
Potential Chemotherapeutic Effect of Selenium for Improved Canceration of Esophageal Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
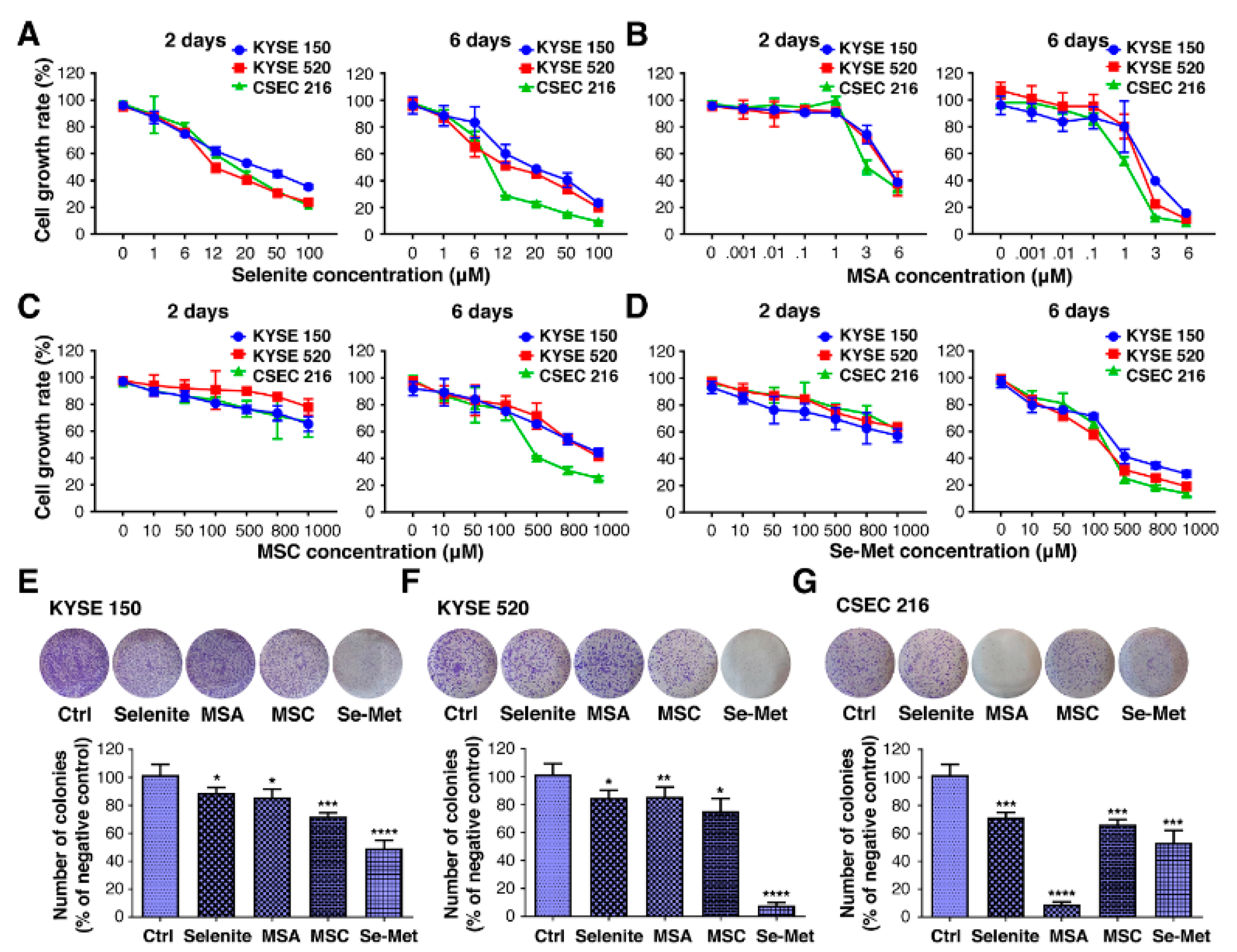
2.1. Selenium Inhibits the Proliferation and Colony Formation of ESCC Cell Lines
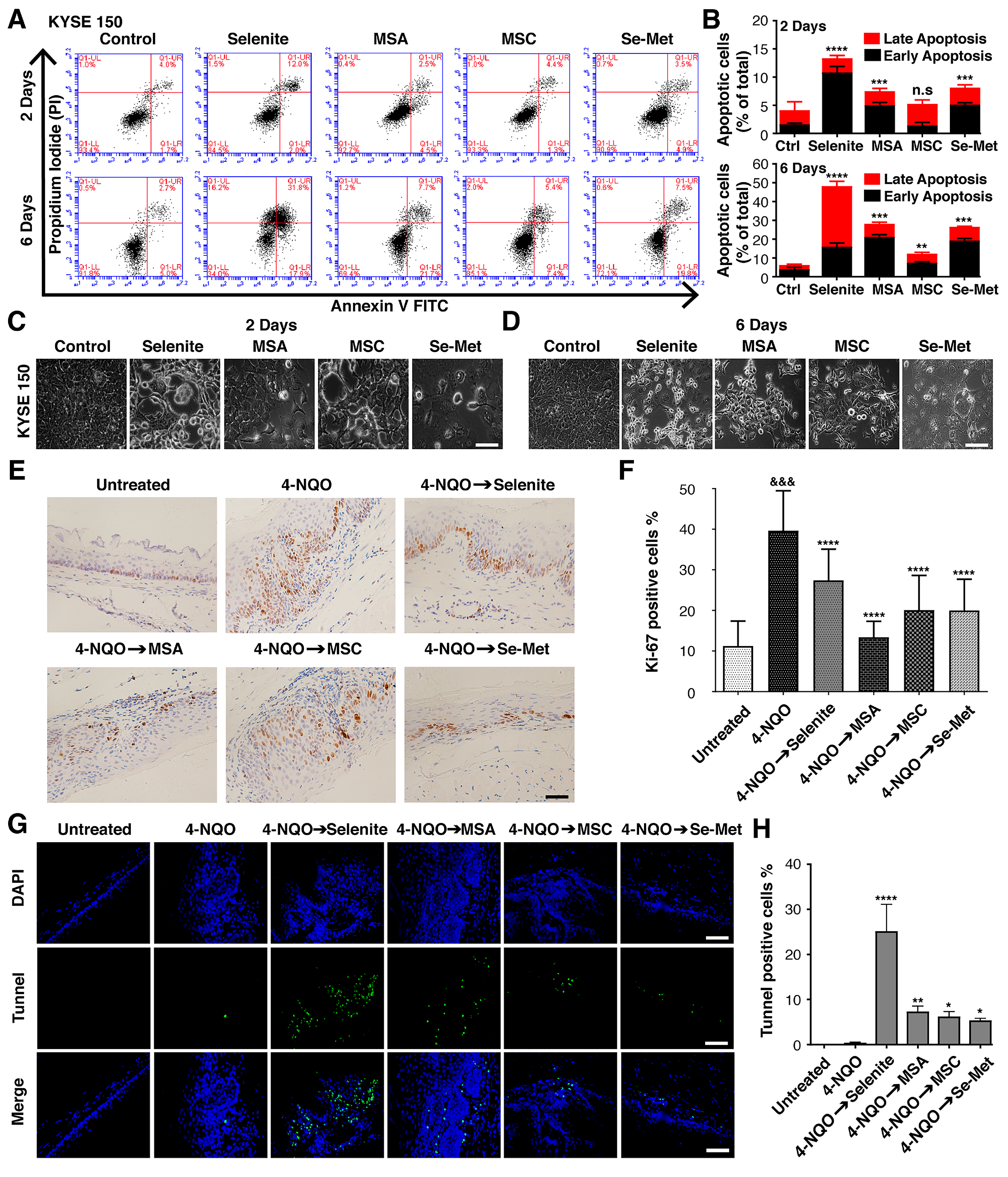
2.2. Selenium Controls the High-Grade Intraepithelial Dysplasia of Mice Induced by 4-NQO
2.3. Effectiveness of Selenium for Preventing the Development of ESCC Lesions
2.4. Increased Expression of Inflammatory Markers in ESCC That Are Reversible in Treatment with Selenium
2.5. Selenium Treatment Results in Lower ROS Stress and Oxidative DNA Damage in ESCC
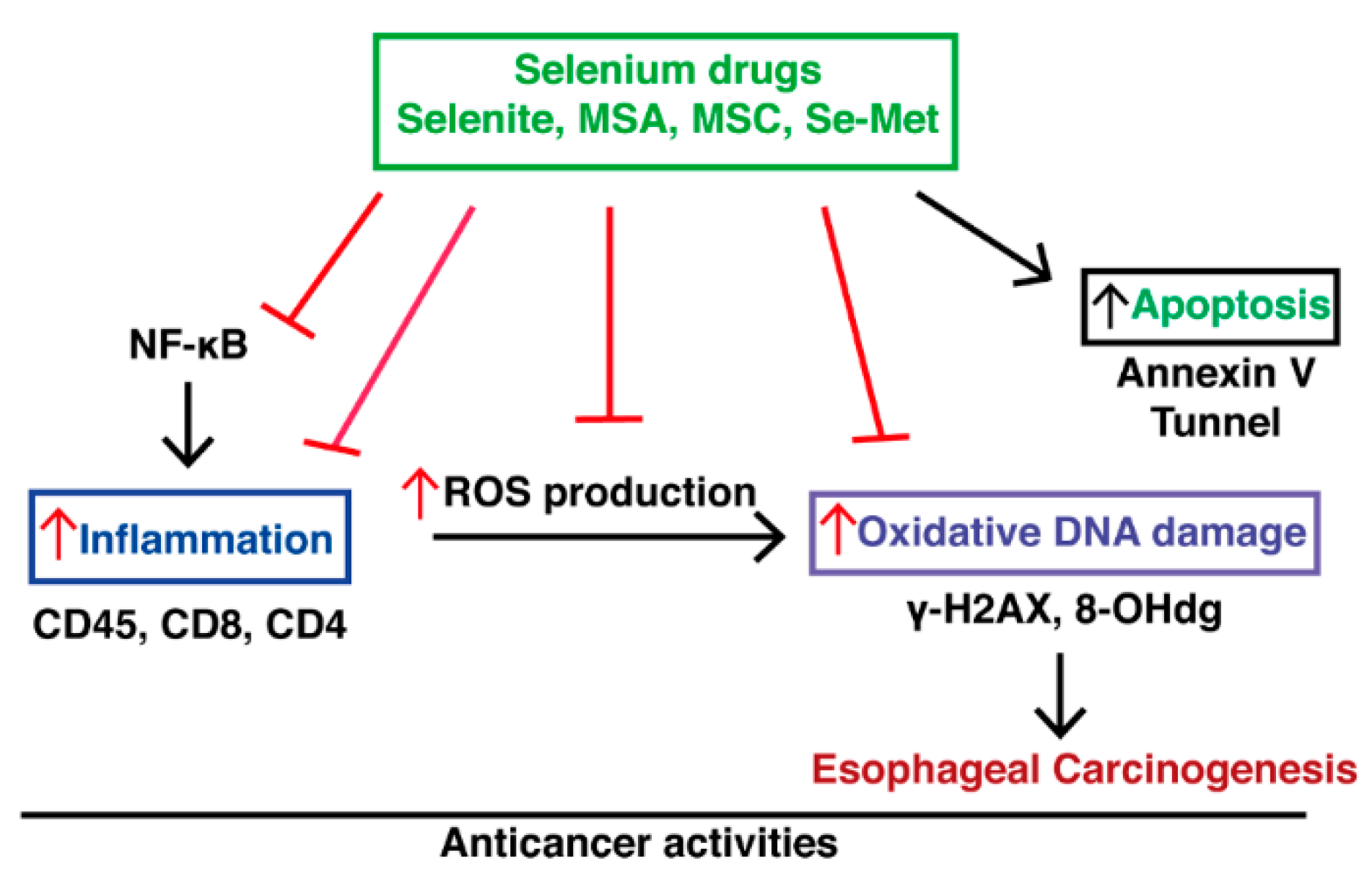
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. ESCC Cell Lines and Culture Conditions
4.3. Cell Counting Kit-8 (CCK8) Assay and Morphological Examination
4.4. Clonogenic Assay
4.5. Measurement of Apoptosis
4.6. Quantification of Intracellular ROS
4.7. Immunofluorescence Staining
4.8. Animals, Study Design, and Endpoints
4.9. Histopathological Analysis
4.10. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End Labeling (TUNEL) Assay
4.11. Immunohistochemical Staining (IHC)
4.12. Data and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y. Epidemiology of esophageal cancer. World J. Gastroenterol. 2013, 19, 5598–5606. [Google Scholar] [CrossRef]
- Su, M.; Liu, M.; Tian, D.P.; Li, X.Y.; Zhang, G.H.; Yang, H.L.; Fan, X.; Huang, H.H.; Gao, Y.X. Temporal trends of esophageal cancer during 1995–2004 in Nanao Island, an extremely high-risk area in China. Eur. J. Epidemiol. 2007, 22, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Fan, J.H.; Qiao, Y.L. Epidemiology, etiology, and prevention of esophageal squamous cell carcinoma in China. Cancer Biol. Med. 2017, 14, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Kundu, J.K.; Surh, Y.J. Inflammation: Gearing the journey to cancer. Mutat. Res. 2008, 659, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Ying, S.; Zhang, C.; Lin, R.; Zheng, J.; Zhang, G.; Tian, D.; Guo, Y.; Du, C.; et al. Genetic Alterations in Esophageal Tissues From Squamous Dysplasia to Carcinoma. Gastroenterology 2017, 153, 166–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Xiao, D.; Guo, Y.; Tian, D.; Yun, H.; Chen, D.; Su, M. Chronic inflammation-related DNA damage response: A driving force of gastric cardia carcinogenesis. Oncotarget 2015, 6, 2856–2864. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.; Zhang, C.; Zheng, J.; Tian, D.; Lei, Z.; Chen, D.; Xu, Z.; Su, M. Chronic inflammation-associated genomic instability paves the way for human esophageal carcinogenesis. Oncotarget 2016, 7, 24564–24571. [Google Scholar] [CrossRef]
- Yan, R.; Guo, Y.; Gong, Q.; Chen, M.; Guo, Y.; Yang, P.; Huang, H.; Huang, H.; Huang, W.; Ma, Z.; et al. Microbiological evidences for gastric cardiac microflora dysbiosis inducing the progression of inflammation. J. Gastroenterol. Hepatol. 2020, 35, 1032–1041. [Google Scholar] [CrossRef]
- Evans, S.O.; Khairuddin, P.F.; Jameson, M.B. Optimising Selenium for Modulation of Cancer Treatments. Anticancer Res. 2017, 37, 6497–6509. [Google Scholar] [CrossRef] [Green Version]
- Younesian, O.; Younesian, S.; Hosseinzadeh, S.; Joshaghani, H.R. Association of Selenium and Risk of Esophageal Cancer: A Review. Med. Lab. J. 2020, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mark, S.D.; Qiao, Y.L.; Dawsey, S.M.; Wu, Y.P.; Katki, H.; Gunter, E.W.; Fraumeni, J.F., Jr.; Blot, W.J.; Dong, Z.W.; Taylor, P.R. Prospective study of serum selenium levels and incident esophageal and gastric cancers. J. Natl. Cancer Inst. 2000, 92, 1753–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.Q.; Abnet, C.C.; Qiao, Y.L.; Dawsey, S.M.; Dong, Z.W.; Sun, X.D.; Fan, J.H.; Gunter, E.W.; Taylor, P.R.; Mark, S.D. Prospective study of serum selenium concentrations and esophageal and gastric cardia cancer, heart disease, stroke, and total death. Am. J. Clin. Nutr. 2004, 79, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.; Mo, H.Y.; Lau, A.T.Y.; Xu, Y.M. Selenium Species: Current Status and Potentials in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2018, 20, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandin, V.; Khalkar, P.; Braude, J.; Fernandes, A.P. Organic selenium compounds as potential chemotherapeutic agents for improved cancer treatment. Free Radic. Biol. Med. 2018, 127, 80–97. [Google Scholar] [CrossRef]
- Barrett, C.W.; Short, S.P.; Williams, C.S. Selenoproteins and oxidative stress-induced inflammatory tumorigenesis in the gut. Cell. Mol. Life Sci. 2017, 74, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Liu, T.; Sun, Y.; Yang, S.; Liang, X. Inhibitory Effect of Selenium on Esophagus Cancer Cells and the Related Mechanism. J. Nutr. Sci. Vitaminol. 2020, 66, 456–461. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Shishodia, S.; Sandur, S.K.; Pandey, M.K.; Sethi, G. Inflammation and cancer: How hot is the link? Biochem. Pharmacol. 2006, 72, 1605–1621. [Google Scholar] [CrossRef]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [Green Version]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. GammaH2AX and cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Kinner, A.; Wu, W.; Staudt, C.; Iliakis, G. Gamma-H2AX in recognition and signaling of DNA double-strand breaks in the context of chromatin. Nucleic Acids Res. 2008, 36, 5678–5694. [Google Scholar] [CrossRef] [PubMed]
- Castillo Ferrer, C.; Berthenet, K.; Ichim, G. Apoptosis—Fueling the oncogenic fire. FEBS J. 2021, 288, 4445–4463. [Google Scholar] [CrossRef]
- Pang, K.L.; Chin, K.Y. Emerging Anticancer Potentials of Selenium on Osteosarcoma. Int. J. Mol. Sci. 2019, 20, 5318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, B.; He, B.; Cai, J.; Lam, A.K.; He, W. Induction of oxidative stress and cell apoptosis by selenium: The cure against oral carcinoma. Oncotarget 2017, 8, 113614–113621. [Google Scholar] [CrossRef] [Green Version]
- Guttenplan, J.; Chen, K.M.; Khmelnitsky, M.; Kosinska, W.; Hennessy, J.; Bruggeman, R.; Desai, D.; Amin, S.; Sun, Y.W.; Spratt, T.E.; et al. Effects of 1,4-phenylenebis(methylene)selenocyanate on mutagenesis and p53 protein expression in the tongue of lacI rats treated with 4-nitroquinoline-N-oxide. Mutat. Res. 2007, 634, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Makita, H.; Kawabata, K.; Mori, H.; El-Bayoumy, K. 1,4-phenylenebis(methylene)selenocyanate exerts exceptional chemopreventive activity in rat tongue carcinogenesis. Cancer Res. 1997, 57, 3644–3648. [Google Scholar]
- Hu, C.; Liu, M.; Zhang, W.; Xu, Q.; Ma, K.; Chen, L.; Wang, Z.; He, S.; Zhu, H.; Xu, N. Upregulation of KLF4 by methylseleninic acid in human esophageal squamous cell carcinoma cells: Modification of histone H3 acetylation through HAT/HDAC interplay. Mol. Carcinog. 2015, 54, 1051–1059. [Google Scholar] [CrossRef]
- Zhang, W.; Yan, S.; Liu, M.; Zhang, G.; Yang, S.; He, S.; Bai, J.; Quan, L.; Zhu, H.; Dong, Y.; et al. beta-Catenin/TCF pathway plays a vital role in selenium induced-growth inhibition and apoptosis in esophageal squamous cell carcinoma (ESCC) cells. Cancer Lett. 2010, 296, 113–122. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.J. Breaking the relay in deregulated cellular signal transduction as a rationale for chemoprevention with anti-inflammatory phytochemicals. Mutat. Res. 2005, 591, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Jia, X.; Chen, X.; Yang, C.S.; Li, N. Time-selective chemoprevention of vitamin E and selenium on esophageal carcinogenesis in rats: The possible role of nuclear factor kappaB signaling pathway. Int. J. Cancer 2012, 131, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, S.; Ma, N.; Thanan, R.; Pinlaor, S.; Hammam, O.; Murata, M.; Kawanishi, S. DNA damage in inflammation-related carcinogenesis and cancer stem cells. Oxid. Med. Cell. Longev. 2013, 2013, 387014. [Google Scholar] [CrossRef]
- Razaghi, A.; Poorebrahim, M.; Sarhan, D.; Björnstedt, M. Selenium stimulates the antitumour immunity: Insights to future research. Eur. J. Cancer 2021, 155, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.E.; Duffield-Lillico, A.J.; Garland, L.; Turnbull, B.W.; Clark, L.C.; Marshall, J.R. Selenium supplementation and lung cancer incidence: An update of the nutritional prevention of cancer trial. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1285–1291. [Google Scholar]
- Li, X.; Tian, D.; Guo, Y.; Qiu, S.; Xu, Z.; Deng, W.; Su, M. Genomic characterization of a newly established esophageal squamous cell carcinoma cell line from China and published esophageal squamous cell carcinoma cell lines. Cancer Cell Int. 2020, 20, 184. [Google Scholar] [CrossRef]
- Ahsan, A.; Han, G.; Pan, J.; Liu, S.; Padhiar, A.A.; Chu, P.; Sun, Z.; Zhang, Z.; Sun, B.; Wu, J.; et al. Phosphocreatine protects endothelial cells from oxidized low-density lipoprotein-induced apoptosis by modulating the PI3K/Akt/eNOS pathway. Apoptosis 2015, 20, 1563–1576. [Google Scholar] [CrossRef]
- Tang, X.H.; Knudsen, B.; Bemis, D.; Tickoo, S.; Gudas, L.J. Oral cavity and esophageal carcinogenesis modeled in carcinogen-treated mice. Clin. Cancer Res. 2004, 10, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Li, Z. A sex-dimorphic mouse model of esophageal squamous cell carcinoma. Am. J. Cancer Res. 2019, 9, 429–433. [Google Scholar]
- Hawkins, B.L.; Heniford, B.W.; Ackermann, D.M.; Leonberger, M.; Martinez, S.A.; Hendler, F.J. 4NQO carcinogenesis: A mouse model of oral cavity squamous cell carcinoma. Head Neck 1994, 16, 424–432. [Google Scholar] [CrossRef]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of substances to laboratory animals: Routes of administration and factors to consider. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2011, 50, 600–613. [Google Scholar]
- Yang, H.; Jia, X. Safety evaluation of Se-methylselenocysteine as nutritional selenium supplement: Acute toxicity, genotoxicity and subchronic toxicity. Regul. Toxicol. Pharmacol. 2014, 70, 720–727. [Google Scholar] [CrossRef]
- Cao, S.; Durrani, F.A.; Rustum, Y.M. Selective modulation of the therapeutic efficacy of anticancer drugs by selenium containing compounds against human tumor xenografts. Clin. Cancer Res. 2004, 10, 2561–2569. [Google Scholar] [CrossRef] [Green Version]
- Li, G.X.; Lee, H.J.; Wang, Z.; Hu, H.; Liao, J.D.; Watts, J.C.; Combs, G.F., Jr.; Lu, J. Superior in vivo inhibitory efficacy of methylseleninic acid against human prostate cancer over selenomethionine or selenite. Carcinogenesis 2008, 29, 1005–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagtegaal, I.D.; Odze, R.D.; Klimstra, D.; Paradis, V.; Rugge, M.; Schirmacher, P.; Washington, K.M.; Carneiro, F.; Cree, I.A. The 2019 WHO classification of tumours of the digestive system. Histopathology 2020, 76, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahsan, A.; Liu, Z.; Su, R.; Liu, C.; Liao, X.; Su, M. Potential Chemotherapeutic Effect of Selenium for Improved Canceration of Esophageal Cancer. Int. J. Mol. Sci. 2022, 23, 5509. https://doi.org/10.3390/ijms23105509
Ahsan A, Liu Z, Su R, Liu C, Liao X, Su M. Potential Chemotherapeutic Effect of Selenium for Improved Canceration of Esophageal Cancer. International Journal of Molecular Sciences. 2022; 23(10):5509. https://doi.org/10.3390/ijms23105509
Chicago/Turabian StyleAhsan, Anil, Zhiwei Liu, Ruibing Su, Chencai Liu, Xiaoqi Liao, and Min Su. 2022. "Potential Chemotherapeutic Effect of Selenium for Improved Canceration of Esophageal Cancer" International Journal of Molecular Sciences 23, no. 10: 5509. https://doi.org/10.3390/ijms23105509
APA StyleAhsan, A., Liu, Z., Su, R., Liu, C., Liao, X., & Su, M. (2022). Potential Chemotherapeutic Effect of Selenium for Improved Canceration of Esophageal Cancer. International Journal of Molecular Sciences, 23(10), 5509. https://doi.org/10.3390/ijms23105509