
Reduced Environmental Dose Rates Are Responsible for the Increased Susceptibility to Radiation-Induced DNA Damage in Larval Neuroblasts of Drosophila Grown inside the LNGS Underground Laboratory



 ,
,  and
and 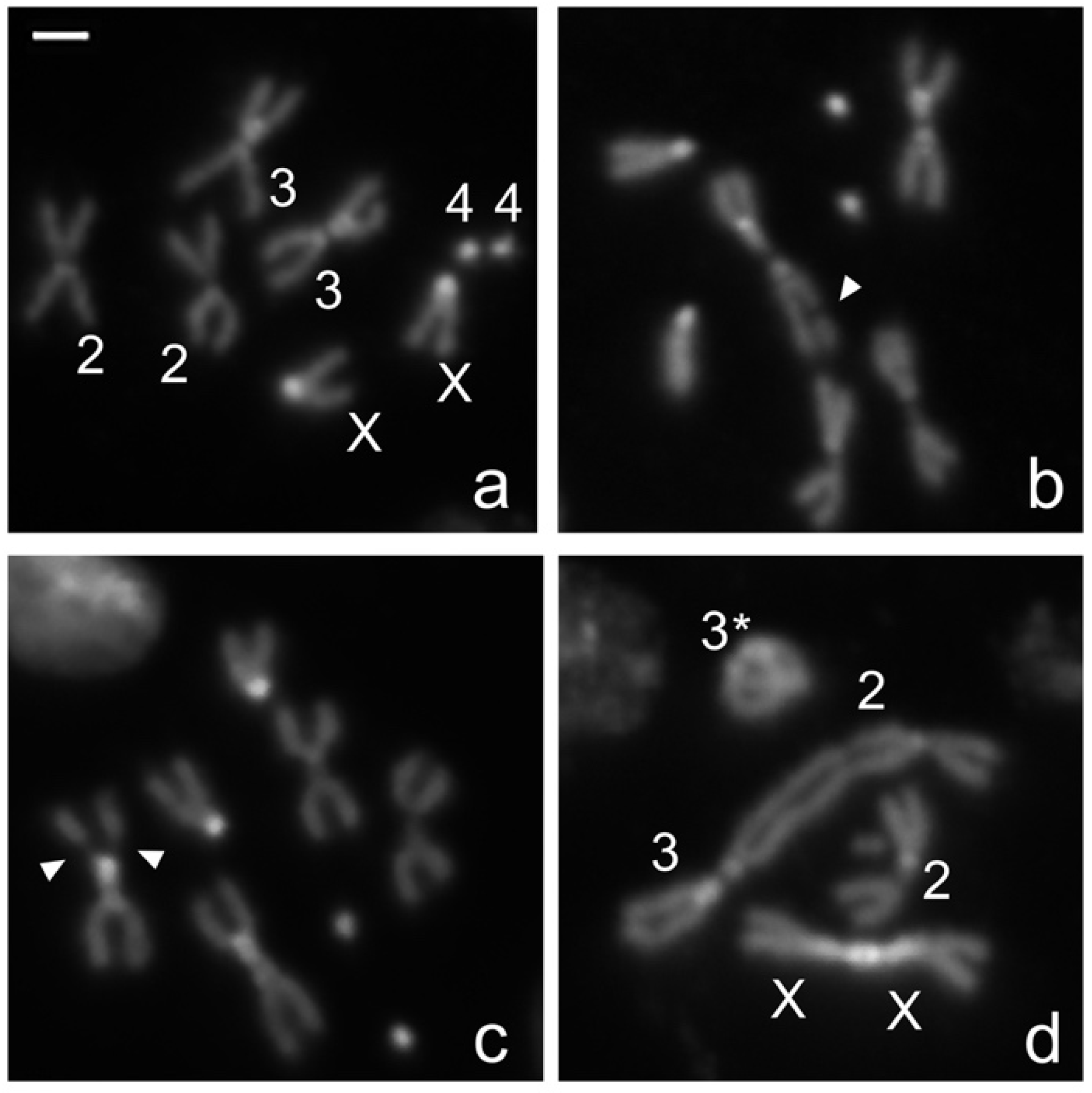{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
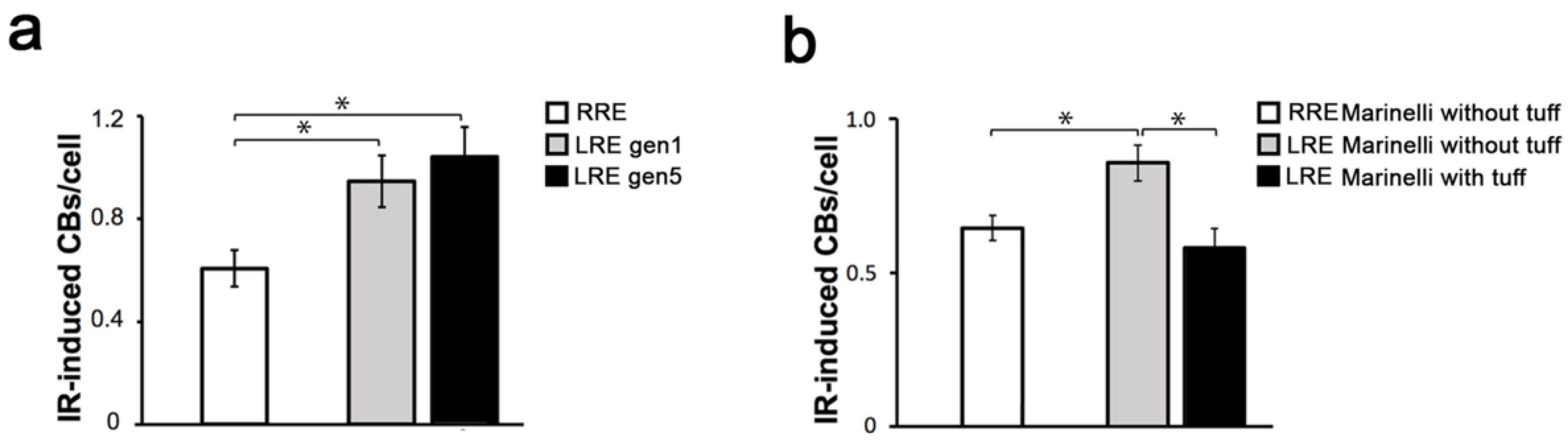
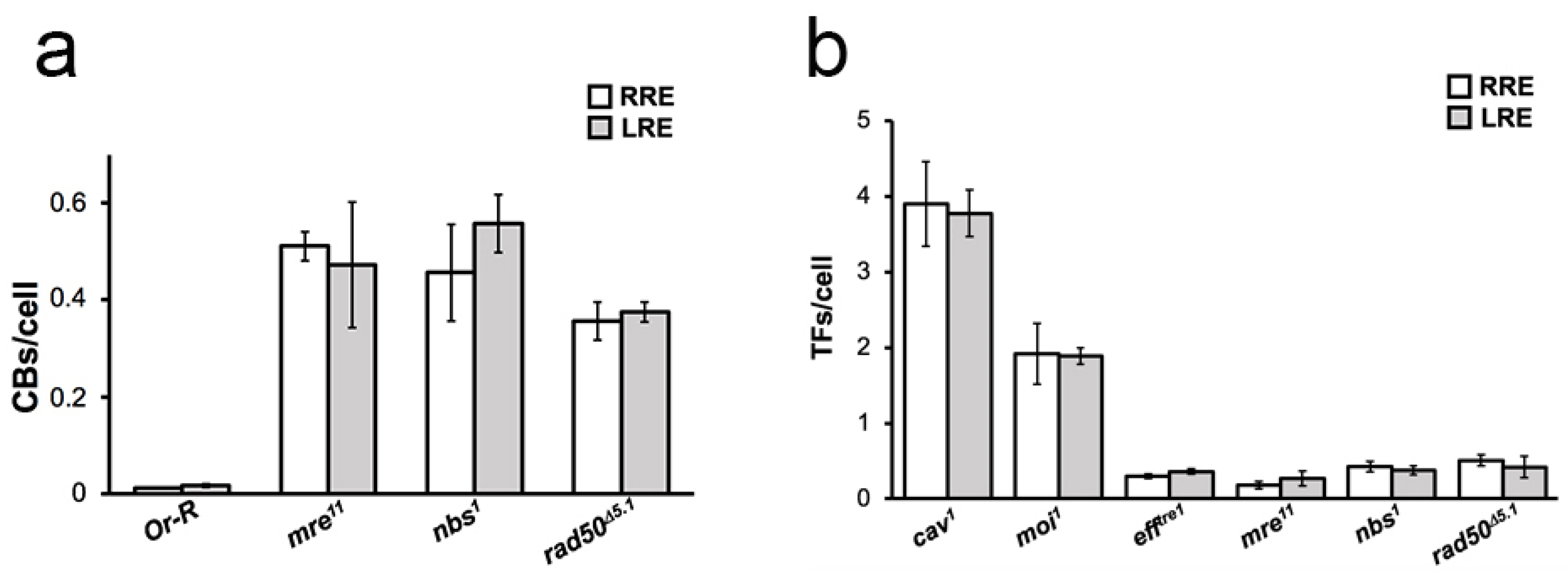
2. Results and Discussion
3. Materials and Methods
3.1. Drosophila Strains
3.2. Dosimetry
3.3. Irradiation Treatments
3.4. Chromosome Cytology and Microscopy
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belli, M.; Indovina, L. The Response of Living Organisms to Low Radiation Environment and Its Implications in Radiation Protection. Front Public Health 2020, 8, 601711. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, T.; Liu, Y.; Zou, J.; Gao, M.; Zhang, R.; Wu, J.; Liu, S.; Xie, H. History, advancements, and perspective of biological research in deep-underground laboratories: A brief review. Environ. Int. 2018, 120, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Morciano, P.; Cipressa, F.; Porrazzo, A.; Esposito, G.; Tabocchini, M.A.; Cenci, G. Fruit Flies Provide New Insights in Low-Radiation Background Biology at the INFN Underground Gran Sasso National Laboratory (LNGS). Radiat. Res. 2018, 190, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Castillo, H.; Li, X.; Smith, G.B. Deinococcus radiodurans UWO298 Dependence on Background Radiation for Optimal Growth. Front. Genet. 2021, 12, 644292. [Google Scholar] [CrossRef]
- Castillo, H.; Winder, J.; Smith, G. Chinese hamster V79 cells’ dependence on background ionizing radiation for optimal growth. Radiat. Environ. Biophys. 2021, 61, 49–57. [Google Scholar] [CrossRef]
- Van Voorhies, W.A.; Castillo, H.A.; Thawng, C.N.; Smith, G.B. The Phenotypic and Transcriptomic Response of the Caenorhabditis elegans Nematode to Background and Below-Background Radiation Levels. Front. Public Health 2020, 8, 581796. [Google Scholar] [CrossRef]
- Zarubin, M.; Gangapshev, A.; Gavriljuk, Y.; Kazalov, V.; Kravchenko, E. First transcriptome profiling of D. melanogaster after development in a deep underground low radiation background laboratory. PLoS ONE 2021, 16, e0255066. [Google Scholar] [CrossRef]
- Carbone, M.C.; Pinto, M.; Antonelli, F.; Amicarelli, F.; Balata, M.; Belli, M.; Conti Devirgiliis, L.; Ioannucci, L.; Nisi, S.; Sapora, O.; et al. The Cosmic Silence experiment: On the putative adaptive role of environmental ionizing radiation. Radiat. Environ. Biophys. 2009, 48, 189–196. [Google Scholar] [CrossRef]
- Fratini, E.; Carbone, C.; Capece, D.; Esposito, G.; Simone, G.; Tabocchini, M.A.; Tomasi, M.; Belli, M.; Satta, L. Low-radiation environment affects the development of protection mechanisms in V79 cells. Radiat. Environ. Biophys. 2015, 54, 183–194. [Google Scholar] [CrossRef]
- Satta, L.; Antonelli, F.; Belli, M.; Sapora, O.; Simone, G.; Sorrentino, E.; Tabocchini, M.A.; Amicarelli, F.; Ara, C.; Ceru, M.P.; et al. Influence of a low background radiation environment on biochemical and biological responses in V79 cells. Radiat. Environ. Biophys. 2002, 41, 217–224. [Google Scholar] [CrossRef]
- Calabrese, E.J. The linear No-Threshold (LNT) dose response model: A comprehensive assessment of its historical and scientific foundations. Chem. Biol. Interact. 2019, 301, 6–25. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Anello, P.; Ampollini, M.; Bortolin, E.; De Angelis, C.; D’Imperio, G.; Dini, V.; Nuccetelli, C.; Quattrini, M.C.; Tomei, C.; et al. Underground Radiobiology: A Perspective at Gran Sasso National Laboratory. Front. Public Health 2020, 8, 611146. [Google Scholar] [CrossRef] [PubMed]
- Morciano, P.; Iorio, R.; Iovino, D.; Cipressa, F.; Esposito, G.; Porrazzo, A.; Satta, L.; Alesse, E.; Tabocchini, M.A.; Cenci, G. Effects of reduced natural background radiation on Drosophila melanogaster growth and development as revealed by the FLYINGLOW program. J. Cell Physiol. 2018, 233, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischietti, M.; Fratini, E.; Verzella, D.; Vecchiotti, D.; Capece, D.; Di Francesco, B.; Esposito, G.; Balata, M.; Ioannuci, L.; Sykes, P.; et al. Low Radiation Environment Switches the Overgrowth-Induced Cell Apoptosis Toward Autophagy. Front. Public Health 2020, 8, 594789. [Google Scholar] [CrossRef]
- Pirkkanen, J.; Laframboise, T.; Liimatainen, P.; Sonley, T.; Stankiewicz, S.; Hood, M.; Obaid, M.; Zarnke, A.; Tai, T.C.; Lees, S.J.; et al. A novel specialized tissue culture incubator designed and engineered for radiobiology experiments in a sub-natural background radiation research environment. J. Environ. Radioact. 2021, 228, 106512. [Google Scholar] [CrossRef]
- Goodhead, D.T. Spatial and temporal distribution of energy. Health Phys. 1988, 231–240. [Google Scholar] [CrossRef]
- Campa, A.; Balduzzi, M.; Dini, V.; Esposito, G.; Tabocchini, M.A. The complex interactions between radiation induced non-targeted effects and cancer. Cancer Lett. 2015, 356, 126–136. [Google Scholar] [CrossRef]
- Castillo, H.; Schoderbek, D.; Dulal, S.; Escobar, G.; Wood, J.; Nelson, R.; Smith, G. Stress induction in the bacteria Shewanella oneidensis and Deinococcus radiodurans in response to below-background ionizing radiation. Int. J. Radiat. Biol. 2015, 91, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Sekelsky, J. DNA Repair in Drosophila: Mutagens, Models, and Missing Genes. Genetics 2017, 205, 471–490. [Google Scholar] [CrossRef] [Green Version]
- Cacchione, S.; Cenci, G.; Raffa, G.D. Silence at the End: How Drosophila Regulates Expression and Transposition of Telomeric Retroelements. J. Mol. Biol. 2020, 432, 4305–4321. [Google Scholar] [CrossRef]
- Raffa, G.D.; Ciapponi, L.; Cenci, G.; Gatti, M. Terminin: A protein complex that mediates epigenetic maintenance of Drosophila telomeres. Nucleus 2011, 2, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipressa, F.; Romano, S.; Centonze, S.; zur Lage, P.I.; Verni, F.; Dimitri, P.; Gatti, M.; Cenci, G. Effete, a Drosophila chromatin-associated ubiquitin-conjugating enzyme that affects telomeric and heterochromatic position effect variegation. Genetics 2013, 195, 147–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitri, P.; Corradini, N.; Rossi, F.; Verni, F.; Cenci, G.; Belloni, G.; Zhimulev, I.F.; Koryakov, D.E. Vital genes in the heterochromatin of chromosomes 2 and 3 of Drosophila melanogaster. Genetica 2003, 117, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Satta, L.; Augusti-Tocco, G.; Ceccarelli, R.; Esposito, A.; Fiore, M.; Paggi, P.; Poggesi, I.; Ricordy, R.; Scarsella, G.; Cundari, E. Low environmental radiation background impairs biological defence of the yeast Saccharomyces cerevisiae to chemical radiomimetic agents. Mutat. Res. Lett. 1995, 347, 3–4. [Google Scholar] [CrossRef]
- Shuryak, I.; Brenner, D.J. Review of Quantitative Mechanistic Models of Radiation-Induced Non-Targeted Effects (Nte). Radiat. Prot. Dosim. 2020, 192, 236–252. [Google Scholar] [CrossRef]
- Planel, H.; Soleilhavoup, J.P.; Tixador, R.; Richoilley, G.; Conter, A.; Croute, F.; Caratero, C.; Gaubin, Y. Influence on cell proliferation of background radiation or exposure to very low, chronic gamma radiation. Health Phys 1987, 52, 571–578. [Google Scholar] [CrossRef]
- Ciapponi, L.; Cenci, G.; Ducau, J.; Flores, C.; Johnson-Schlitz, D.; Gorski, M.M.; Engels, W.R.; Gatti, M. The Drosophila Mre11/Rad50 complex is required to prevent both telomeric fusion and chromosome breakage. Curr. Biol. 2004, 14, 1360–1366. [Google Scholar] [CrossRef] [Green Version]
- Ciapponi, L.; Cenci, G.; Gatti, M. The Drosophila Nbs protein functions in multiple pathways for the maintenance of genome stability. Genetics 2006, 173, 1447–1454. [Google Scholar] [CrossRef] [Green Version]
- Cenci, G.; Rawson, R.B.; Belloni, G.; Castrillon, D.H.; Tudor, M.; Petrucci, R.; Goldberg, M.L.; Wasserman, S.A.; Gatti, M. UbcD1, a Drosophila ubiquitin-conjugating enzyme required for proper telomere behavior. Genes Dev. 1997, 11, 863–875. [Google Scholar] [CrossRef] [Green Version]
- Cenci, G.; Siriaco, G.; Raffa, G.D.; Kellum, R.; Gatti, M. The Drosophila HOAP protein is required for telomere capping. Nat. Cell Biol. 2003, 5, 82–84. [Google Scholar] [CrossRef]
- Raffa, G.D.; Siriaco, G.; Cugusi, S.; Ciapponi, L.; Cenci, G.; Wojcik, E.; Gatti, M. The Drosophila modigliani (moi) gene encodes a HOAP-interacting protein required for telomere protection. Proc. Natl. Acad. Sci. USA 2009, 106, 2271–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cipressa, F.; Morciano, P.; Bosso, G.; Mannini, L.; Galati, A.; Raffa, G.D.; Cacchione, S.; Musio, A.; Cenci, G. A role for Separase in telomere protection. Nat. Commun. 2016, 7, 10405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messina, G.; Damia, E.; Fanti, L.; Atterrato, M.T.; Celauro, E.; Mariotti, F.R.; Accardo, M.C.; Walther, M.; Verni, F.; Picchioni, D.; et al. Yeti, an essential Drosophila melanogaster gene, encodes a protein required for chromatin organization. J. Cell Sci. 2014, 127, 2577–2588. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porrazzo, A.; Esposito, G.; Grifoni, D.; Cenci, G.; Morciano, P.; Tabocchini, M.A. Reduced Environmental Dose Rates Are Responsible for the Increased Susceptibility to Radiation-Induced DNA Damage in Larval Neuroblasts of Drosophila Grown inside the LNGS Underground Laboratory. Int. J. Mol. Sci. 2022, 23, 5472. https://doi.org/10.3390/ijms23105472
Porrazzo A, Esposito G, Grifoni D, Cenci G, Morciano P, Tabocchini MA. Reduced Environmental Dose Rates Are Responsible for the Increased Susceptibility to Radiation-Induced DNA Damage in Larval Neuroblasts of Drosophila Grown inside the LNGS Underground Laboratory. International Journal of Molecular Sciences. 2022; 23(10):5472. https://doi.org/10.3390/ijms23105472
Chicago/Turabian StylePorrazzo, Antonella, Giuseppe Esposito, Daniela Grifoni, Giovanni Cenci, Patrizia Morciano, and Maria Antonella Tabocchini. 2022. "Reduced Environmental Dose Rates Are Responsible for the Increased Susceptibility to Radiation-Induced DNA Damage in Larval Neuroblasts of Drosophila Grown inside the LNGS Underground Laboratory" International Journal of Molecular Sciences 23, no. 10: 5472. https://doi.org/10.3390/ijms23105472
APA StylePorrazzo, A., Esposito, G., Grifoni, D., Cenci, G., Morciano, P., & Tabocchini, M. A. (2022). Reduced Environmental Dose Rates Are Responsible for the Increased Susceptibility to Radiation-Induced DNA Damage in Larval Neuroblasts of Drosophila Grown inside the LNGS Underground Laboratory. International Journal of Molecular Sciences, 23(10), 5472. https://doi.org/10.3390/ijms23105472