4. RNA Extraction and Microarray Analysis
4.5. Analysis of Microarray Data
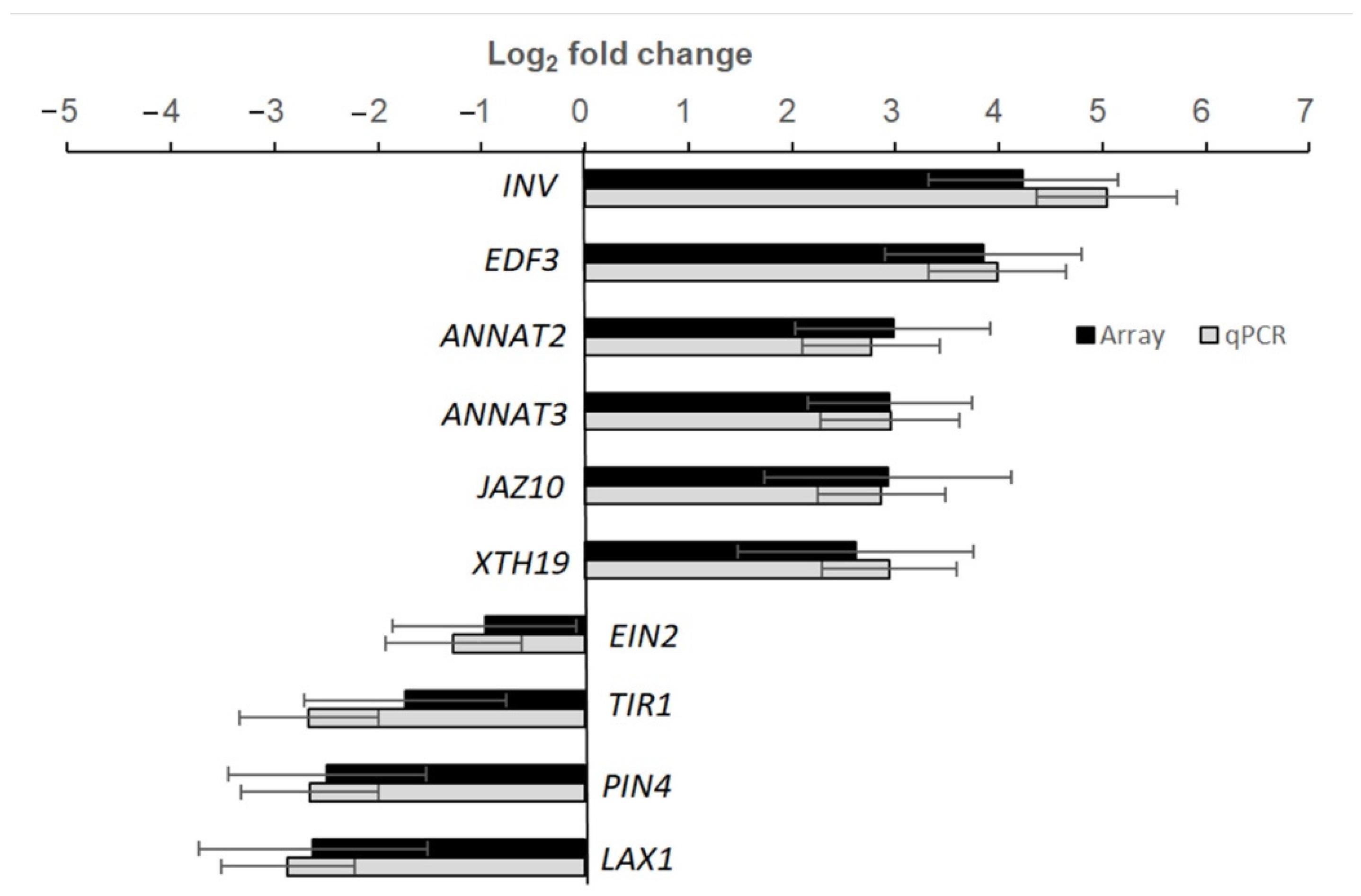
4.6. Real-Time qRT-PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gitelson, J.I. Biological life-support systems for Mars mission. Adv. Space Res. 1992, 12, 167–192. [Google Scholar] [CrossRef]
- De Micco, V.; De Pascale, S.; Paradiso, R.; Aronne, G. Microgravity effects on different stages of higher plant life cycle and completion of the seed-to-seed cycle. Plant Biol. 2014, 16, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.R. Isolation and integrated testing: An introduction to the Lunar-Mars life Support Test Project. In Isotion-NASA Experiments in Closed Environment Living; Science and Technology Series; Lane, H.W., Sauer, R.L., Feeback, D.L., Eds.; Univelt Inc.: San Diego, CA, USA, 2002; pp. 1–6. [Google Scholar]
- Park, H.J.; Kim, W.Y.; Pardo, J.M.; Yun, D.J. Molecular interactions between flowering time and abiotic stress pathways. Int. Rev. Cell Mol. Biol. 2016, 327, 371–412. [Google Scholar]
- Olas, J.J.; Van Dingenen, J.; Abel, C.; Dzialo, M.A.; Feil, R.; Krapp, A.; Schlereth, A.; Wahl, V. Nitrate acts at the Arabidopsis thaliana shoot apical meristem to regulate flowering time. New Phytol. 2019, 223, 814–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunkar, R.; Chinnusamy, V.; Zhu, J.; Zhu, J.K. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 2007, 12, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.C.; Takeno, K. Stress-induced flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef]
- Wolff, S.A.; Coelho, L.H.; Zabrodina, M.; Brinckmann, E.; Kittang, A.I. Plant mineral nutrition, gas exchange and photosynthesis in space: A review. Adv. Space Res. 2013, 51, 465–475. [Google Scholar] [CrossRef]
- Wang, L.; Han, F.; Zheng, H. Photoperiod-controlling guttation and growth of rice seedlings under microgravity on board Chinese spacelab TG-2. Microgravity Sci. Technol. 2018, 30, 839–847. [Google Scholar] [CrossRef]
- Halstead, T.W.; Dutcher, F.R. Plants in space. Annu. Rev. Plant Physiol. 1987, 38, 317–345. [Google Scholar] [CrossRef]
- Nechitailo, G.S.; Maskinsky, A.L. Space Biology: Studies at Orbital Station; Mir Publishers: Moscow, Russia, 1993; pp. 293–330. [Google Scholar]
- Strickland, D.T.; Campbell, W.F.; Salisbury, F.B.; Bingham, G.E. Morphological assessment of reproductive structures of wheat grown on Mir. Gravit. Space Biol. Bull. 1997, 11, 14. [Google Scholar]
- Xie, J.; Zheng, H. Arabidopsis flowering induced by photoperiod under 3-D clinostat rotational simulated microgravity. Acta Astronaut. 2020, 166, 567–572. [Google Scholar] [CrossRef]
- Kordyum, E.L.; Sytnik, K.M.; Chernyaeva, I.I. Peculiarities of genital organ formation in Arabidopsis thaliana (L) Heynh. under spaceflight conditions. Adv. Space Res. 1983, 3, 247–250. [Google Scholar] [CrossRef]
- Kuang, A.; Xiao, Y.; McClure, G.; Musgrave, M.E. Influence of microgravity on ultrastructure and storage reserves in seeds of Brassica rapa L. Ann. Bot. 2000, 85, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Campbell, F.; Salisbury, B.; Bugbee, B.; Klassen, S.; Naegle, E.; Strickland, T.; Bingham, E.; Levinkikn, M.; Iljina, M.; Sytchev, N.; et al. Comparative floral development of Mir-grown and ethylene-treated, Earth-grown Super Dwarf wheat. J. Plant Physiol. 2001, 158, 1051–1060. [Google Scholar] [CrossRef] [Green Version]
- Link, B.M.; Durst, S.J.; Zhou, W.; Stankovic, B. Seed-to-seed growth of Arabidopsis thaliana on the international space station. Space Life Sciences: Gravity-Related Processes in Plants. Adv. Space Res. 2003, 31, 2237–2243. [Google Scholar] [CrossRef]
- Zheng, H. Flowering in space. Microgravity Sci. Technol. 2018, 30, 783–791. [Google Scholar] [CrossRef]
- Karahara, I.; Suto, T.; Yamaguchi, T.; Yashiro, U.; Tamaoki, D.; Okamoto, E.; Yano, S.; Tanigaki, F.; Shimazu, T.; Kasahara, H.; et al. Vegetative and reproductive growth of Arabidopsis under microgravity conditions in space. J. Plant Res. 2020, 133, 571–585. [Google Scholar] [CrossRef]
- Musgrave, M.E.; Kuang, A. Plant reproductive development during spaceflight. In Developmental Biology Research in Space; Marthy, H.-J., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2003; pp. 1–23. [Google Scholar]
- Putterill, J.; Laurie, R.; Macknight, R. It’s time to flower: The genetic control of flowering time. Bioessays 2004, 26, 363–373. [Google Scholar] [CrossRef]
- Salazar, J.D.; Saithong, T.; Brown, P.E.; Foreman, J.; Locke, J.C.; Halliday, K.J.; Carré, I.A.; Rand, D.A.; Millar, A.J. Prediction of photoperiodic regulators from quantitative gene circuit models. Cell 2009, 139, 1170–1179. [Google Scholar] [CrossRef] [Green Version]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef]
- Paul, A.L.; Wheeler, R.M.; Levine, H.G.; Ferl, R.J. Fundamental plant biology enabled by the space shuttle. J. Am. Bot. 2013, 100, 226–234. [Google Scholar] [CrossRef]
- Zheng, H.; Han, F.; Le, J. Higher plants in space: Microgravity perception, response, and adaptation. Microgravity Sci. Technol. 2015, 27, 377–386. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, H.; Wei, N.; Chen, A.; Wang, L.; Zheng, W.; Zhang, T. Live imaging technique for studies of growth and development of Chinese cabbage under microgravity in a recoverable satellite (SJ-8). Microgravity Sci. Technol. 2008, 20, 137–143. [Google Scholar] [CrossRef]
- Krzymuski, M.; Andrés, F.; Cagnola, J.I.; Jang, S.; Yanovsky, M.J.; Coupland, G.; Casal, J.J. The dynamics of FLOWERING LOCUS T expression encodes long-day information. Plant J. 2015, 83, 952–961. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [Green Version]
- Bouché, F.; Lobert, G.; Tocquin, P.; Périlleux, C. FLOR-ID: An interactive database of flowering- time gene networks in Arabidopsis thaliana. Nucleic Acids Res. 2016, 44, 1167–1171. [Google Scholar] [CrossRef] [Green Version]
- Gocal, G.F.; Sheldon, C.C.; Gubler, F.; Moritz, T.; Bagnall, D.J.; MacMillan, C.P.; Li, S.F.; Parish, R.W.; Dennis, E.S.; Weigel, D.; et al. GAMYB-like genes, flowering, and gibberellin signaling in Arabidopsis. Plant Physiol. 2001, 127, 1682–1693. [Google Scholar] [CrossRef]
- Millar, A.A.; Gubler, F. The Arabidopsis GAMYB-like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Škiljaica, A.; Lechner, E.; Jagić, M.; Majsec, K.; Malenica, N.; Genschik, P.; Bauer, N. The protein turnover of Arabidopsis BPM1 is involved in regulation of flowering time and abiotic stress response. Plant Mol. Biol. 2020, 102, 359–372. [Google Scholar] [CrossRef]
- Yanagisawa, S. The Dof family of plant transcription factors. Trends Plant Sci. 2002, 7, 555–560. [Google Scholar] [CrossRef]
- Merkys, A.I.; Laurinavicius, R.S. Complete cycle of individual development of Arabidopsis thaliana (L.) Heynh. plants on board the Salyut-7 orbital station. Dokl. Bot. Sci.-Akad. Nauk. SSSR 1983, 271, 512–557. [Google Scholar]
- Link, B.M.; Busse, J.; Stankovic, B. Seed-to-seed-to-seed growth and development of Arabidopsis in microgravity. Astrobiology 2014, 14, 866–875. [Google Scholar] [CrossRef] [Green Version]
- Musgrave, M.E.; Kuang, A.; Tuominen, L.K.; Levine, L.H.; Morrow, R.C. Seed storage reserves and glucosinolates in Brassica rapa L. grown on the International Space Station. J. Am. Soc. Hortic. Sci. 2005, 130, 848–856. [Google Scholar] [CrossRef]
- Popova, A.F.; Musgrave, M.; Kuang, A. The development of embryos in Brassica rapa L. in microgravity. Cytol. Genet. 2009, 43, 89–93. [Google Scholar] [CrossRef]
- Yamamoto, K.; Takahashi, K.; Hara, M.; Miyata, K.; Hayama, R.; Mizoguchi, T. Density effects on late flowering mutants of Arabidopsis thaliana under continuous light. Plant Biotechnol. 2016, 33, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Nefissi, R.; Natsui, Y.; Miyata, K.; Oda, A.; Hase, Y.; Nakagawa, M.; Ghorbel, A.; Mizoguchi, T. Double loss-of-function mutation in EARLY FLOWERING3 and CRYPTOCHROME 2 genes delays flowering under continuous light but accelerates it under long days and short days: An important role for Arabidopsis CRY2 to accelerate flowering time in continuous light. J. Exp. Bot. 2011, 62, 2731–2744. [Google Scholar] [CrossRef] [Green Version]
- Miyata, K.; Calviño, M.; Oda, A.; Sugiyama, H.; Mizoguchi, T. Suppression of late-flowering and semi-dwarf phenotypes in the arabidopsis clock mutant lhy-12; cca1-101 by phyB under continuous light. Plant Signal. Behav. 2011, 6, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Porterfield, D.M.; Matthews, S.W.; Daugherty, C.J.; Musgrave, M.E. Spaceflight exposure effects on transcription, activity, and localization of alcohol dehydrogenase in the roots of Arabidopsis thaliana. Plant Physiol. 1997, 113, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.L.; Daugherty, C.J.; Bihn, E.A.; Chapman, D.K.; Norwood, K.L.; Ferl, R.J. Transgene expression patterns indicate that spaceflight affects stress signal perception and transduction in Arabidopsis. Plant Physiol. 2001, 126, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Ferl, R.J.; Koh, J.; Denison, F.; Paul, A.L. Spaceflight induces specific alterations in the proteomes of Arabidopsis. Astrobiology 2015, 15, 32–56. [Google Scholar] [CrossRef]
- Song, H.R.; Song, J.D.; Cho, J.N.; Amasino, R.M.; Noh, B.; Noh, Y.S. The RNA binding protein ELF9 directly reduces SUPPRESSOR OF OVEREXPRESSION OF CO1 transcript levels in arabidopsis, possibly via nonsense-mediated mRNA decay. Plant Cell 2009, 21, 1195–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanzawa, Y.; Money, T.; Bradley, D. A single amino acid converts a repressor to an activator of flowering. Proc. Natl. Acad. Sci. USA 2005, 102, 7748–7753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xie, J.; Wang, L.; Zheng, H. Circumnutation and growth of inflorexcence stems of Arabidopsis thaliana in response to microgravity under different photoperiod conditions. Life 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Huang, X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol Update for large-scale genome and gene function analysis with the PANTHER classification system (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef]
- Sun, H.; Wang, J.; Gong, Z.; Yao, J.; Wang, Y.; Xu, J.; Yuan, G.; Zhang, Y.; Shao, Z. Quantitative integration of epigenomic variation and transcription factor binding using MA motif toolkit identifies an important role of IRF2 as transcription activator at gene promoters. Cell Discov. 2018, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Ran, X.; Zhao, F.; Wang, Y.; Liu, J.; Zhuang, Y.; Ye, L.; Qi, M.; Cheng, J.; Zhang, Y. Plant regulomics: A data-driven interface for retrieving upstream regulators from plant multi-omics data. Plant J. 2020, 101, 237–248. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, J.; Wang, L.; Zheng, H. Molecular Basis to Integrate Microgravity Signals into the Photoperiodic Flowering Pathway in Arabidopsis thaliana under Spaceflight Condition. Int. J. Mol. Sci. 2022, 23, 63. https://doi.org/10.3390/ijms23010063
Xie J, Wang L, Zheng H. Molecular Basis to Integrate Microgravity Signals into the Photoperiodic Flowering Pathway in Arabidopsis thaliana under Spaceflight Condition. International Journal of Molecular Sciences. 2022; 23(1):63. https://doi.org/10.3390/ijms23010063
Chicago/Turabian StyleXie, Junyan, Lihua Wang, and Huiqiong Zheng. 2022. "Molecular Basis to Integrate Microgravity Signals into the Photoperiodic Flowering Pathway in Arabidopsis thaliana under Spaceflight Condition" International Journal of Molecular Sciences 23, no. 1: 63. https://doi.org/10.3390/ijms23010063
APA StyleXie, J., Wang, L., & Zheng, H. (2022). Molecular Basis to Integrate Microgravity Signals into the Photoperiodic Flowering Pathway in Arabidopsis thaliana under Spaceflight Condition. International Journal of Molecular Sciences, 23(1), 63. https://doi.org/10.3390/ijms23010063