
Fascial Innervation: A Systematic Review of the Literature


 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
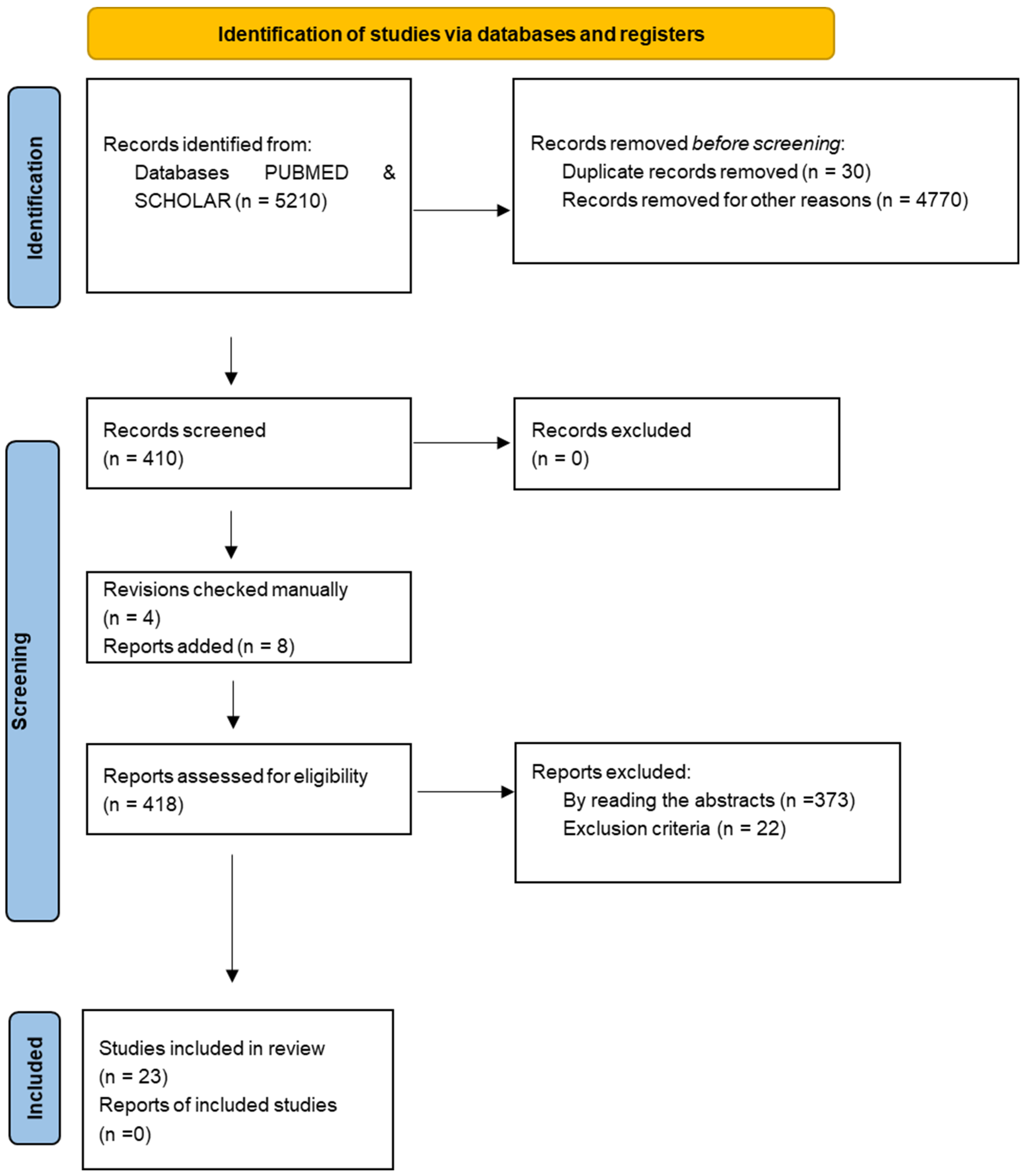
2. Materials and Methods
3. Results
3.1. Sample
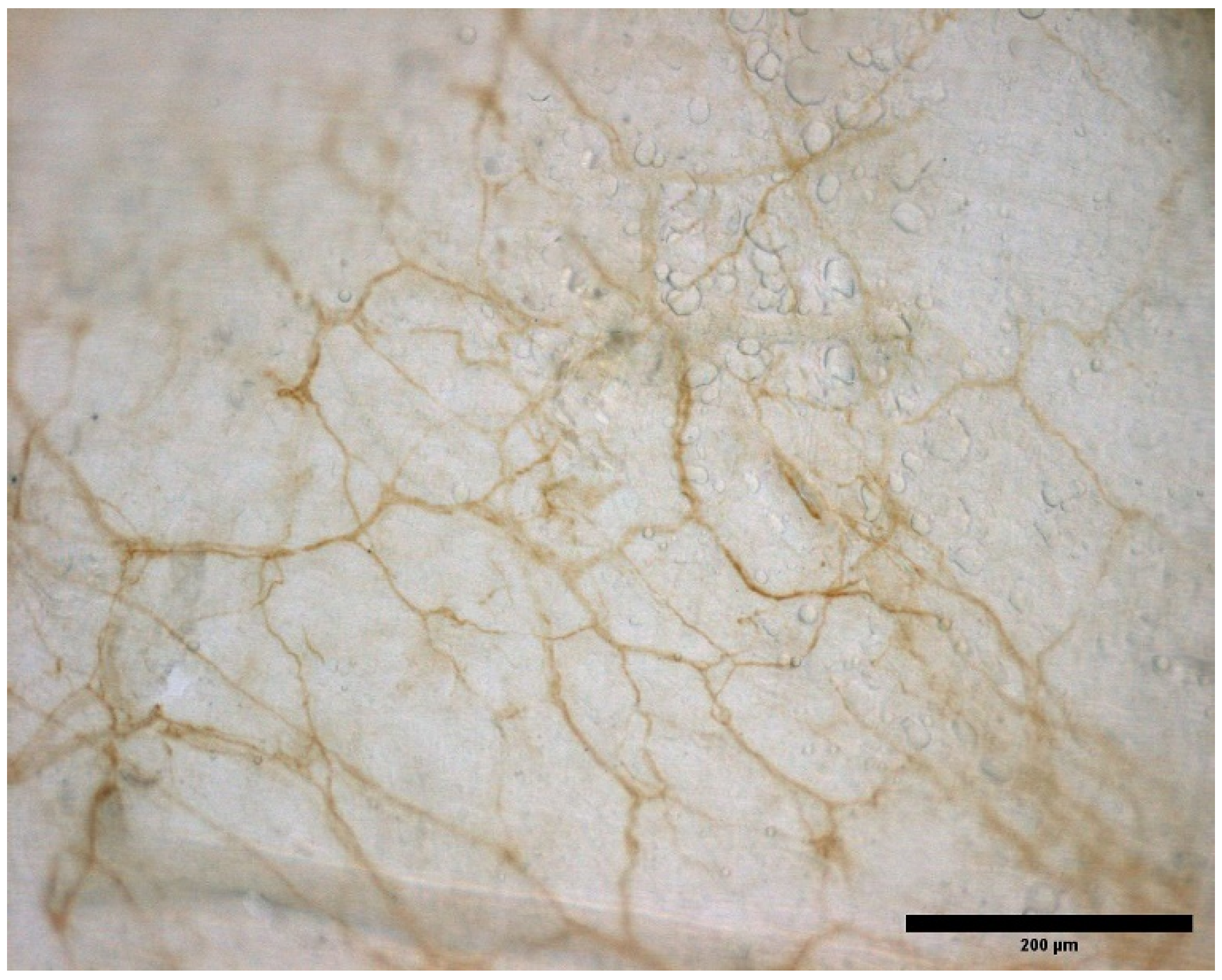
3.2. Type of Immunohistochemical Markers
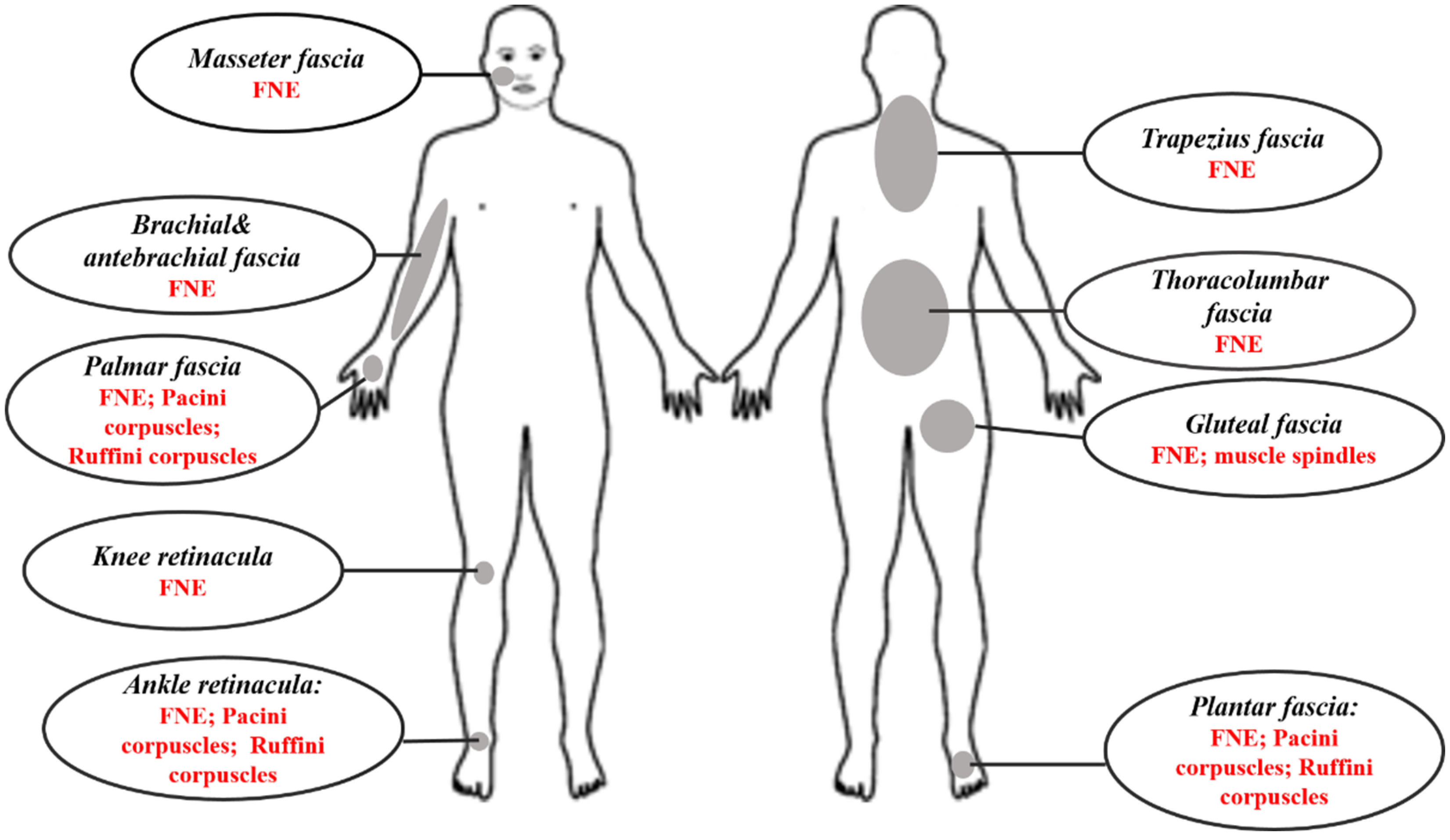
3.3. Type and Localization of Nerve Receptors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sakada, S. Mechanoreceptors in Fascia, Periosteum and Periodontal Ligament. Bull. Tokyo Med. Dent. Univ. 1974, 21, 11–13. [Google Scholar] [PubMed]
- Stilwell, D.L. Regional Variations in the Innervation of Deep Fasciae and Aponeuroses. Anat. Rec. 1957, 127, 635–653. [Google Scholar] [CrossRef] [PubMed]
- Still, A.T. The Philosophy and Mechanical Principles of Osteopathy; Hudson-Kimberly Pub. Co.: Kansas City, MO, USA, 1902. [Google Scholar]
- Findley, T.W.; Shalwala, M. Fascia Research Congress Evidence from the 100 Year Perspective of Andrew Taylor Still. J. Bodyw. Mov. Ther. 2013, 17, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Stecco, C. Functional Atlas of the Human Fascial System; Churchill Livingstone Elsevier: London, UK, 2015. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, n71. [Google Scholar] [CrossRef] [PubMed]
- Gajda, M.; Litwin, J.A.; Adriaensen, D.; Timmermans, J.-P.; Cichocki, T. Segmental Distribution and Morphometric Features of Primary Sensory Neurons Projecting to the Tibial Periosteum in the Rat. Folia Histochem. Cytobiol. 2004, 42, 95–99. [Google Scholar] [PubMed]
- Corey, S.M.; Vizzard, M.A.; Badger, G.J.; Langevin, H.M. Sensory Innervation of the Nonspecialized Connective Tissues in the Low Back of the Rat. Cells Tissues Organs 2011, 194, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, T.; Yasui, M.; Kubo, A.; Abe, M.; Kiyama, H.; Yamanaka, A.; Mizumura, K. Nociception Originating from the Crural Fascia in Rats. Pain 2013, 154, 1103–1114. [Google Scholar] [CrossRef]
- Hoheisel, U.; Rosner, J.; Mense, S. Innervation Changes Induced by Inflammation of the Rat Thoracolumbar Fascia. Neuroscience 2015, 300, 351–359. [Google Scholar] [CrossRef]
- Mense, S.; Hoheisel, U. Evidence for the Existence of Nociceptors in Rat Thoracolumbar Fascia. J. Bodyw. Mov. Ther. 2016, 20, 623–628. [Google Scholar] [CrossRef]
- Martin, C.D.; Jimenez-Andrade, J.M.; Ghilardi, J.R.; Mantyh, P.W. Organization of a Unique Net-like Meshwork of CGRP+ Sensory Fibers in the Mouse Periosteum: Implications for the Generation and Maintenance of Bone Fracture Pain. Neurosci. Lett. 2007, 427, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.M.; Kestell, G.; Gillan, M.; Haberberger, R.V.; Gibbins, I.L. Sensory Nerve Fibers Containing Calcitonin Gene-Related Peptide in Gastrocnemius, Latissimus Dorsi and Erector Spinae Muscles and Thoracolumbar Fascia in Mice. Neuroscience 2015, 291, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Thai, J.; Kyloh, M.; Travis, L.; Spencer, N.J.; Ivanusic, J.J. Identifying Spinal Afferent (Sensory) Nerve Endings That Innervate the Marrow Cavity and Periosteum Using Anterograde Tracing. J. Comp. Neurol. 2020, 528, 1903–1916. [Google Scholar] [CrossRef] [PubMed]
- Fede, C.; Petrelli, L.; Guidolin, D.; Porzionato, A.; Pirri, C.; Fan, C.; de Caro, R.; Stecco, C. Evidence of a New Hidden Neural Network into Deep Fasciae. Sci. Rep. 2021, 11, 12623. [Google Scholar] [CrossRef] [PubMed]
- Lusi, C.M.; Davies, H.M.S. The Connectivity and Histological Structure of the Equine Ergot—A Preliminary Study. J. Equine Vet. Sci. 2017, 51, 79–85. [Google Scholar] [CrossRef]
- Skalec, A.; Egerbacher, M. The Deep Fascia and Retinacula of the Equine Forelimb—Structure and Innervation. J. Anat. 2017, 231, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Stecco, C.; Gagey, O.; Belloni, A.; Pozzuoli, A.; Porzionato, A.; Macchi, V.; Aldegheri, R.; de Caro, R.; Delmas, V. Anatomy of the Deep Fascia of the Upper Limb. Second Part: Study of Innervation. Morphologie 2007, 91, 38–43. [Google Scholar] [CrossRef]
- Benetazzo, L.; Bizzego, A.; de Caro, R.; Frigo, G.; Guidolin, D.; Stecco, C. 3D Reconstruction of the Crural and Thoracolumbar Fasciae. Surg. Radiol. Anat. 2011, 33, 855–862. [Google Scholar] [CrossRef]
- Domingo, T.; Blasi, J.; Casals, M.; Mayoral, V.; Ortiz-Sagristá, J.C.; Miguel-Pérez, M. Is Interfascial Block with Ultrasound-Guided Puncture Useful in Treatment of Myofascial Pain of the Trapezius Muscle? Clin. J. Pain 2011, 27, 297–303. [Google Scholar] [CrossRef]
- Stecco, C.; Corradin, M.; Macchi, V.; Morra, A.; Porzionato, A.; Biz, C.; de Caro, R. Plantar Fascia Anatomy and Its Relationship with Achilles Tendon and Paratenon. J. Anat. 2013, 223, 665–676. [Google Scholar] [CrossRef]
- Satoh, M.; Yoshino, H.; Fujimura, A.; Hitomi, J.; Isogai, S. Three-Layered Architecture of the Popliteal Fascia That Acts as a Kinetic Retinaculum for the Hamstring Muscles. Anat. Sci. Int. 2016, 91, 341–349. [Google Scholar] [CrossRef]
- Marpalli, S.; Mohandas Rao, K.G.; Venkatesan, P.; George, B.M. The Morphological and Microscopical Characteristics of Posterior Layer of Human Thoracolumbar Fascia; A Potential Source of Low Back Pain. Morphologie 2021, 105, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Sanchis-Alfonso, V.; Roselló -Sastre, E. Immunohistochemical Analysis for Neural Markers of the Lateral Retinaculum in Patients with Isolated Symptomatic Patellofemoral Malalignment. A Neuroanatomic Basis for Anterior Knee Pain in the Active Young Patient. Am. J. Sports Med. 2000, 28, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Alhilou, A.M.; Shimada, A.; Svensson, C.I.; Ernberg, M.; Cairns, B.E.; Christidis, N. Density of Nerve Fibres and Expression of Substance P, NR2B-Receptors and Nerve Growth Factor in Healthy Human Masseter Muscle: An Immunohistochemical Study. J. Oral Rehabil. 2021, 48, 35–44. [Google Scholar] [CrossRef]
- Stecco, C.; Macchi, V.; Barbieri, A.; Tiengo, C.; Porzionato, A.; de Caro, R. Hand Fasciae Innervation: The Palmar Aponeurosis. Clin. Anat. 2018, 31, 677–683. [Google Scholar] [CrossRef]
- Fede, C.; Porzionato, A.; Petrelli, L.; Fan, C.; Pirri, C.; Biz, C.; de Caro, R.; Stecco, C. Fascia and Soft Tissues Innervation in the Human Hip and Their Possible Role in Post-Surgical Pain. J. Orthop. Res. 2020, 38, 1646–1654. [Google Scholar] [CrossRef]
- Tesarz, J.; Hoheisel, U.; Wiedenhöfer, B.; Mense, S. Sensory Innervation of the Thoracolumbar Fascia in Rats and Humans. Neuroscience 2011, 194, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Mense, S. Innervation of the Thoracolumbar Fascia. Eur. J. Transl. Myol. 2019, 29, 151–158.e8297. [Google Scholar] [CrossRef]
- Langevin, H.M.; Huijing, P.A. Communicating about Fascia: History, Pitfalls, and Recommendations. Int. J. Ther. Massage Bodyw. 2009, 2, 3. [Google Scholar]
- Wilke, J.; Schleip, R.; Klingler, W.; Stecco, C. The Lumbodorsal Fascia as a Potential Source of Low Back Pain: A Narrative Review. BioMed Res. Int. 2017, 2017, 5349620. [Google Scholar] [CrossRef] [Green Version]
- Gibson, W.; Arendt-Nielsen, L.; Mizumura, T.T.K.; Graven-Nielsen, T. Increased Pain from Muscle Fascia Following Eccentric Exercise: Animal and Human Findings. Exp. Brain Res. 2009, 194, 299–308. [Google Scholar] [CrossRef]
- Schilder, A.; Hoheisel, U.; Magerl, W.; Benrath, J.; Klein, T.; Treede, R.D. Sensory Findings after Stimulation of the Thoracolumbar Fascia with Hypertonic Saline Suggest Its Contribution to Low Back Pain. PAIN 2014, 155, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Langevin, H.M. Fascia Mobility, Proprioception, and Myofascial Pain. Life 2021, 11, 668. [Google Scholar] [CrossRef] [PubMed]
- Langevin, H.M.; Stevens-Tuttle, D.; Fox, J.R.; Badger, G.J.; Bouffard, N.A.; Krag, M.H.; Wu, J.; Henry, S.M. Ultrasound Evidence of Altered Lumbar Connective Tissue Structure in Human Subjects with Chronic Low Back Pain. BMC Musculoskelet. Disord. 2009, 10, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langevin, H.M.; Fox, J.R.; Koptiuch, C.; Badger, G.J.; Greenan- Naumann, A.C.; Bouffard, N.A.; Konofagou, E.E.; Lee, W.N.; Triano, J.J.; Henry, S.M. Reduced Thoracolumbar Fascia Shear Strain in Human Chronic Low Back Pain. BMC Musculoskelet. Disord. 2011, 12, 203. [Google Scholar] [CrossRef] [PubMed]
- Stecco, A.; Gesi, M.; Stecco, C.; Stern, R. Fascial Components of the Myofascial Pain Syndrome Topical Collection on Myofascial Pain. Curr. Pain Headache Rep. 2013, 17, 352. [Google Scholar] [CrossRef]
- Stecco, C.; Stern, R.; Porzionato, A.; MacChi, V.; Masiero, S.; Stecco, A.; de Caro, R. Hyaluronan within Fascia in the Etiology of Myofascial Pain. Surg. Radiol. Anat. 2011, 33, 891–896. [Google Scholar] [CrossRef]
- Casato, G.; Stecco, C.; Busin, R. Fasciae and Nonspecific Low Back Pain Role of Fasciae in Nonspecific Low Back Pain. Eur. J. Transl. Myol. 2019, 29, 8330. [Google Scholar] [CrossRef]
- Grace Ganjaei, K.; Ray, J.W.; Waite, B.; Burnham, K.J. The Fascial System in Musculoskeletal Function and Myofascial Pain. Sports Med. Rehabil. 2020, 8, 364–372. [Google Scholar] [CrossRef]
- Schilder, A.; Magerl, W.; Hoheisel, U.; Klein, T.; Treede, R.D. Electrical High-Frequency Stimulation of the Human Thoracolumbar Fascia Evokes Long-Term Potentiation-like Pain Amplification. Pain 2016, 157, 2309–2317. [Google Scholar] [CrossRef]
- Deising, S.; Weinkauf, B.; Blunk, J.; Obreja, O.; Schmelz, M.; Rukwied, R. NGF-Evoked Sensitization of Muscle Fascia Nociceptors in Humans. Pain 2012, 153, 1673–1679. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Marker | Detection | Study |
|---|---|---|
| SP | Nociceptors | Sanchis-Alfonso and Rosello-Sastre, 2000 [19]; Tesarz et al., 2011 [22]; Barry et al., 2015 [8]; Hoheisel, Rosner and Mense, 2015 [24]; Mense and Hoheisel 2016 [25]; Mense, 2019 [23]; Alhilou et al., 2020 [26] |
| PGP 9.5 | Neuronal | Sanchis-Alfonso and Rosello-Sastre, 2000 [19]; Corey et al., 2011 [27]; Tesarz et al., 2011 [22]; Taguchi et al., 2013 [28]; Barry et al., 2015 [8]; Hoheisel, Rosner and Mense, 2015 [24]; Mense and Hoheisel 2016 [25]; Skalec and Egerbacher, 2017 [12]; Mense, 2019 [23]; Alhilou et al., 2020 [26]; Thai et al., 2020 [9]; Fede et al., 2021 [10] |
| CGRP | Nociceptors | Sanchis-Alfonso and Rosello-Sastre, 2000 [19]; Martin et al., 2007 [7]; Corey et al., 2011 [27]; Tesarz et al., 2011 [22]; Taguchi et al., 2013 [28]; Barry et al., 2015 [8]; Hoheisel, Rosner and Mense, 2015 [24]; Mense and Hoheisel 2016 [25]; Mense, 2019 [23]; Thai et al., 2020 [9] |
| NGF | Sensory and sympathetic neurons | Sanchis-Alfonso and Rosello-Sastre, 2000 [19]; Mense, 2019 [23]; Alhilou et al., 2020 [26] |
| TH | Postganglionic sympathetic fibers- dopaminergic and noradrenergic neurons | Martin et al., 2007 [7]; Tesarz et al., 2011 [22]; Hoheisel, Rosner and Mense, 2015 [24]; Mense, 2019 [23]; Thai et al., 2020 [9]; Fede et al., 2021 [10] |
| S100 | Schwann cells | Sanchis-Alfonso and Rosello-Sastre, 2000 [19]; Stecco et al., 2007 [13]; Benetazzo et al., 2011 [14]; Domingo et al., 2011 [15]; Stecco et al., 2013 [16]; Skalec and Egerbacher, 2017 [12]; Lusi and Davies, 2017 [11]; Stecco et al., 2018 [20]; Fede et al., 2020 [21]; Fede et al., 2021 [10] |
| Neurofilament protein | Complex networks of axons | Sanchis-Alfonso and Rosello-Sastre, 2000 [19] |
| DiI | Tracer for neuronal and other cells | Gajda et al., 2004 [29] |
| NF200 | Neuronal marker | Martin et al., 2007 [7]; Thai et al., 2020 [9] |
| MBP | Myelinating glia | Domingo et al., 2011 [15] |
| Peripherin | Peripheral neurons, including enteric ganglion cells | Taguchi et al., 2013 [28] |
| NSE | Neuron specific enolase | Barry et al., 2015 [8] |
| TRPV1 | Epidermal and dermal cells, as well as free nerve fibers and Merkel cells | Mense and Hoheisel 2016 [25]; Mense, 2019 [23] |
| RT97 | Neuronal marker | Satoh et al., 2016 [17] |
| Tubulin | Neurons | Stecco et al., 2018 [20] |
| NR2B | Glutamatergic neuron | Alhilou et al., 2020 [26] |
| Type of Receptors (Mean Number/cm2) | Brachial Fascia | Lacertus Fibrosus | Antebrachial Fascia | Flexor Retinaculum |
|---|---|---|---|---|
| Free nerve endings | 48.57 | 27.36 | 44.37 | 53.55 |
| Pacini corpuscles | 0.43 | 0.26 | 0.26 | 0.66 |
| Ruffini corpuscles | 0.29 | 0.1 | 0.26 | 0.55 |
| Markers | CGRP+ | CGRP- | NF200+ | NF200- | NF200+ CGRP- | NF200+ CGRP+ | NF200- CGRP+ | NF200- CGRP- |
|---|---|---|---|---|---|---|---|---|
| N° of fibers | >20 | 5–20 | 5–20 | >20 | 5–20 | 5 | >20 | >20 |
| Descriptive | Sanchis-Alfonso and Rosello-Sastre, 2000 [19]; Gajda et al., 2004 [29]; Domingo et al., 2011 [15]; Stecco et al., 2013 [16]; Satoh et al., 2016 [17]; Skalec and Egerbacher, 2017 [12]; Lusi and Davies, 2017 [11] |
| Number | Stecco et al., 2007 [13]; Martin et al., 2007 [7]; Benetazzo et al., 2011 [14]; Taguchi et al., 2013 [28]; Thai et al., 2020 [9] |
| Thickness | Fede et al., 2021 [10] |
| Length | Taguchi et al., 2013 [28]; Hoheisel, Rosner and Mense, 2015 [24]; Mense and Hoheisel 2016 [25]; Mense, 2019 [23]; Fede et al., 2021 [10] |
| Diameter | Corey et al., 2011 [27]; Benetazzo et al., 2011 [14]; Fede et al., 2020 [21] |
| Proportion | Corey et al., 2011 [27]; Barry et al., 2015 [8] |
| Density | Corey et al., 2011 [27]; Tesarz et al., 2011 [22]; Barry et al., 2015 [8]; Stecco et al., 2018 [20]; Alhilou et al., 2020 [26]; Fede et al., 2020 [21]; Fede et al., 2021 [10] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suarez-Rodriguez, V.; Fede, C.; Pirri, C.; Petrelli, L.; Loro-Ferrer, J.F.; Rodriguez-Ruiz, D.; De Caro, R.; Stecco, C. Fascial Innervation: A Systematic Review of the Literature. Int. J. Mol. Sci. 2022, 23, 5674. https://doi.org/10.3390/ijms23105674
Suarez-Rodriguez V, Fede C, Pirri C, Petrelli L, Loro-Ferrer JF, Rodriguez-Ruiz D, De Caro R, Stecco C. Fascial Innervation: A Systematic Review of the Literature. International Journal of Molecular Sciences. 2022; 23(10):5674. https://doi.org/10.3390/ijms23105674
Chicago/Turabian StyleSuarez-Rodriguez, Vidina, Caterina Fede, Carmelo Pirri, Lucia Petrelli, Juan Francisco Loro-Ferrer, David Rodriguez-Ruiz, Raffaele De Caro, and Carla Stecco. 2022. "Fascial Innervation: A Systematic Review of the Literature" International Journal of Molecular Sciences 23, no. 10: 5674. https://doi.org/10.3390/ijms23105674
APA StyleSuarez-Rodriguez, V., Fede, C., Pirri, C., Petrelli, L., Loro-Ferrer, J. F., Rodriguez-Ruiz, D., De Caro, R., & Stecco, C. (2022). Fascial Innervation: A Systematic Review of the Literature. International Journal of Molecular Sciences, 23(10), 5674. https://doi.org/10.3390/ijms23105674