
SK119, a Novel Shikonin Derivative, Leads to Apoptosis in Melanoma Cell Lines and Exhibits Synergistic Effects with Vemurafenib and Cobimetinib

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
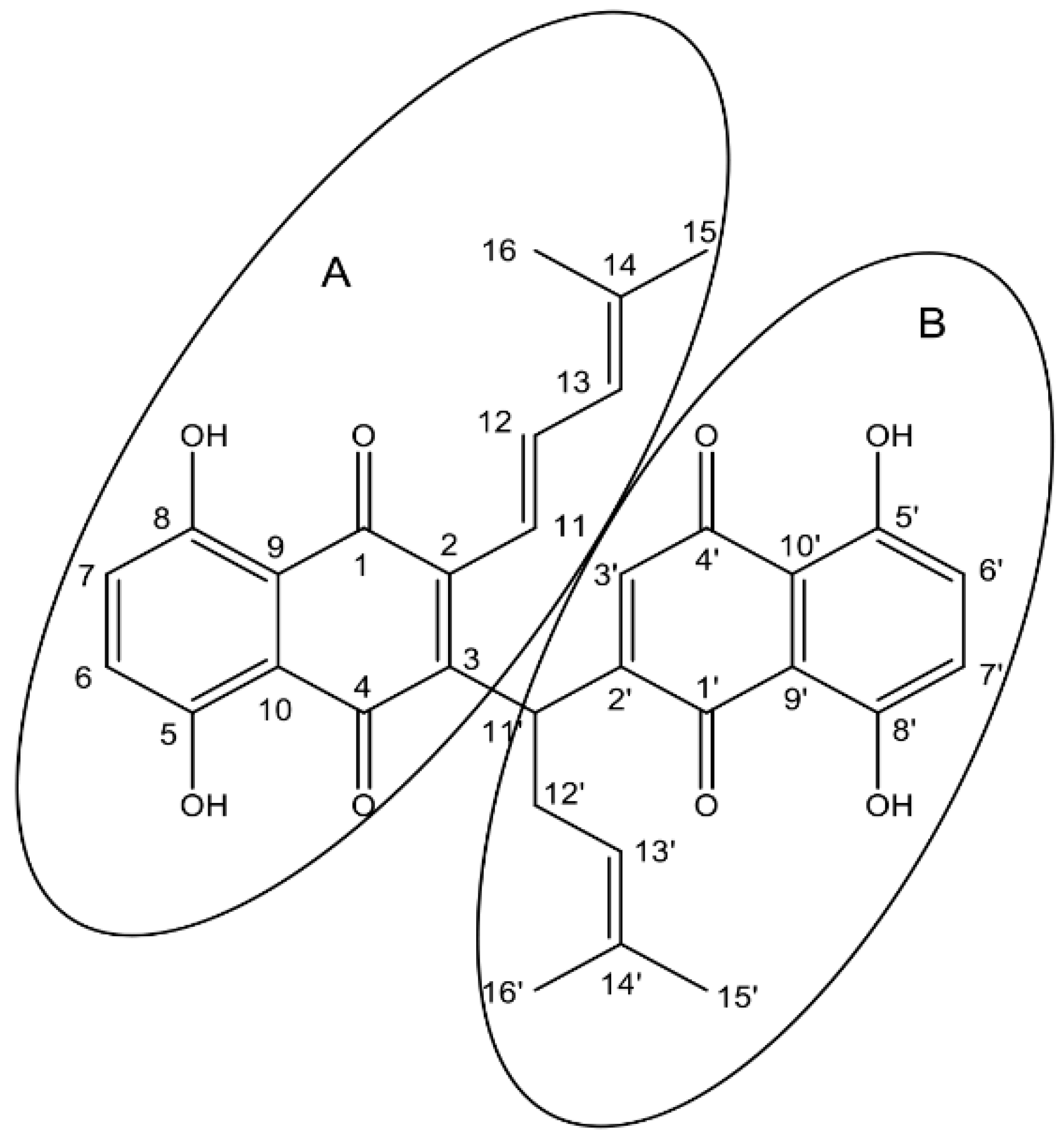
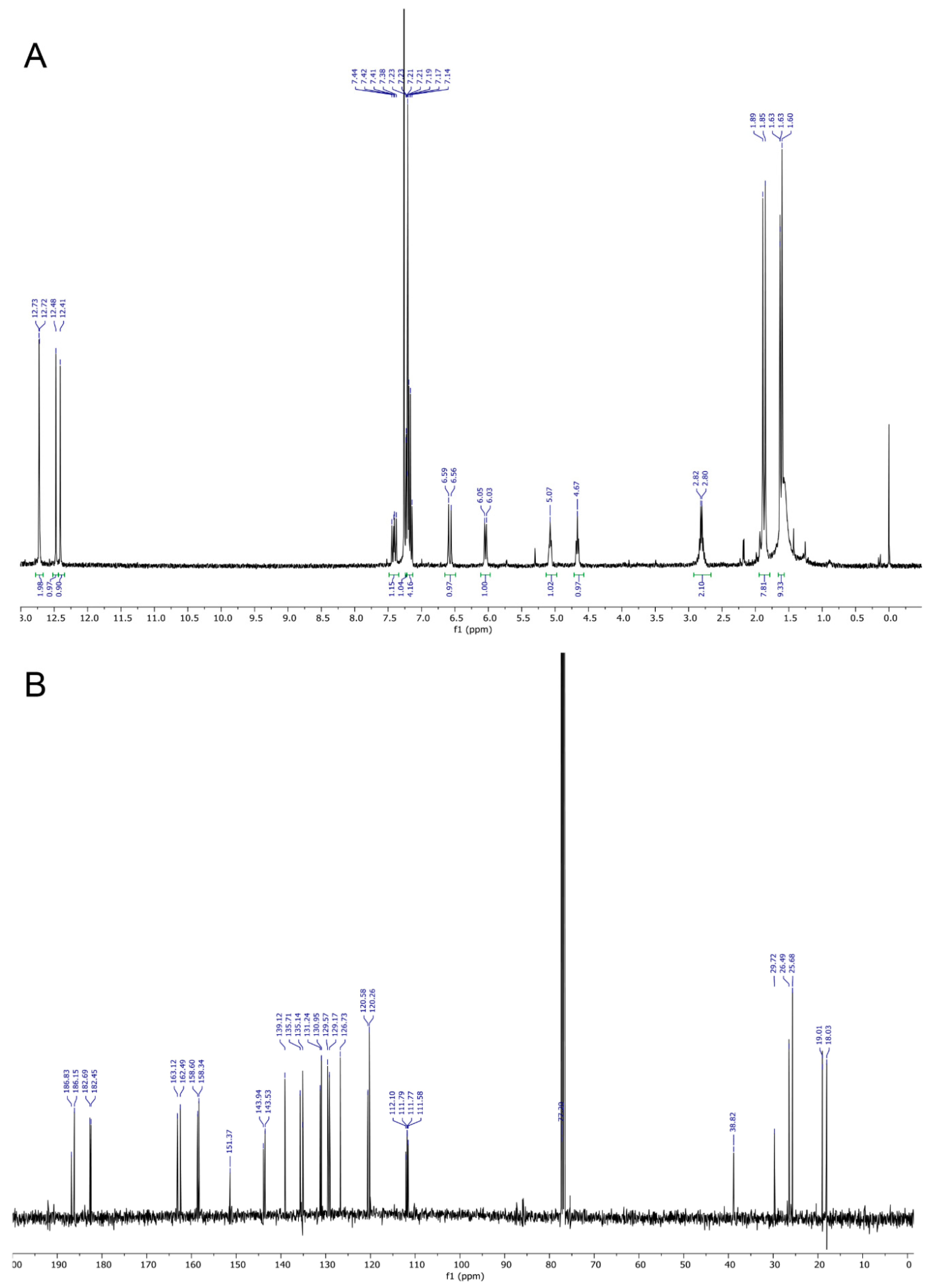
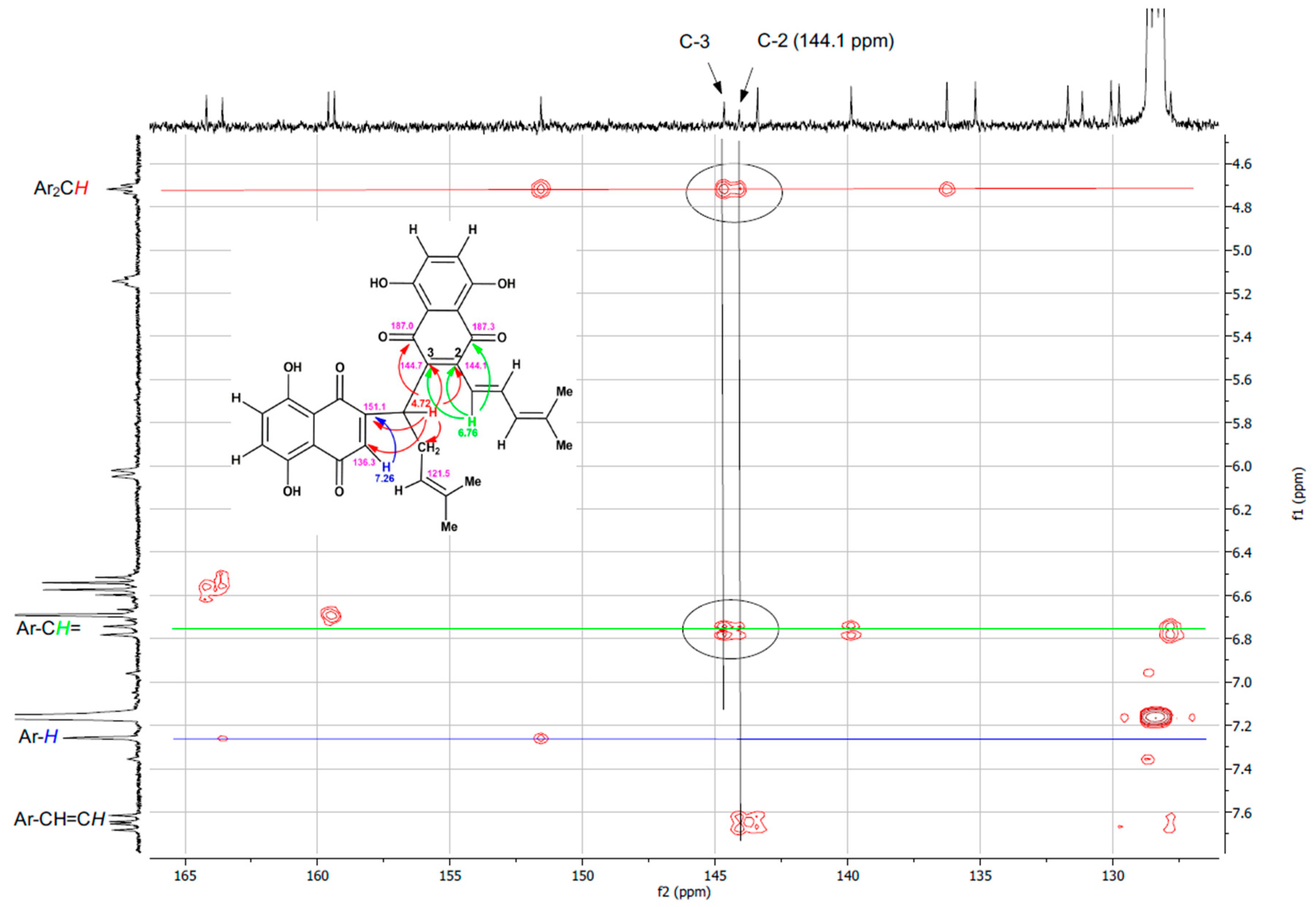
2.1. Synthesis and Structure Elucidation of SK119
2.2. Screening of SK119 against Different Melanoma Cell Lines
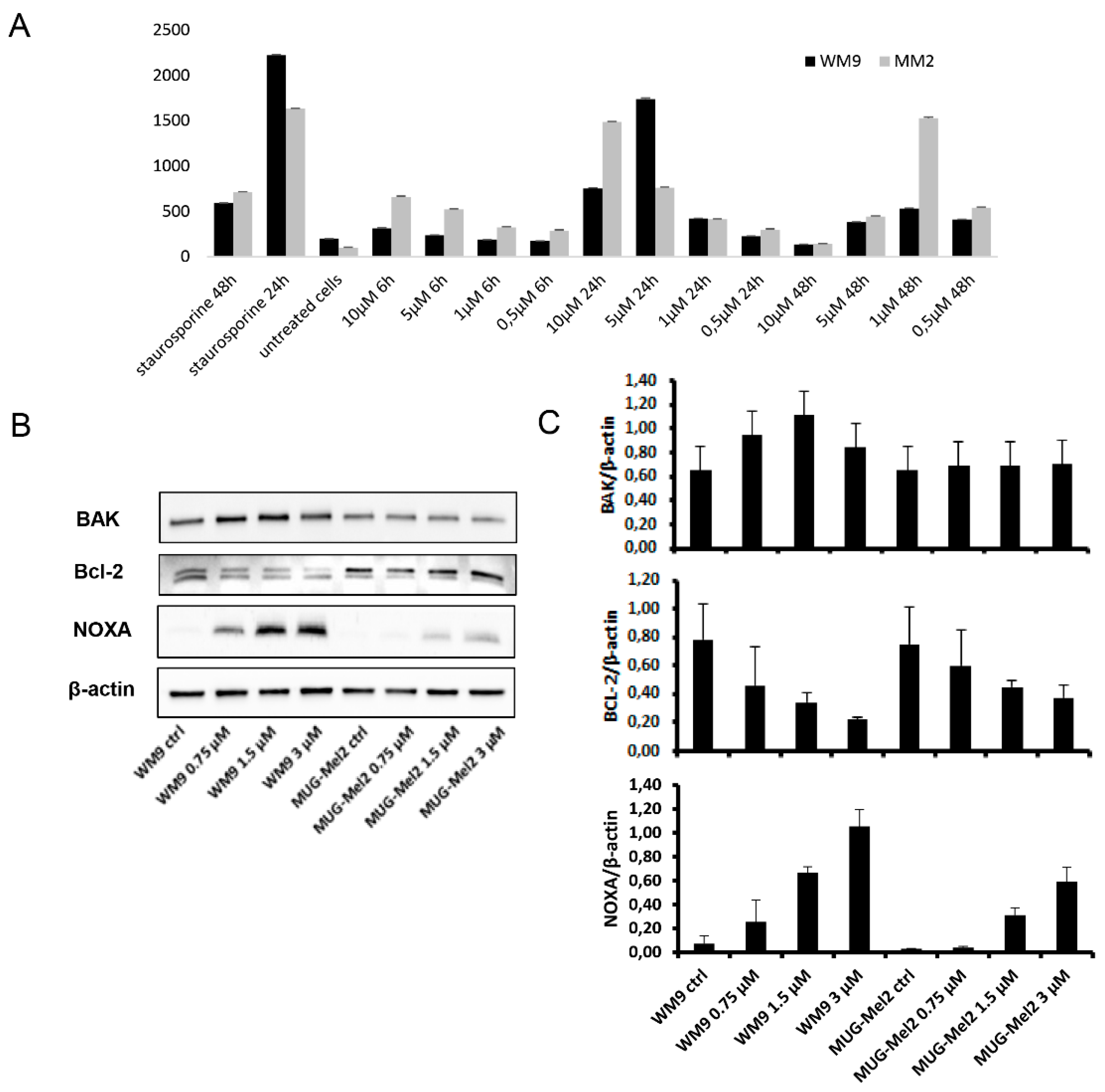
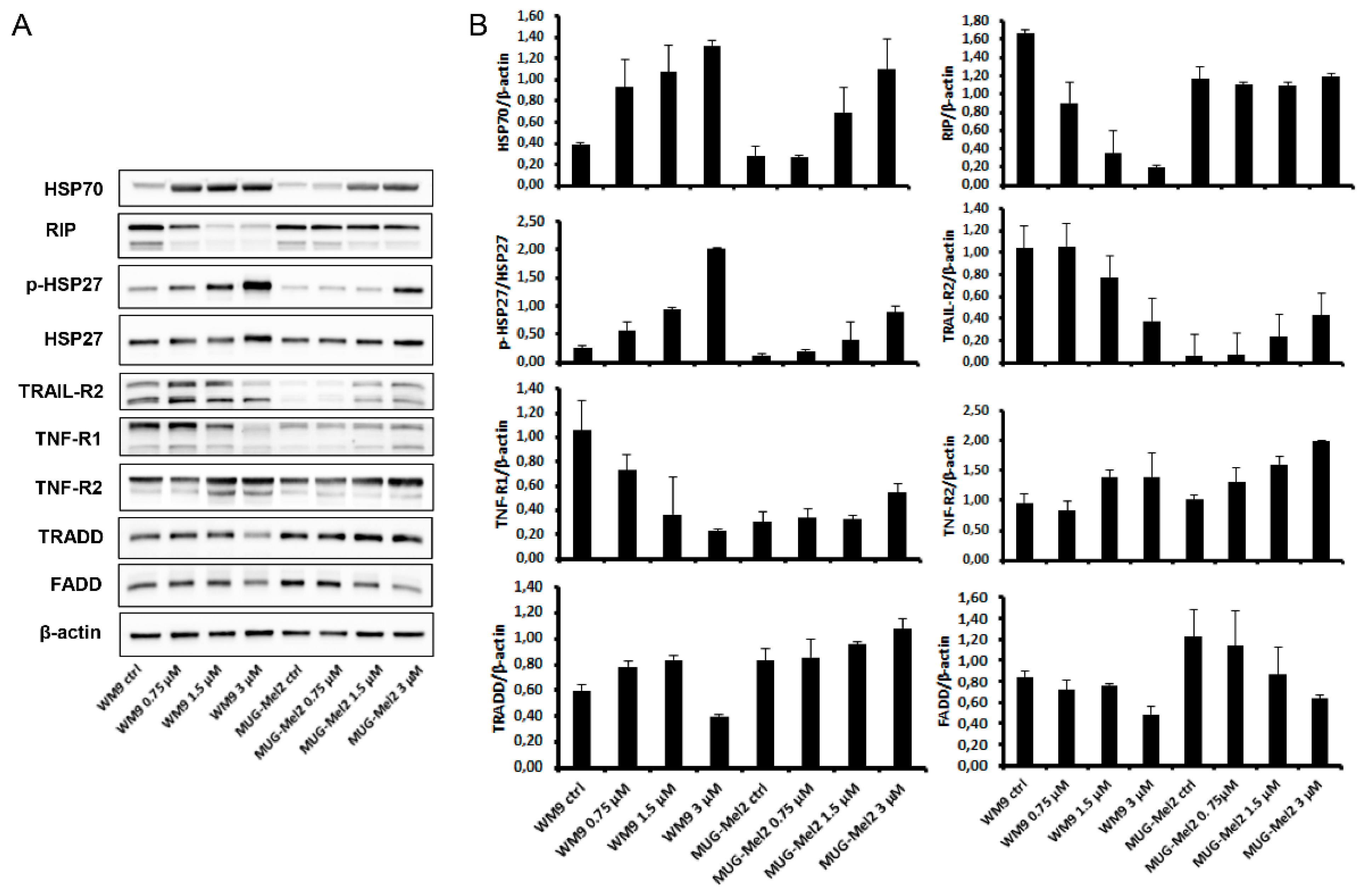
2.3. SK119 Treatment Changed Expression Levels of Apoptosis Genes and Death Receptor Expression
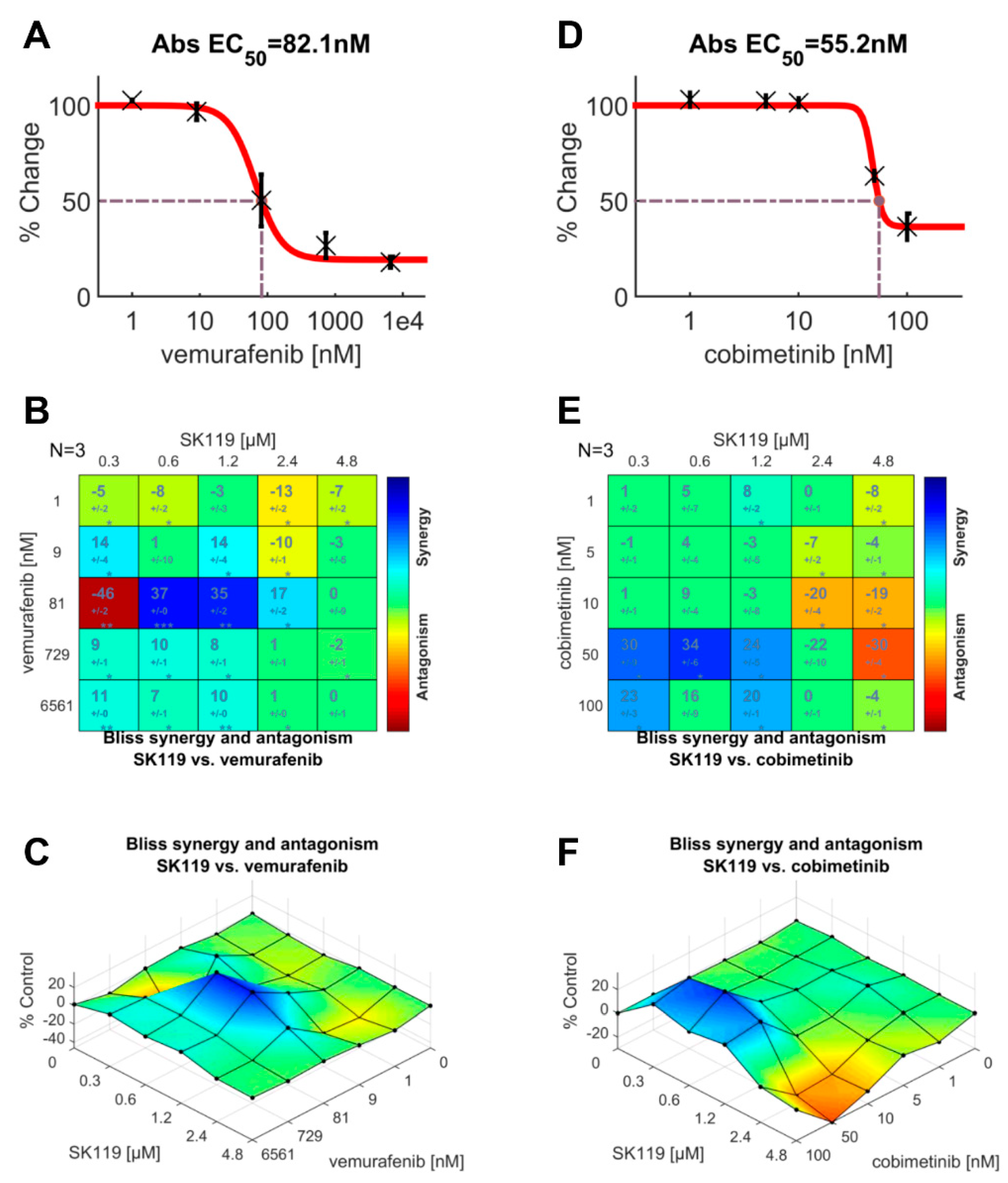
2.4. SK119 Exhibited Synergistic Effects with Vemurafenib and Cobimetinib
3. Materials and Methods
3.1. Synthesis of 1,4-Dihydro-[2-(1-(1,4-dihydro-5,8-dihydroxy-1,4-dioxonaphthalen-3-yl)-4-methylpent-3-enyl)]-5,8-dihydroxy-3-((E)-4-methylpenta-1,3-dienyl)naphthalene-1,4-dione (SK119)
3.2. Spectroscopic Data of SK119
3.3. Cell Culture
3.4. Preparation of Test Compounds
3.5. Cell Viability Assay
3.6. Caspase-Glo® 3/7 Assay
3.7. Western Blot Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuphal, S.; Bosserhoff, A. Recent progress in understanding the pathology of malignant melanoma. J. Pathol. 2009, 219, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.R. The influence of climate change on skin cancer incidence—A review of the evidence. Int. J. Women’s Dermatol. 2020, 7, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.E.; Shalin, S.C.; Tackett, A.J. Current state of melanoma diagnosis and treatment. Cancer Biol. Ther. 2019, 20, 1366–1379. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.E.; Wani, M.C. Camptothecin. Discovery to clinic. Ann. N. Y. Acad. Sci. 1996, 803, 1–12. [Google Scholar] [CrossRef]
- Bailly, C. Irinotecan: 25 years of cancer treatment. Pharmacol. Res. 2019, 148, 104398. [Google Scholar] [CrossRef]
- Thomas, A.; Pommier, Y. Targeting Topoisomerase I in the Era of Precision Medicine. Clin. Cancer Res. 2019, 25, 6581–6589. [Google Scholar] [CrossRef]
- Rinner, B.; Kretschmer, N.; Knausz, H.; Mayer, A.; Boechzelt, H.; Hao, X.-J.; Heubl, G.; Efferth, T.; Schaider, H.; Bauer, R. A petrol ether extract of the roots of Onosma paniculatum induces cell death in a caspase dependent manner. J. Ethnopharmacol. 2010, 129, 182–188. [Google Scholar] [CrossRef]
- Kretschmer, N.; Rinner, B.; Deutsch, A.; Lohberger, B.; Knausz, H.; Kunert, O.; Blunder, M.; Boechzelt, H.; Schaider, H.; Bauer, R. Naphthoquinones from Onosma paniculata induce cell-cycle arrest and apoptosis in melanoma Cells. J. Nat. Prod. 2012, 75, 865–869. [Google Scholar] [CrossRef]
- Kretschmer, N.; Deutsch, A.; Durchschein, C.; Rinner, B.; Stallinger, A.; Higareda-Almaraz, J.C.; Scheideler, M.; Lohberger, B.; Bauer, R. Comparative Gene Expression Analysis in WM164 Melanoma Cells Revealed That β-β-Dimethylacrylshikonin Leads to ROS generation, Loss of Mitochondrial Membrane Potential, and Autophagy Induction. Molecules 2018, 23, 2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stallinger, A.; Kretschmer, N.; Kleinegger, F.; Brvar, L.; Liegl-Atzwanger, B.; Prokesch, A.; Durchschein, C.; Bauer, R.; Deutsch, A.; Rinner, B. β,β-Dimethylacrylshikonin Induces Apoptosis in Melanoma Cell Lines by NOXA Upregulation. J. Nat. Prod. 2020, 83, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Zhongzhen, Z. An Illustrated Chinese Materia Medica in Hong Kong; Hong Kong School of Chinese Medicine: Hong Kong, China, 2004. [Google Scholar]
- Durchschein, C.; Hufner, A.; Rinner, B.; Stallinger, A.; Deutsch, A.; Lohberger, B.; Bauer, R.; Kretschmer, N. Synthesis of novel shikonin derivatives and pharmacological effects of cyclopropylacetylshikonin on melanoma cells. Molecules 2018, 23, 2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretschmer, N.; Hufner, A.; Durchschein, C.; Popodi, K.; Rinner, B.; Lohberger, B.; Bauer, R. Synthesis and pharmacological in vitro investigations of novel shikonin derivatives with a special focus on cyclopropane bearing derivatives. Int. J. Mol. Sci. 2021, 22, 2774. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Zhou, J.; Xu, Z.; Ye, T. Concise Total Synthesis of Nannocystin A. Angew. Chem. Int. Ed. 2016, 55, 13263–13266. [Google Scholar] [CrossRef] [PubMed]
- Meselhy, M.R.; Kadota, S.; Tsubono, K.; Hattori, M.; Nambaa, T. Biotransformation of Shikonin by Human Intestinal Bacteria. Tetrahedron 1994, 50, 3081–3098. [Google Scholar] [CrossRef]
- Meselhy, M.R.; Kadota, S.; Tsubono, K.; Kusai, A.; Hattori, M.; Namba, T. Shikometabolins A, B, C, and D, novel dimeric naphthoquinones metabolites obtained from shikonin by human intestinal bacteria. Tetrahedron Lett. 1994, 35, 583–586. [Google Scholar] [CrossRef]
- Pollock, P.; Harper, U.L.; Hansen, K.S.; Yudt, L.M.; Stark, M.; Robbins, C.M.; Moses, T.Y.; Hostetter, G.; Wagner, U.; Kakareka, J.; et al. High frequency of BRAF mutations in nevi. Nat. Genet. 2003, 33, 19–20. [Google Scholar] [CrossRef]
- Tsao, H.; Chin, L.; Garraway, L.A.; Fisher, D.E. Melanoma: From mutations to medicine. Genes Dev. 2012, 26, 1131–1155. [Google Scholar] [CrossRef] [Green Version]
- Mackiewicz, J.; Mackiewicz, A. BRAF and MEK Inhibitors in the Era of Immunotherapy in Melanoma Patients. Contemp. Oncol. 2018, 22, 68–72. [Google Scholar] [CrossRef]
- Rinner, B.; Gandolfi, G.; Meditz, K.; Frisch, M.-T.; Wagner, K.; Ciarrocchi, A.; Torricelli, F.; Koivuniemi, R.; Niklander, J.; Liegl-Atzwanger, B.; et al. MUG-Mel2, a novel highly pigmented and well characterized NRAS mutated human melanoma cell line. Sci. Rep. 2017, 7, 2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Couselo, E.; Adelantado, E.Z.; Ortiz, C.; Soberino-García, J.; Perez-Garcia, J. NRAS-Mutant Melanoma: Current Challenges and Future Prospect. OncoTargets Ther. 2017, 10, 3941–3947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Assimopoulou, A.N.; Papageorgiou, V.P.; Kolodziej, H. Structure/antileishmanial activity relationship study of naphthoquinones and dependency of the mode of action on the substitution patterns. Planta Med. 2011, 77, 2003–2012. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhao, C.; Yuan, K.; Zhou, G.; Wang, Y.; Xiao, Y.; Wang, C.; Xu, J.; Yang, W. Two new dimeric naphthoquinones with neuraminidase inhibitory activity from Lithospermum erythrorhizon. Nat. Prod. Res. 2015, 29, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Liu, D.; Li, Y.H.; Chen, X.Q.; Luo, K.; Zhang, Y.M.; Li, R.T. Naphthoquinones from Onosma paniculata with potential anti-inflammatory activity. Planta Med. 2017, 83, 631–635. [Google Scholar] [PubMed]
- Cao, H.; Zhang, W.; Liu, D.; Hou, M.; Liu, S.; He, W.; Lin, J.; Shao, M. Identification, in vitro evaluation and modeling studies of the constituents from the roots of Arnebia euchroma for antitumor activity and STAT3 inhibition. Bioorg. Chem. 2020, 96, 103655. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Meselhy, M.R.; Hattori, M.; Kim, H.M.; Kim, Y.H. Cytotoxicity of shikonin metabolites with biotransformation of human intestinal bacteria. J. Microbiol. Biotechnol. 2000, 10, 514–517. [Google Scholar]
- Guo, C.; He, J.; Song, X.; Tan, L.; Wang, M.; Jiang, P.; Li, Y.; Cao, Z.; Peng, C. Pharmacological properties and derivatives of shikonin—A review in recent years. Pharmacol. Res. 2019, 149, 104463. [Google Scholar] [CrossRef]
- Zhang, X.D.; Gillespie, S.K.; Hersey, P. Staurosporine induces apoptosis of melanoma by both caspase-dependent and -independent apoptotic pathways. Mol. Cancer Ther. 2004, 3, 187–197. [Google Scholar]
- Thorburn, A. Death receptor-induced cell killing. Cell. Signal. 2004, 16, 139–144. [Google Scholar] [CrossRef]
- Schulze-Osthoff, K.; Ferrari, D.; Los, M.; Wesselborg, S.; Peter, M.E. Apoptosis signaling by death receptors. Eur. J. Biochem. 1998, 254, 439–459. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, L.A.; Weber, R.F.; Figari, I.S.; Reynolds, C.; Palladino, M.A.; Goeddel, D.V. The two different receptors for tumor necrosis factor mediate distinct cellular responses. Proc. Natl. Acad. Sci. USA 1991, 88, 9292–9296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jego, G.; Hazoumé, A.; Seigneuric, R.; Garrido, C. Targeting heat shock proteins in cancer. Cancer Lett. 2013, 332, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Bruey, J.-M.; Ducasse, C.; Bonniaud, P.; Ravagnan, L.; Susin, S.A.; Diaz-Latoud, C.; Gurbuxani, S.; Arrigo, A.P.; Kroemer, G.; Solary, E.; et al. Hsp27 negatively regulates cell death by interacting with cytochrome C. Nat. Cell Biol. 2000, 2, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Brunet, M.; Didelot, C.; Zermati, Y.; Schmitt, E.; Kroemer, G. Heat shock proteins 27 and 70: Anti-apoptotic proteins with tumorigenic properties. Cell Cycle 2006, 5, 2592–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponti, G.; Manfredini, M.; Greco, S.; Pellacani, G.; Depenni, R.; Tomasi, A.; Maccaferri, M.; Cascinu, S. BRAF, NRAS and C-KIT Advanced Melanoma: Clinico-pathological Features, Targeted-Therapy Strategies and Survival. Anticancer Res. 2017, 37, 7043–7048. [Google Scholar]
- Davies, M.A.; Liu, P.; McIntyre, S.; Kim, K.B.; Papadopoulos, N.; Hwu, W.-J.; Hwu, P.; Bedikian, A. Prognostic factors for survival in melanoma patients with brain metastases. Cancer 2012, 117, 1687–1696. [Google Scholar] [CrossRef]
- Luke, J.J.; Hodi, F.S. Ipilimumab, vemurafenib, dabrafenib, and trametinib: Synergistic competitors in the clinical management of BRAF mutant malignant melanoma. Oncologist 2013, 18, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.B.; Puzanov, I. Treatment of NRAS-mutant melanoma. Curr. Treat. Options Oncol. 2015, 16, 15. [Google Scholar] [CrossRef]
- Di Veroli, G.Y.; Fornari, C.; Wang, D.; Mollard, S.; Bramhall, J.L.; Richards, F.M.; Jodrell, D.I. Combenefit: An interactive platform for the analysis and visualization of drug combinations. Bioinformatics 2016, 32, 2866–2868. [Google Scholar] [CrossRef]
- Broman, K.K.; Dossett, L.A.; Sun, J.; Eroglu, Z.; Zager, J.S. Update on BRAF and MEK inhibition for treatment of melanoma in metastatic, unresectable, and adjuvant settings. Expert Opin. Drug Saf. 2019, 18, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.F.; Saad, H.-E.A.; El-Karim, E.M.A. Two Polycyclic Geranylhydroquinone-Derived Metabolites from Roots of Arnebia hispidissima (Lehm.) DC. Molecules 2014, 19, 5940–5951. [Google Scholar] [CrossRef] [PubMed]
- Assimopoulou, A.N.; Ganzera, M.; Stuppner, H.; Papageorgiou, V.P. Simultaneous determination of monomeric and oligomeric alkannins and shikonins by high-performance liquid chromatography–diode array detection–mass spectrometry. Biomed. Chromatogr. 2008, 22, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yu, P.; Zhou, Q.; Tian, Z.; Sun, M.; Li, X.; Wang, X.; Jiang, H. An integrated data filtering and identification strategy for rapid profiling of chemical constituents, with Arnebiae Radix as an example. J. Chrom. A 2020, 1629, 461496. [Google Scholar] [CrossRef]
- Liao, M.; Li, A.; Chen, C.; Ouyang, H.; Zhang, Y.; Xu, Y.; Feng, Y.; Jiang, H. Systematic identification of shikonins and shikonofurans in medicinal Zicao species using ultra-high performance liquid chromatography quadrupole time of flight tandem mass spectrometry combined with a data mining strategy. J. Chrom. A 2015, 1425, 158–172. [Google Scholar] [CrossRef]
- Liao, M.; Yan, P.; Liu, X.; Du, Z.; Jia, S.; Aybek, R.; Li, A.; Kaisa, S.; Jiang, H. Spectrum-effect relationship for anti-tumor activity of shikonins and shikonofurans in medicinal Zicao by UHPLC-MS/MS and chemometric approaches. J. Chrom. B 2020, 1136, 121924. [Google Scholar] [CrossRef]
- Noula, E.; Samanidou, V.F.; Assimopoulou, A.N.; Papageorgiou, V.P.; Papadoyannis, I.N. Solid-phase extraction for purification of alkannin/shikonin samples and isolation of monomeric and dimeric fractions. Anal. Bioanal. Chem. 2010, 397, 2221–2232. [Google Scholar] [CrossRef]
- Spyros, A.; Assimopoulou, A.N.; Papageorgiou, V.P. Structure determination of oligomeric alkannin and shikonin derivatives. Biomed. Chromatogr. 2005, 19, 498–505. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent CDCl3 | Solvent C6D6 | |||
|---|---|---|---|---|
| No. | 13C | 1H | 13C | 1H |
| 1 | 186.8 | - | 187.3 | - |
| 2 | 143.9? | - | 144.1 | - |
| 3 | 143.9 | - | 144.7 | - |
| 4 | 186.2 | - | 187.0 | - |
| 5 | 158.3 ** | OH: 12.72 s | 159.4 | OH: 13.10 s |
| 6 | 129.6 *** | 7.20 s | 130.1 | 6.70 d, J = 9.1 Hz |
| 7 | 129.2 *** | 7.20 s | 129.8 | 6.69 d, J = 9.1 Hz |
| 8 | 158.6 ** | OH: 12.72 s | 159.6 | OH: 13.03 s |
| 9 | 112.1 * | - | 112.8 | - |
| 10 | 111.80 * | - | 112.52 * | - |
| 11 | 120.6 | 6.58 d, J = 15.6 Hz | 121.8 | 6.76 d, J = 15.3 Hz |
| 12 | 139.2 | 7.41 dd, J = 15.6, 11.3 Hz | 139.9 | 7.65 dd, J = 15.3, 11.1 Hz |
| 13 | 126.7 | 6.04 d, J = 11.2 Hz | 127.8 | 6.03 d, J = 11.4 Hz |
| 14 | 143.5 | - | 143.4 | - |
| 15 | 26.5 | 1.63 s | 26.6 | 1.58 s |
| 16 | 19.0 | 1.60 s | 19.1 | 1.60 s |
| 1′ | 182.5 | - | 182.79 ** | - |
| 2′ | 151.4 | - | 151.6 | - |
| 3′ | 135.7 | 7.23 d, J = 1.1 Hz | 136.3 | 7.26 d, J = 0.9 Hz |
| 4′ | 182.7 | - | 182.72 ** | - |
| 5′ | 162.5 | OH: 12.47 s | 163.6 | OH: 12.85 s |
| 6′ | 131.2 | 7.19 d, J = 9.6 Hz | 131.8 | 6.59 d, J = 9.7 Hz |
| 7′ | 131.0 | 7.16 d, J = 9.6 Hz | 131.2 | 6.53 d, J = 9.7 Hz |
| 8′ | 163.1 | OH: 12.41 s | 164.2 | OH: 12.71 s |
| 9′ | 111.77 * | - | 112.2 | - |
| 10′ | 111.6 | - | 112.48 * | - |
| 11′ | 38.8 | 4.67 tm, J = 7.6 Hz | 39.6 | 4.72 tm, J = 7.0 Hz |
| 12′ | 29.7 | 2.73–2.87 m, 2H | 30.6 | 2.62–2.79 m, 2H |
| 13′ | 120.3 | 5.08 tm, J = 6.8 Hz | 121.5 | 5.14 tm, J = 7.0 Hz |
| 14′ | 135.2 | - | 135.2 | - |
| 15′ | 25.7 | 1.89 s | 26.1 | 1.53 s |
| 16′ | 18.0 | 1.85 s | 18.3 | 1.51 s |
| Cell Line | Sbcl2 | WM9 | WM164 | MUG-Mel2 | FS-1 | HEK-293 |
|---|---|---|---|---|---|---|
| SK119 | 0.3 ± 0.01 | 0.6 ± 0.01 | 1.7 ± 0.3 | 0.7 ± 0.01 | 0.5 ± 0.01 | 2.2 ± 0.4 |
| DMAS | 1.1 ± 0.1 | 2.7 ± 0.3 | 8.3 ± 0.3 | 7.2 ± 0.5 | 1.8 ± 0.3 | 9.5 ± 1.4 |
| x-fold ∆ | 3.7 | 4.5 | 4.9 | 10.3 | 3.7 | 4.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kretschmer, N.; Durchschein, C.; Hufner, A.; Rinner, B.; Lohberger, B.; Bauer, R. SK119, a Novel Shikonin Derivative, Leads to Apoptosis in Melanoma Cell Lines and Exhibits Synergistic Effects with Vemurafenib and Cobimetinib. Int. J. Mol. Sci. 2022, 23, 5684. https://doi.org/10.3390/ijms23105684
Kretschmer N, Durchschein C, Hufner A, Rinner B, Lohberger B, Bauer R. SK119, a Novel Shikonin Derivative, Leads to Apoptosis in Melanoma Cell Lines and Exhibits Synergistic Effects with Vemurafenib and Cobimetinib. International Journal of Molecular Sciences. 2022; 23(10):5684. https://doi.org/10.3390/ijms23105684
Chicago/Turabian StyleKretschmer, Nadine, Christin Durchschein, Antje Hufner, Beate Rinner, Birgit Lohberger, and Rudolf Bauer. 2022. "SK119, a Novel Shikonin Derivative, Leads to Apoptosis in Melanoma Cell Lines and Exhibits Synergistic Effects with Vemurafenib and Cobimetinib" International Journal of Molecular Sciences 23, no. 10: 5684. https://doi.org/10.3390/ijms23105684
APA StyleKretschmer, N., Durchschein, C., Hufner, A., Rinner, B., Lohberger, B., & Bauer, R. (2022). SK119, a Novel Shikonin Derivative, Leads to Apoptosis in Melanoma Cell Lines and Exhibits Synergistic Effects with Vemurafenib and Cobimetinib. International Journal of Molecular Sciences, 23(10), 5684. https://doi.org/10.3390/ijms23105684