
Impact of Chemical Endocrine Disruptors and Hormone Modulators on the Endocrine System


 and
and 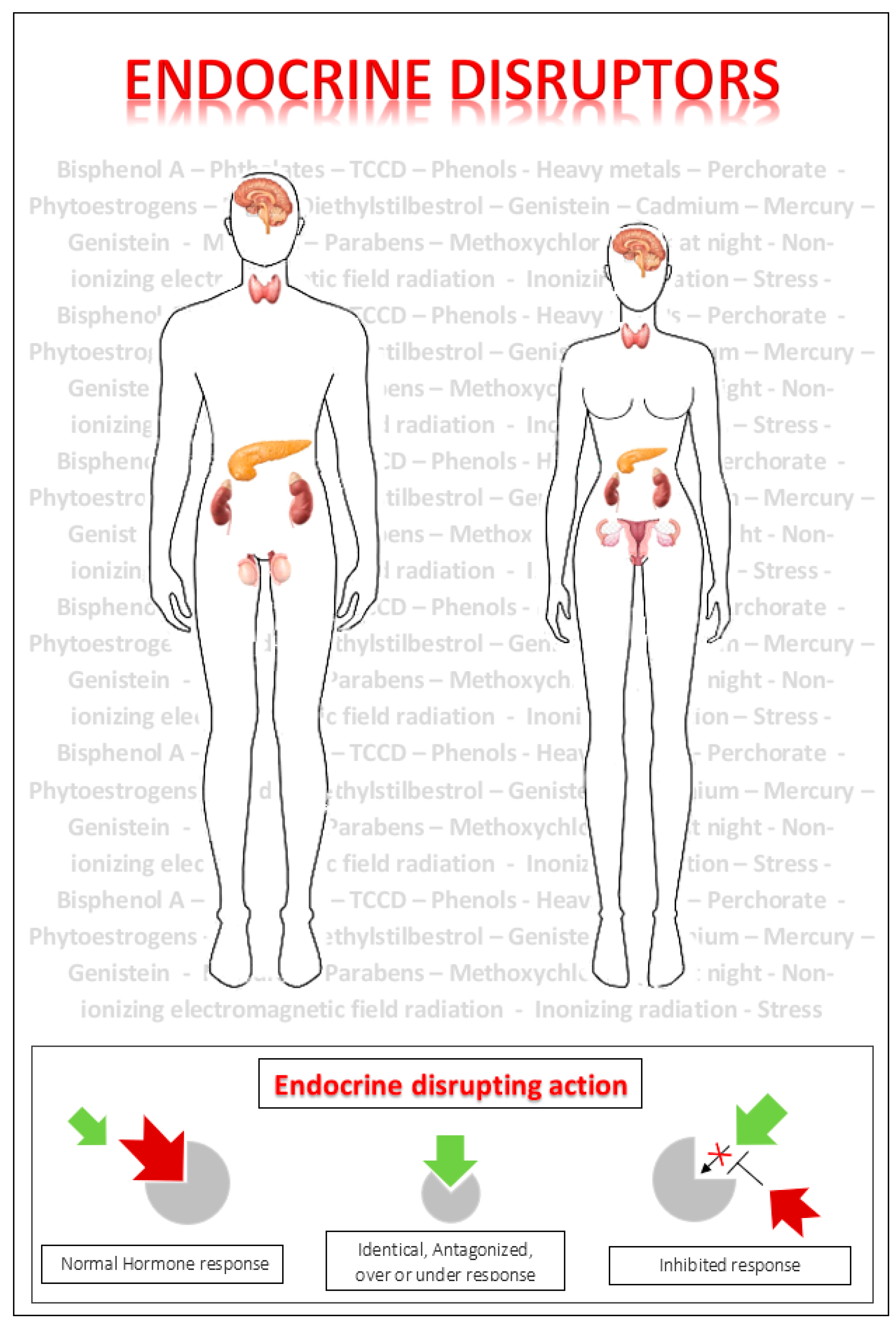{kind=link}
Abstract
:1. Introduction
2. Thyroid and Parathyroid Glands
3. Adrenal Glands
4. Pancreatic Beta-Cells
5. Testes
6. Ovaries
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Committee ES. Scientific Opinion on the hazard assessment of endocrine disruptors: Scientific criteria for identification of endocrine disruptors and appropriateness of existing test methods for assessing effects mediated by these substances on human health and the environment. EFSA J. 2013, 11, 3132. [Google Scholar]
- Slama, R.; Bourguignon, J.P.; Demeneix, B.; Ivell, R.; Panzica, G.; Kortenkamp, A.; Zoeller, R.T. Scientific issues relevant to setting regulatory criteria to identify endocrine-disrupting substances in the European Union. Environ. Health Perspect. 2016, 124, 1497–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfo, A.; Nuzzo, A.M.; De Amicis, R.; Moretti, L.; Bertoli, S.; Leone, A. Fetal-Maternal Exposure to Endocrine Disruptors: Correlation with Diet Intake and Pregnancy Outcomes. Nutrients 2020, 12, 1744. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, J.; Park, J.; Kim, H.J.; Cho, G.; Kim, G.H.; Eun, S.H.; Lee, J.J.; Choi, G.; Suh, E.; et al. Concentrations of phthalate metabolites in breast milk in Korea: Estimating exposure to phthalates and potential risks among breast-fed infants. Sci. Total Environ. 2015, 508, 13–19. [Google Scholar] [CrossRef]
- Di Nisio, A.; Foresta, C. Water and soil pollution as determinant of water and food quality/contamination and its impact on male fertility. Reprod. Biol. Endocrinol. 2019, 17, 4. [Google Scholar] [CrossRef]
- De Coster, S.; van Larebeke, N. Endocrine-disrupting chemicals: Associated disorders and mechanisms of action. J. Environ. Public Health 2012, 2012, 713696. [Google Scholar] [CrossRef]
- Russart, K.L.G.; Nelson, R.J. Light at night as an environmental endocrine disruptor. Physiol. Behav. 2018, 190, 82–89. [Google Scholar] [CrossRef]
- Marci, R.; Mallozzi, M.; Di Benedetto, L.; Schimberni, M.; Mossa, S.; Soave, I.; Palomba, S.; Caserta, D. Radiations and female fertility. Reprod. Biol. Endocrinol. 2018, 16, 112. [Google Scholar] [CrossRef]
- Palomba, S.; Daolio, J.; Romeo, S.; Battaglia, F.A.; Marci, R.; La Sala, G.B. Lifestyle and fertility: The influence of stress and quality of life on female fertility. Reprod. Biol. Endocrinol. 2018, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine disrupting chemicals: Exposure, effects on human health, mechanism of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147. [Google Scholar] [CrossRef]
- Casals-Casas, C.; Desvergne, B. Endocrine Disruptors: From Endocrine to Metabolic Disruption. Annu. Rev. Physiol. 2011, 73, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamanti-Kandarakis, E.; Palioura, E.; Kandarakis, S.A.; Koutsilieris, M. The impact of endocrine disruptors on endocrine targets. Horm. Metab. Res. 2010, 42, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Parker, V.G.; Mayo, R.M.; Logan, B.N.; Holder, B.J.; Smart, P.T. Toxins and diabetes mellitus: An environmental connection? Diabetes Spectr. 2002, 15, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Biemann, R.; Blüher, M.; Isermann, B. Best Exposure to endocrine-disrupting compounds such as phthalates and bisphenol A is associated with an increased risk for obesity. Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101546. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fang, R.; Wang, H.; Xu, D.X.; Yang, J.; Huang, X.; Cozzolino, D.; Fang, M.; Huang, Y. A review of environmental metabolism disrupting chemicals and effect biomarkers associating disease risks: Where exposomics meets metabolomics. Environ. Int. 2022, 158, 106941. [Google Scholar] [CrossRef]
- Predieri, B.; Alves, C.A.D.; Iughetti, L. New insights on the effects of endocrine-disrupting chemicals on children. J. Pediatr. (Rio J.) 2021, 15, S73–S85. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, I.K.; Porta, M.; Steffes, M.; Jacobs, D.R., Jr. Relationship between serum concentrations of persistent organic pollutants and the prevalence of metabolic syndrome among nondiabetic adults: Results from the National Health and Nutrition Examination Survey 1999–2002. Diabetologia 2007, 50, 1841–1851. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K.; Son, H.K.; Lee, S.K.; Kang, J.H.; Chang, Y.S.; Jacobs, D.R.; Lee, D.H. Relationship between serum concentrations of organochlorine pesticides and metabolic syndrome among non-diabetic adults. J. Prev. Med. Public Health 2010, 43, 1–8. [Google Scholar] [CrossRef]
- Sargis, R.M.; Simmons, R.A. Environmental neglect: Endocrine disruptors as underappreciated but potentially modifiable diabetes risk factors. Diabetologia 2019, 62, 1811–1822. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S.; Pedersen, K.M.; Bruun, N.H.; Laurberg, P. Narrow individual variations in serum T(4) and T(3) in nor- mal subjects: A clue to the understanding of subclinical thyroid disease. J. Clin. Endocrinol. Metab. 2002, 87, 1068–1072. [Google Scholar] [CrossRef]
- Andersen, S.; Bruun, N.H.; Pedersen, K.M.; Laurberg, P. Biologic variation is important for interpretation of thyroid function tests. Thyroid 2003, 13, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Fekete, C.; Lechan, R.M. Central regulation of hypotha- lamic-pituitary-thyroid axis under physiological and pathophysiological conditions. Endocr. Rev. 2014, 35, 159–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, M.E. Impact of low-level thyroid hormone dis- ruption induced by propylthiouracil on brain develop- ment and function. Toxicol. Sci. 2011, 124, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Zoeller, T.R. Environmental chemicals targeting thyroid. Hormones 2010, 9, 28–40. [Google Scholar] [CrossRef]
- Triggiani, V.; Tafaro, E.; Giagulli, V.A.; Sabbà, C.; Resta, F.; Licchelli, B.; Guastamacchia, E. Role of iodine, selenium and other micronutrients in thyroid function and disorders. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 277–294. [Google Scholar] [CrossRef]
- Burniat, A.; Pirson, I.; Vilain, C.; Kulik, W.; Afink, G.; Moreno-Reyes, R.; Corvilain, B.; Abramowicz, M. Iodotyrosine deiodi- nase defect identified via genome-wide approach. J. Clin. Endocrinol. Metab. 2012, 97, E1276–E1283. [Google Scholar] [CrossRef] [Green Version]
- Dumitrescu, A.M.; Refetoff, S. Inherited defects of thyroid hormone metabolism. Ann. Endocrinol. 2011, 72, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Council on Environmental Health; Rogan, W.J.; Paulson, J.A.; Baum, C.; Brock-Utne, A.C.; Brumberg, H.L.; Campbell, C.C.; Lanphear, B.P.; Lowry, J.A.; Osterhoudt, K.C.; et al. Iodine deficiency, pollutant chemicals, and the thyroid: New information on an old problem. Pediatrics 2014, 133, 1163–1166. [Google Scholar]
- Taylor, P.N.; Okosieme, O.E.; Murphy, R.; Hales, C.; Chiusano, E.; Maina, A.; Joomun, M.; Bestwick, J.P.; Smyth, P.; Paradice, R.; et al. Maternal perchlorate levels in women with borderline thyroid func- tion during pregnancy and the cognitive development of their offspring: Data from the Controlled Antenatal Thyroid Study. J. Clin. Endocrinol. Metab. 2014, 99, 4291–4298. [Google Scholar] [CrossRef]
- Köhrle, J. Environment and endocrinology: The case of thyroidology. Ann. Endocrinol. 2008, 69, 116–122. [Google Scholar] [CrossRef]
- Zoeller, R.T.; Tyl, R.W.; Tan, S.W. Current and potential rodent screens and tests for thyroid toxicants. Crit. Rev. Toxicol. 2007, 37, 55–95. [Google Scholar] [CrossRef] [PubMed]
- Howdeshell, K.L. A model of the development of the brain as a construct of the thyroid system. Environ. Health Perspect. 2002, 110, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeker, J.D.; Ferguson, K.K. Relationship between urinary phthalate and bisphenol A concentrations and serum thyroid measures in U.S. adults and adolescents from the National Health and Nutrition Examination Survey (NHANES) 2007–2008. Environ. Health Perspect. 2011, 119, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Montaño, M.; Cocco, E.; Guignard, C.; Marsh, G.; Hoffmann, L.; Bergman, A.; Gutleb, A.C.; Murk, A.J. New approaches to assess the transthyretin binding capacity of bioactivated thyroid hormone disruptors. Toxicol. Sci. 2012, 130, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Guo, L.H.; Wan, B.; Wei, Y. In vitro fluorescence displacement investigation of thyroxine transport disruption by bisphenol A. J. Environ. Sci. 2011, 23, 315–321. [Google Scholar] [CrossRef]
- Gutleb, A.C.; Cenijn, P.; van Velzen, M.; Lie, E.; Ropstad, E.; Skaare, J.U.; Malmberg, T.; Bergman, A.; Gabrielsen, G.W.; Legler, J. In vitro assay shows that PCB metabolites completely saturate thyroid hormone transport capacity in blood of wild polar bears (Ursus maritimus). Environ. Sci. Technol. 2010, 44, 3149–3154. [Google Scholar] [CrossRef]
- Cao, J.; Lin, Y.; Guo, L.H.; Zhang, A.Q.; Wei, Y.; Yang, Y. Structure-based investigation on the binding interaction of hydroxylated polybrominated diphenyl ethers with thyroxine transport proteins. Toxicology 2010, 277, 20–28. [Google Scholar] [CrossRef]
- Weiss, J.M.; Andersson, P.L.; Lamoree, M.H.; Leonards, P.E.; van Leeuwen, S.P.; Hamers, T. Competitive binding of poly- and perfluorinated compounds to the thyroid hormone transport protein transthyretin. Toxicol. Sci. 2009, 109, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Marchesini, G.R.; Meimaridou, A.; Haasnoot, W.; Meulenberg, E.; Albertus, F.; Mizuguchi, M.; Takeuchi, M.; Irth, H.; Murk, A.J. Biosensor discovery of thyroxine transport disrupting chemicals. Toxicol. Appl. Pharmacol. 2008, 232, 150–160. [Google Scholar] [CrossRef]
- Roques, B.B.; Leghait, J.; Lacroix, M.Z.; Lasserre, F.; Pineau, T.; Viguié, C.; Martin, P.G.P. The nuclear receptors pregnane X receptor and constitutive androstane receptor contribute to the impact of fipronil on hepatic gene expression linked to thyroid hormone metabolism. Biochem. Pharmacol. 2013, 86, 997–1039. [Google Scholar] [CrossRef]
- Greer, M.A.; Goodman, G.; Pleus, R.C.; Greer, S.E. Health effects assessment for environmental perchlorate contamination: The dose response for inhibition of thyroidal radioiodine uptake in humans. Environ. Health Perspect. 2002, 110, 927–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsberg, G.L.; Hattis, D.B.; Zoeller, R.T.; Rice, D.C. Evaluation of the U.S. EPA/OSWER preliminary remediation goal for perchlorate in groundwater: Focus on exposure to nursing infants. Environ. Health Perspect. 2007, 115, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoeller, R.T.; Rovet, J. Timing of thyroid hormone action in the developing brain: Clinical observations and experimental findings. J. Neuroendocrinol. 2004, 16, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Pearce, E.N.; Leung, A.M.; Blount, B.C.; Bazrafshan, H.R.; He, X.; Pino, S.; Valentin-Blasini, L.; Braverman, L.E. Breast milk iodine and perchlorate concentrations in lactating Boston- area women. J. Clin. Endocrinol. Metab. 2007, 92, 1673–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schantz, S.L.; Widholm, J.J.; Rice, D.C. Effects of PCB exposure on neuropsychological function in children. Environ. Health Perspect. 2003, 111, 357–576. [Google Scholar] [CrossRef] [Green Version]
- Herbstman, J.B.; Sjödin, A.; Kurzon, M.; Lederman, S.A.; Jones, R.S.; Rauh, V.; Needham, L.L.; Tang, D.; Niedzwiecki, M.; Wang, R.Y.; et al. Prenatal exposure to PBDEs and neurodevelopment. Environ. Health Perspect. 2010, 118, 712–719. [Google Scholar] [CrossRef]
- Huang, P.C.; Kuo, P.L.; Guo, Y.L.; Liao, P.C.; Lee, C.C. Associations between urinary phthalate monoesters and thyroid hormones in pregnant women. Hum. Reprod. 2007, 22, 2715–2722. [Google Scholar] [CrossRef] [Green Version]
- Frederiksen, M.; Vorkamp, K.; Thomsen, M.; Knudsen, L.E. Human internal and external exposure to PBDEs–a re- view of levels and sources. Int. J. Hyg. Environ. Health 2009, 212, 109–134. [Google Scholar] [CrossRef]
- Costa, L.G.; de Laat, R.; Tagliaferri, S.; Pellacani, C. A mechanistic view of polybrominated diphenyl ether (PBDE) developmental neurotoxicity. Toxicol. Lett. 2014, 230, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Suvorov, A.; Girard, S.; Lachapelle, S.; Abdelouahab, N.; Sebire, G.; Takser, L. Perinatal exposure to low-dose BDE- 47, an emergent environmental contaminant, causes hyperactivity in rat offspring. Neonatology 2009, 95, 203–209. [Google Scholar] [CrossRef]
- Rice, D.C.; Reeve, E.A.; Herlihy, A.; Zoeller, R.T.; Thompson, W.D.; Markowski, V.P. Developmental delays and locomotor activity in the C57BL6/J mouse following neonatal exposure to the fully-brominated PBDE, decabromodi- phenyl ether. Neurotoxicol. Teratol. 2007, 29, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Viberg, H.; Johansson, N.; Fredriksson, A.; Eriksson, J.; Marsh, G.; Eriksson, P. Neonatal exposure to higher brominated diphenyl ethers, hepta-, octa-, or nonabromodiphenyl ether, impairs spontaneous behavior and learning and memory functions of adult mice. Toxicol. Sci. 2006, 92, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, P.; Fischer, C.; Fredriksson, A. Polybrominated diphenyl ethers, a group of brominated flame retardants, can interact with polychlorinated biphenyls in enhancing developmental neurobehavioral defects. Toxicol. Sci. 2006, 94, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeker, J.D.; Calafat, A.M.; Hauser, R. Di(2-ethylhexyl) phthalate metabolites may alter thyroid hormone levels in men. Environ. Health Perspect. 2007, 115, 1029–1034. [Google Scholar] [CrossRef]
- Bansal, R.; Tighe, D.; Danai, A.; Rawn, D.F.K.; Gaertner, D.W.; Arnold, D.L.; Gilbert, M.E.; Zoeller, R.T. Polybrominated di- phenyl ether (DE-71) interferes with thyroid hormone action independent of effects on circulating levels of thyroid hormone in male rats. Endocrinology 2014, 155, 1104–4112. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.T.; Wu, C.F.; Chen, B.H.; Lederman, S.A.; Jones, R.S.; Rauh, V.; Needham, L.L.; Tang, D.; Niedzwiecki, M.; Wang, R.Y.; et al. Intake of phthalate-tainted foods alters thyroid functions in Taiwanese children. PLoS ONE 2013, 8, e55005. [Google Scholar] [CrossRef]
- Ibhazehiebo, K.; Koibuchi, N. Thyroid hormone receptor- mediated transcription is suppressed by low dose phthalate. Niger. J. Physiol. Sci. 2011, 26, 143–149. [Google Scholar]
- Kim, U.J.; Oh, J.E. Tetrabromobisphenol A and hexabro- mocyclododecane flame retardants in infant-mother paired serum samples, and their relationships with thyroid hormones and environmental factors. Environ. Pollut. 2014, 184, 193–200. [Google Scholar] [CrossRef]
- Wang, T.; Lu, J.; Xu, M.; Xu, Y.; Li, M.; Liu, Y.; Tian, X.; Chen, Y.; Dai, M.; Wang, W.; et al. Urinary bisphenol A concentration and thyroid function in Chinese adults. Epidemiology 2013, 24, 295–302. [Google Scholar] [CrossRef]
- Sriphrapradang, C.; Chailurkit, L.O.; Aekplakorn, W.; Ong-phiphadhanakul, B. Association between bisphenol A and abnormal free thyroxine level in men. Endocrine 2013, 44, 441–447. [Google Scholar] [CrossRef]
- Moriyama, K.; Tagami, T.; Akamizu, T.; Usui, T.; Saijo, M.; Kanamoto, N.; Hataya, Y.; Shimatsu, A.; Kuzuya, H.; Nakao, K. Thyroid hormone action is disrupted by bisphenol A as an antagonist. J. Clin. Endocrinol. Metab. 2002, 87, 5185–5190. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Jinno, N.; Ohta, S.; Kuroki, H.; Fujimoto, N. Thyroid hormonal activity of the flame retardants tetra-bromobisphenol A and tetrachlorobisphenol A. Biochem. Biophys. Res. Commun. 2002, 293, 554–559. [Google Scholar] [CrossRef]
- Sheng, Z.G.; Tang, Y.; Liu, Y.X.; Usui, T.; Saijo, M.; Kanamoto, N.; Hataya, Y.; Shimatsu, A.; Kuzuya, H.; Nakao, K. Low concentrations of bisphenol A suppress thyroid hormone receptor transcription through a nongenomic mechanism. Toxicol. Appl. Pharmacol. 2012, 259, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Saunders, N.; Safley, S.; Smith, M.R.; Liang, Y.; Tran, V.; Sharma, J.; Jones, D.P.; Weber, C.J. Environmental chemicals and metabolic disruption in primary and secondary human parathyroid tumors. Surgery 2021, 169, 102–108. [Google Scholar] [CrossRef]
- Harvey, P.W. Adrenocortical endocrine disruption. J. Steroid Biochem. Mol. Biol. 2016, 155, 199–206. [Google Scholar] [CrossRef]
- Ulleras, E.; Ohlsson, A.; Oskarsson, A. Secretion of cortisol and aldosterone as a vulnerable target for adrenal endocrine disruption—Screening of 30 selected chemicals in the human H295R cell model. J. Appl. Toxicol. 2008, 28, 1045–1053. [Google Scholar] [CrossRef]
- Sargis, R.M. Metabolic disruption in context: Clinical avenues for synergistic perturbations in energy homeostasis by endocrine disrupting chemicals. Endocr. Disruptors 2015, 3, e1080788. [Google Scholar] [CrossRef]
- Hampl, R.; Kubatova, J.; Starka, L. Steroids and endocrine disruptors–history, recent state of art and open questions. J. Steroid Biochem. Mol. Biol. 2016, 155, 217–223. [Google Scholar] [CrossRef]
- Wagner, R.L.; White, P.F.; Kan, P.B.; Rosenthal, M.H.; Feldman, D. Inhibition of adrenal steroidogenesis by the anaesthetic etomidate. N. Engl. J. Med. 1984, 310, 1415–1421. [Google Scholar] [CrossRef]
- Cai, W.; Benitez, R.; Counsell, R.E.; Djanegara, T.; Schteingart, D.E.; Sinsheimer, J.E.; Wotring, L.L. Bovine adrenal cortex transformations of mitotane [1-(2-chlorophenyl)-1-(4-chlorophenyl)-2,2- dichloroethane; o,p’-DDD] and its p,p’- and m,p’-isomers. Biochem. Pharmacol. 1995, 49, 1483–1489. [Google Scholar] [CrossRef]
- Johansson, M.K.; Sanderson, J.T.; Lund, B.O. Effects of 3-MeSO2-DDE and some CYP inhibitors on glucocorticoid steroidogenesis in the H295R human adrenocortical carcinoma cell line. Toxicol. Vitr. 2002, 16, 113–121. [Google Scholar] [CrossRef]
- Wang, S.W.; Pu, H.F.; Kan, S.F.; Tseng, C.I.; Lo, M.J.; Wang, P.S. Inhibitory effects of digoxin and digitoxin on corticosterone production in rat zona fasciculata-reticularis cells. Br. J. Pharmacol. 2004, 142, 1123–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jager, L.P.; de Graaf, G.J.; Widjaja-Greefkes, H.C. Differential effects of nitrofurans on the production/release of steroid hormones by porcine adrenocortical cells in vitro. Eur. J. Pharmacol. 1997, 331, 325–331. [Google Scholar] [CrossRef]
- Creange, J.E.; Schane, H.P.; Anzalone, A.J.; Potts, G.O. Interruption of pregnancy in rats by azastene, an inhibitor of ovarian and adrenal steroidogenesis. Fertil. Steril. 1978, 30, 86–90. [Google Scholar] [CrossRef]
- Li, L.-A.; Wang, P.-W.; Chang, L.W. Polychlorinated biphenyl 126 stimulates basal and inducible aldosterone biosynthesis of human adrenocortical H295R cells. Toxicol. Appl. Pharmacol. 2004, 195, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Goodfriend, T.L.; Ball, D.L.; Elliott, M.E.; Shackleton, C. Lead increases aldosterone production by rat adrenal cells. Hypertension 1995, 25, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Hinson, J.P.; Raven, P.W. Effects of endocrine-disrupting chemicals on adrenal function. Best Pract. Res. Clin. Endocrinol. Metab. 2006, 20, 111–120. [Google Scholar] [CrossRef]
- Timokhina, E.P. Dichlorodiphenyltrichloroethane and the Adrenal Gland: From Toxicity to Endocrine Disruption. Toxics 2021, 9, 243. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Obernikhin, S.S.; Yaglov, V.V.; Nazimova, S.V.; Timokhina, E.P.; Tsomartova, D.A. Low-Dose Exposure to Endocrine Disruptor Dichlorodiphenyltrichloroethane (DDT) Affects Transcriptional Regulation of Adrenal Zona Reticularis in Male Rats. Bull. Exp. Biol. Med. 2021, 170, 682–685. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Timokhina, E.P.; Yaglov, V.V.; Obernikhin, S.S.; Nazimova, S.V.; Tsomartova, D.A. Changes in Histophysiology of the Adrenal Medulla in Rats after Prenatal and Postnatal Exposure to Endocrine Disruptor DDT. Bull. Exp. Biol. Med. 2020, 169, 398–400. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, Q.; Lin, L.; Zhang, P.; Li, Z.; Yu, Y.; Ma, F.; Ying, Y.; Li, X.; Ge, R.S. Triadimefon suppresses fetal adrenal gland development after in utero exposure. Toxicology 2021, 462, 152932. [Google Scholar] [CrossRef] [PubMed]
- Niazi, A.K.; Niazi, S.K. Endocrine effects of Fukushima: Radiation-induced endocrinopathy. Indian J. Endocrinol. Metab. 2011, 15, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, S.A.; Rice, S. Endocrine-disrupting chemicals as modulators of sex steroid synthesis. Best Pract. Res. Clin. Endocrinol. Metab. 2006, 20, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Haverinen, E.; Fernandez, M.F.; Mustieles, V.; Tolonen, H. Metabolic Syndrome and Endocrine Disrupting Chemicals: An Overview of Exposure and Health Effects. Int. J. Environ. Res. Public Health 2021, 18, 13047. [Google Scholar] [CrossRef]
- Quesada, I.; Fuentes, E.; Viso-Leon, M.C.; Soria, B.; Ripoll, C.; Nadal, A. Low doses of the endocrine disruptor bisphenol-A and the native hormone 17-beta estradiol rapidly activate transcription factor CREB1. FASEB J. 2002, 16, 1671–1673. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Morimoto, S.; Ripoll, C.; Fuentes, E.; Nadal, A. The estrogenic effect of bisphenol-A disrupts β-cell function in vivo and induces insulin-resistance. Environ. Health Perspect. 2006, 114, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Magdalena, P.; Laribi, O.; Ropero, A.B.; Fuentes, E.; Ripoll, C.; Soria, B.; Nadal, A. Low doses of bisphenol-A and diethystilbestrol impair Ca2+ signals in pancreatic α-cells through a non classical membrane estrogen receptor within intact islets of Langerhans. Environ. Health Perspect. 2005, 113, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Vasu, S.; McClenaghan, N.H.; Flatt, P.R. Molecular Mechanisms of Toxicity and Cell Damage by Chemicals in a Human Pancreatic Beta Cell Line, 1.1B4. Pancreas 2016, 45, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Frederiksen, H.; Nielsen, O.; Koch, H.M.; Skakkebaek, N.E.; Juul, A.; Jørgensen, N.; Andersson, A.M. Changes in urinary excretion of phthalates, phthalate substitutes, bisphenols and other polychlorinated and phenolic substances in young Danish men; 2009–2017. Int. J. Hyg. Environ. Health 2020, 223, 93–105. [Google Scholar] [CrossRef]
- Gyllenhammar, I.; Glynn, A.; Jönsson, B.A.G.; Lindh, C.H.; Darnerud, P.O.; Svensson, K.; Lignell, S. Diverging temporal trends of human exposure to bisphenols and plastizisers, such as phthalates, caused by substitution of legacy EDCs? Environ. Res. 2017, 153, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Lenzen, S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia 2008, 51, 216–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Jin, R.; Lang, J.; Yang, L.A.; Zhao, D.; Ma, Y. PM2.5 promotes β cell damage by increasing inflammatory factors in mice with streptozotocin. Exp. Ther. Med. 2021, 22, 832. [Google Scholar] [CrossRef] [PubMed]
- Roep, B.O.; Thomaidou, S.; van Tienhoven, R.; Zaldumbide, A. Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat. Rev. Endocrinol. 2021, 17, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Stancill, J.S.; Kasmani, M.Y.; Khatun, A.; Cui, W.; Corbett, J.A. Cytokine and Nitric Oxide-Dependent Gene Regulation in Islet Endocrine and Nonendocrine Cells. Function 2021, 3, zqab063. [Google Scholar] [CrossRef]
- Cargnelutti, F.; Di Nisio, A.; Pallotti, F.; Sabovic, I.; Spaziani, M.; Tarsitano, M.G.; Paoli, D.; Foresta, C. Effects of endocrine disruptors on fetal testis development, male puberty, and transition age. Endocrine 2021, 72, 358–374. [Google Scholar] [CrossRef]
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef]
- Lymperi, S.; Giwercman, A. Endocrine disruptors and testicular function. Metabolism 2018, 86, 79–90. [Google Scholar] [CrossRef]
- Ilacqua, A.; Izzo, G.; Emerenziani, G.P.; Baldari, C.; Aversa, A. Lifestyle and fertility: The influence of stress and quality of life on male fertility. Reprod. Biol. Endocrinol. 2018, 16, 115. [Google Scholar] [CrossRef]
- Barbagallo, F.; Condorelli, R.A.; Mongioì, L.M.; Cannarella, R.; Aversa, A.; Calogero, A.E.; La Vignera, S. Effects of Bisphenols on Testicular Steroidogenesis. Front. Endocrinol. 2020, 11, 373. [Google Scholar] [CrossRef]
- Toyama, Y.; Yuasa, S. Effects of neonatal administration of 17beta-estradiol, beta-estradiol 3-benzoate, or bisphenol A on mouse and rat spermatogenesis. Reprod. Toxicol. 2004, 19, 181–188. [Google Scholar] [CrossRef]
- Meeker, J.D.; Ferguson, K.K. Urinary phthalate metabolites are associated with decreased serum testosterone in men, women, and children from NHANES 2011-2012. J. Clin. Endocrinol. Metab. 2014, 99, 4346–4352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.Y.; Wang, Y.; Xie, F.Q.; Li, Y.X.; Wan, X.L.; Ma, W.W.; Wang, D.C.; Wu, Y.H. Analysis of PAEs in semen of infertile men. Int. J. Occup. Environ. Health 2015, 21, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pant, N.; Pant, A.; Shukla, M.; Mathur, N.; Gupta, Y.; Saxena, D. Environmental and experimental exposure of phthalate esters: The toxicological consequence on human sperm. Hum. Exp. Toxicol. 2011, 30, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Gould, J.C.; Leonard, L.S.; Maness, S.C.; Wagner, B.L.; Conner, K.; Zacharewski, T.; Safe, S.; McDonnell, D.P.; Gaido, K.W. Bisphenol A interacts with the estrogen receptor alpha in a distinct manner from estradiol. Mol. Cell. Endocrinol. 1998, 142, 203–214. [Google Scholar] [CrossRef]
- Richter, C.A.; Birnbaum, L.S.; Farabollini, F.; Newbold, R.R.; Rubin, B.S.; Talsness, C.E.; Vandenbergh, J.G.; Walser-Kuntz, D.R.; vom Saal, F.S. In vivo effects of bisphenol A in laboratory rodent studies. Reprod. Toxicol. 2007, 24, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Salian, S.; Doshi, T.; Vanage, G. Perinatal exposure of rats to Bisphenol A affects the fertility of male offspring. Life Sci. 2009, 85, 742–752. [Google Scholar] [CrossRef]
- De Angelis, C.; Mazzella, M.; Pivonello, R. Interferenti endocrini e funzione gonadica: Focus su steroidogenesi testicolare e infertilità maschile. L’Endocrinologo 2022, 23, 52–60. [Google Scholar] [CrossRef]
- Fittipaldi, S.; Bimonte, V.M.; Soricelli, A.; Aversa, A.; Lenzi, A.; Greco, E.A.; Migliaccio, S. Cadmium exposure alters steroid receptors and proinflammatory cytokine levels in endothelial cells in vitro: A potential mechanism of endocrine disruptor atherogenic effect. J. Endocrinol. Investig. 2019, 42, 727–739. [Google Scholar] [CrossRef]
- De Angelis, C.; Galdiero, M.; Pivonello, C.; Salzano, C.; Gianfrilli, D.; Piscitelli, P.; Lenzi, A.; Colao, A.; Pivonello, R. The environment and male reproduction: The effect of cadmium exposure on reproductive function and its implication in fertility. Reprod. Toxicol. 2017, 73, 105–127. [Google Scholar] [CrossRef]
- Bonde, J.P.; Flachs, E.M.; Rimborg, S.; Glazer, C.H.; Giwercman, A.; Ramlau-Hansen, C.H.; Hougaard, K.S.; Høyer, B.B.; Hærvig, K.K.; Petersen, S.B.; et al. The epidemiologic evidence linking prenatal and postnatal exposure to endocrine disrupting chemicals with male reproductive disorders: A systematic review and meta-analysis. Hum. Reprod. Update 2016, 23, 104–125. [Google Scholar] [CrossRef] [Green Version]
- Toppari, J. Environmental endocrine disrupters. Sex Dev. 2008, 2, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, K.K.; Peterson, K.E.; Lee, J.M.; Mercado-García, A.; Blank-Goldenberg, C.; Téllez-Rojo, M.M.; Meeker, J.D. Prenatal and peripubertal phthalates and bisphenol A in relation to sex hormones and puberty in boys. Reprod. Toxicol. 2014, 47, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggert, A.; Cisneros-Montalvo, S.; Anandan, S.; Musilli, S.; Stukenborg, J.B.; Adamsson, A.; Nurmio, M.; Toppari, J. The effects of perfluorooctanoic acid (PFOA) on fetal and adult rat testis. Reprod. Toxicol. 2019, 90, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Mruk, D.; Lian, Q.; Ge, R.; Li, C.; Silvestrini, B.; Cheng, C.Y. Mechanistic Insights into PFOS-Mediated Sertoli Cell Injury. Trends Mol. Med. 2018, 24, 781–793. [Google Scholar] [CrossRef]
- Watkins, D.J.; Sánchez, B.N.; Téllez-Rojo, M.M.; Lee, J.M.; Mercado-García, A.; Blank-Goldenberg, C.; Peterson, K.E.; Meeker, J.D. Impact of phthalate and BPA exposure during in utero windows of susceptibility on reproductive hormones and sexual maturation in peripubertal males. Environ. Health 2017, 16, 69. [Google Scholar] [CrossRef] [Green Version]
- Sergeyev, O.; Burns, J.S.; Williams, P.L.; Korrick, S.A.; Lee, M.M.; Revich, B.; Hauser, R. The association of peripubertal serum concentrations of organochlorine chemicals and blood lead with growth and pubertal development in a longitudinal cohort of boys: A review of published results from the Russian Children’s Study. Rev. Environ. Health 2017, 32, 83–92. [Google Scholar] [CrossRef]
- Den Hond, E.; Roels, H.A.; Hoppenbrouwers, K.; Nawrot, T.; Thijs, L.; Vandermeulen, C.; Winneke, G.; Vanderschueren, D.; Staessen, J.A. Sexual maturation in relation to polychlorinated aromatic hydrocarbons: Sharpe and Skakkebaek’s hypothesis revisited. Environ. Health Perspect. 2002, 110, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, C.L.; Christiansen, S.; Vinggaard, A.M.; Axelstad, M.; Hass, U.; Svingen, T. Anogenital distance as a toxicological or clinical marker for fetal androgen action and risk for reproductive disorders. Arch. Toxicol. 2019, 93, 253–272. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, M.L.; Hsieh, M.H.; Walters, R.C.; Krasnow, R.; Lipshultz, L.I. The relationship between anogenital distance, fatherhood, and fertility in adult men. PLoS ONE 2011, 6, e18973. [Google Scholar] [CrossRef] [Green Version]
- Swan, S.H.; Main, K.M.; Liu, F.; Stewart, S.L.; Kruse, R.L.; Calafat, A.M.; Mao, C.S.; Redmon, J.B.; Ternand, C.L.; Sullivan, S.; et al. Decrease in anogenital distance among male infants with prenatal phthalate exposure. Environ. Health Perspect. 2005, 113, 1056–1061. [Google Scholar] [CrossRef] [Green Version]
- Swan, S.H.; Sathyanarayana, S.; Barrett, E.S.; Janssen, S.; Liu, F.; Nguyen, R.H.; Redmon, J.B.; TIDES Study Team. First trimester phthalate exposure and anogenital distance in newborns. Hum. Reprod. 2015, 30, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, A.; Sabovic, I.; Valente, U.; Tescari, S.; Rocca, M.S.; Guidolin, D.; Dall’Acqua, S.; Acquasaliente, L.; Pozzi, N.; Plebani, M.; et al. Endocrine Disruption of Androgenic Activity by Perfluoroalkyl Substances: Clinical and Experimental Evidence. J. Clin. Endocrinol. Metab. 2019, 104, 1259–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.L.; Lambert, G.H.; Hsu, C.C.; Hsu, M.M. Yucheng: Health effects of prenatal exposure to polychlorinated biphenyls and dibenzofurans. Int. Arch. Occup. Environ. Health 2004, 77, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Leijs, M.M.; Koppe, J.G.; Olie, K.; van Aalderen, W.M.; Voogt, P.; Vulsma, T.; Westra, M.; ten Tusscher, G.W. Delayed initiation of breast development in girls with higher prenatal dioxin exposure; a longitudinal cohort study. Chemosphere 2008, 73, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Richiardi, L.; Bellocco, R.; Adami, H.O.; Torrång, A.; Barlow, L.; Hakulinen, T.; Rahu, M.; Stengrevics, A.; Storm, H.; Tretli, S.; et al. Testicular cancer incidence in eight northern European countries: Secular and recent trends. Cancer Epidemiol. Biomark. Prev. 2004, 13, 2157–2166. [Google Scholar]
- Choi, H.; Kim, J.; Im, Y.; Lee, S.; Kim, Y. The association between some endocrine disruptors and hypospadias in biological samples. J. Environ. Sci. Health Part A 2012, 47, 2173–2179. [Google Scholar] [CrossRef]
- Carbone, P.; Giordano, F.; Nori, F.; Mantovani, A.; Taruscio, D.; Lauria, L.; Figà-Talamanca, I. The possible role of endocrine disrupting chemicals in the aetiology of cryptorchidism and hypospadias: A population-based case-control study in rural Sicily. Int. J. Androl. 2007, 30, 3–13. [Google Scholar] [CrossRef]
- Main, K.M.; Mortensen, G.K.; Kaleva, M.M.; Boisen, K.A.; Damgaard, I.N.; Chellakooty, M.; Schmidt, I.M.; Suomi, A.M.; Virtanen, H.E.; Petersen, D.V.H.; et al. Human breast milk contamination with phthalates and alterations of endogenous reproductive hormones in infants three months of age. Environ. Health Perspect. 2006, 114, 270–276. [Google Scholar] [CrossRef]
- Brucker-Davis, F.; Wagner-Mahler, K.; Delattre, I.; Ducot, B.; Ferrari, P.; Bongain, A.; Kurzenne, J.Y.; Mas, J.C.; Fénichel, P.; Cryptorchidism Study Group from Nice Area. Cryptorchidism at birth in Nice area (France) is associated with higher prenatal exposure to PCBs and DDE, as assessed by colostrum concentrations. Hum. Reprod. 2008, 23, 1708–1718. [Google Scholar] [CrossRef] [Green Version]
- Vizza, D.; Perri, A.; Toteda, G.; Lupinacci, S.; Leone, F.; Gigliotti, P.; Lofaro, D.; La Russa, A.; Bonofiglio, R. Nerve growth factor exposure promotes tubular epithelial-mesenchymal transition via TGF-β1 signaling activation. Growth Factors 2015, 33, 169–180. [Google Scholar] [CrossRef]
- Cannarella, R.; Condorelli, R.A.; Gusmano, C.; Barone, N.; Burrello, N.; Aversa, A.; Calogero, A.E.; La Vignera, S. Temporal Trend of Conventional Sperm Parameters in a Sicilian Population in the Decade 2011–2020. J. Clin. Med. 2021, 10, 993. [Google Scholar] [CrossRef] [PubMed]
- Guarnotta, V.; Amodei, R.; Giordano, C. Metabolic comorbidities of adrenal insufficiency: Focus on steroid replacement therapy and chronopharmacology. Curr. Opin. Pharmacol. 2021, 60, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Cecconi, S.; Gualtieri, G.; Di Bartolomeo, A.; Troiani, G.; Cifone, M.G.; Canipari, R. Evaluation of the effects of extremely low frequency electromagnetic fields on mammalian follicle development. Hum. Reprod. 2000, 15, 2319–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, S.S.; Khaki, A.A.; Ainehchi, N.; Alihemmati, A.; Khatooni, A.A.; Khaki, A.; Asghari, A. Effect of non-ionizing electromagnetic field on the alteration of ovarian follicles in rats. Electron. Physician. 2016, 8, 2168–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bath, L.E.; Wallace, W.H.; Critchley, H.O. Late effects of the treatment of childhood cancer on the female reproductive system and the potential for fertility preservation. BJOG Int. J. Obstet. Gynaecol. 2002, 109, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Divyashree, S.; Yajurvedi, H.N. Long-term chronic stress exposure induces PCO phenotype in rat. Reproduction 2016, 152, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Chrousos, G.P.; Torpy, D.J.; Gold, P.W. Interactions of the hypothalamo pituitary-adrenal axis and the female reproductive system: Clinical implications. Ann. Intern. Med. 1998, 129, 229–240. [Google Scholar] [CrossRef]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoid, stress, and fertility. Minerva Endocrinol. 2010, 35, 109–125. [Google Scholar]
- Wu, L.M.; Liu, Y.S.; Tong, X.H.; Shen, N.; Jin, R.T.; Han, H.; Hu, M.H.; Wang, W.; Zhou, G.X. Inhibition of follicular development induced by chronic unpredictable stress is associated with growth and differentiation factor 9 and gonadotropin in mice. Biol. Reprod. 2011, 86, 1–7. [Google Scholar] [CrossRef]
- Saraswathi, C.D.; Sreemantula, S.; Prakash, W.S. Effect of chronic cold restraint and immobilization stress on estrous cycle in rats. Pharmacol. Online 2010, 2, 151–160. [Google Scholar]
- Spiers, J.G.; Chen, H.C.; Sernia, C.; Lavidis, N.A. Activation of the hypothalamic-pituitary-adrenal stress axis induces cellular oxidative stress. Front. Neurosci. 2015, 8, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.Y.; Chen, H.H.; Zhang, N.; Ding, M.X.; Qiu, Y.E.; Pan, X.M.; Fang, Y.S.; Lin, Y.P.; Zheng, Q.; Wang, W.Q. Effects of chronic unpredictable mild stress on ovarian reserve in female rats: Feasibility analysis of a rat model of premature ovarian failure. Mol. Med. Rep. 2018, 18, 532–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Wu, X.Y.; Zhu, Q.B.; Li, J.; Shi, L.G.; Wu, J.L.; Zhang, Q.J.; Huang, M.L.; Bao, A.M. Sex differences in the stress response in SD rats. Behav. Brain Res. 2015, 284, 231–237. [Google Scholar] [CrossRef]
- Gao, Z.; Gao, X.; Fan, W.; Liu, S.; Li, M.; Miao, Y.; Ding, C.; Tang, Z.; Yan, L.; Liu, G.; et al. Bisphenol A and genistein have opposite effects on adult chicken ovary by acting on ERα/Nrf2-Keap1-signaling pathway. Chem. Biol. Interact. 2021, 347, 109616. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hafner, K.S.; Flaws, J.A. In utero bisphenol A exposure disrupts germ cell nest breakdown and reduces fertility with age in the mouse. Toxicol. Appl. Pharmacol. 2014, 276, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souter, I.; Smith, K.W.; Dimitriadis, I.; Ehrlich, S.; Williams, P.L.; Calafat, A.M.; Hauser, R. The association of bisphenol-A urinary concentrations with antral follicle counts and other measures of ovarian reserve in women undergoing infertility treatments. Reprod. Toxicol. 2013, 42, 224–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, P.A.; Koehler, K.E.; Susiarjo, M.; Hodges, C.A.; Ilagan, A.; Voigt, R.C.; Thomas, S.; Thomas, B.F.; Hassold, T.J. Bisphenol a exposure causes meiotic aneuploidy in the female mouse. Curr. Biol. 2003, 13, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Téteau, O.; Jaubert, M.; Desmarchais, A.; Papilier, P.; Binet, A.; Maillard, V.; Elis, S. Bisphenol A and S impaired ovine granulosa cell steroidogenesis. Reproduction 2020, 159, 571–583. [Google Scholar] [CrossRef]
- Song, W.; Lu, H.; Wu, K.; Zhang, Z.; Shuk-Wa Lau, E.; Ge, W. Genetic evidence for estrogenicity of bisphenol A in zebrafish gonadal differentiation and its signaling mechanism. J. Hazard. Mater. 2020, 386, 121886. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, J.; Liao, L.; Han, S.; Liu, J. Effect of bisphenol A on steroid hormone production in rat ovarian theca-interstitial and granulosa cells. Mol. Cell. Endocrinol. 2008, 283, 12–18. [Google Scholar] [CrossRef]
- Ziv-Gal, A.; Craig, Z.R.; Wang, W.; Flaws, J.A. Bisphenol A inhibits cultured mouse ovarian follicle growth partially via the aryl hydrocarbon receptor signaling pathway. Reprod. Toxicol. 2013, 42, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Liu, C.; Chen, M.; Yan, J.; Wang, C.; Zuo, Z.; He, C. The interference effects of bisphenol A on the synthesis of steroid hormones in human ovarian granulosa cells. Environ. Toxicol. 2021, 36, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, P.; D’Auria, R.; Viggiano, A.; Ciaglia, E.; Meccariello, R.; Dello Russo, R.; Puca, A.A.; Vecchione, C.; Nori, S.L.; Santoro, A. Bisphenol A induces DNA damage in cells exerting immune surveillance functions at peripheral and central level. Chemosphere 2020, 254, 126819. [Google Scholar] [CrossRef] [PubMed]
- Hannon, P.R.; Brannick, K.E.; Wang, W.; Flaws, J.A. Mono(2-ethylhexyl) phthalate accelerates early folliculogenesis and inhibits steroidogenesis in cultured mouse whole ovaries and antral follicles. Biol. Reprod. 2015, 92, 120. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Chen, J.A.; Qiu, Z.; Zhao, Q.; Luo, J.; Yang, L.; Zeng, H.; Huang, Y.; Zhang, L.; Cao, J.; et al. Ovotoxicity and PPAR-mediated aromatase downregulation in female Sprague-Dawley rats following combined oral exposure to benzo[a]pyrene and di-(2-ethylhexyl) phthalate. Toxicol. Lett. 2010, 199, 323–332. [Google Scholar] [CrossRef]
- Uzumcu, M.; Kuhn, P.E.; Marano, J.E.; Armenti, A.E.; Passantino, L. Early postnatal methoxychlor exposure inhibits folliculogenesis and stimulates anti-Mullerian hormone production in the rat ovary. J. Endocrinol. 2006, 191, 549–558. [Google Scholar] [CrossRef]
- Yu, K.; Zhang, X.; Tan, X.; Ji, M.; Chen, Y.; Wan, Z.; Yu, Z. Multigenerational and transgenerational effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin exposure on ovarian reserve and follicular development through AMH/AMHR2 pathway in adult female rats. Food Chem. Toxicol. 2020, 140, 111309. [Google Scholar] [CrossRef]
- Jablonska, O.; Piasecka, J.; Petroff, B.K.; Nynca, A.; Siawrys, G.; Wąsowska, B.; Zmijewska, A.; Lewczuk, B.; Ciereszko, R.E. In vitro effects of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) on ovarian, pituitary, and pineal function in pigs. Theriogenology 2011, 76, 921–932. [Google Scholar] [CrossRef]
- Liu, T.; Di, Q.N.; Sun, J.H.; Zhao, M.; Xu, Q.; Shen, Y. Effects of nonylphenol induced oxidative stress on apoptosis and autophagy in rat ovarian granulosa cells. Chemosphere 2020, 261, 127693. [Google Scholar] [CrossRef]
- Yu, P.L.; Lin, H.W.; Wang, S.W.; Wang, P.S. Effects of nonylphenol on the production of progesterone on the rats granulosa cells. J. Cell. Biochem. 2011, 112, 2627–2636. [Google Scholar] [CrossRef]
- Chang, C.H.; Tsai, M.S.; Lin, C.L.; Hou, J.W.; Wang, T.H.; Tsai, Y.A.; Liao, K.W.; Mao, I.F.; Chen, M.L. The association between nonylphenols and sexual hormones levels among pregnant women: A cohort study in Taiwan. PLoS ONE 2014, 9, e104245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.S.; Chang, C.H.; Tsai, Y.A.; Liao, K.W.; Mao, I.F.; Wang, T.H.; Hwang, S.M.; Chang, Y.J.; Chen, M.L. Neonatal outcomes of intrauterine nonylphenol exposure--a longitudinal cohort study in Taiwan. Sci. Total Environ. 2013, 458, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Kolesarova, A.; Roychoudhury, S.; Slivkova, J.; Sirotkin, A.; Capcarova, M.; Massanyi, P. In vitro study on the effects of lead and mercury on porcine ovarian granulosa cells. J. Environ. Sci. Health Part A 2010, 45, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Niu, H.; Wang, J.; Wang, Q.; Li, Y. Potential mechanism of lead poisoning to the growth and development of ovarian follicle. Toxicology 2021, 457, 152810. [Google Scholar] [CrossRef]
- da Costa, C.S.; Oliveira, T.F.; Freitas-Lima, L.C.; Padilha, A.S.; Krause, M.; Carneiro, M.T.W.D.; Salgado, B.S.; Graceli, J.B. Subacute cadmium exposure disrupts the hypothalamic-pituitary-gonadal axis, leading to polycystic ovarian syndrome and premature ovarian failure features in female rats. Environ. Pollut. 2021, 269, 116154. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, T.; Zhang, C.; Luo, L.; Xie, M.; Huang, H. Cadmium exposure in newborn rats ovary induces developmental disorders of primordial follicles and the differential expression of SCF/c-kit gene. Toxicol. Lett. 2017, 280, 20–28. [Google Scholar] [CrossRef]
- Merlo, E.; Schereider, I.R.G.; Simões, M.R.; Vassallo, D.V.; Graceli, J.B. Mercury leads to features of polycystic ovary syndrome in rats. Toxicol. Lett. 2019, 312, 45–54. [Google Scholar] [CrossRef]
- Mínguez-Alarcón, L.; Williams, P.L.; Souter, I.; Sacha, C.; Amarasiriwardena, C.J.; Ford, J.B.; Hauser, R.; Chavarro, J.E.; Earth Study Team. Hair mercury levels, intake of omega-3 fatty acids and ovarian reserve among women attending a fertility center. Int. J. Hyg. Environ. Health 2021, 237, 113825. [Google Scholar] [CrossRef]
- Altunkaynak, B.Z.; Akgül, N.; Yahyazadeh, A.; Altunkaynak, M.E.; Turkmen, A.P.; Akgül, H.M.; Ünal, B. Effect of mercury vapor inhalation on rat ovary: Stereology and histopathology. J. Obstet. Gynaecol. Res. 2016, 42, 410–416. [Google Scholar] [CrossRef]
- Troisi, R.; Hatch, E.E.; Titus, L.; Strohsnitter, W.; Gail, M.H.; Huo, D.; Adam, E.; Robboy, S.J.; Hyer, M.; Hoover, R.N.; et al. Prenatal diethylstilbestrol exposure and cancer risk in women. Environ. Mol. Mutagen 2019, 60, 395–403. [Google Scholar] [CrossRef]
- Clement, R.; Guilbaud, E.; Barrios, L.; Rougé-Maillart, C.; Jousset, N.; Rodat, O. DES daughters in France: Experts’ points of view on the various genital, uterine and obstetric pathologies, and in utero DES exposure. Med. Sci. Law 2014, 54, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Medigović, I.; Ristić, N.; Trifunović, S.; Manojlović-Stojanoski, M.; Milošević, V.; Zikić, D.; Nestorović, N. Genistein affects ovarian folliculogenesis: A stereological study. Microsc. Res. Tech. 2012, 75, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Kaminska, B.; Ciereszko, R.; Kiezun, M.; Dusza, L. In vitro effects of genistein and daidzein on the activity of adrenocortical steroidogenic enzymes in mature female pigs. J. Physiol. Pharmacol. 2013, 64, 103–108. [Google Scholar] [PubMed]
- Tiemann, U.; Schneider, F.; Vanselow, J.; Tomek, W. In vitro exposure of porcine granulosa cells to the phytoestrogens genistein and daidzein: Effects on the biosynthesis of reproductive steroid hormones. Reprod. Toxicol. 2007, 24, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Toktay, E.; Selli, J.; Gurbuz, M.A.; Tastan, T.B.; Ugan, R.A.; Un, H.; Halici, Z. Effects of soy isoflavonoids (genistein and daidzein) on endometrial receptivity. Iran. J. Basic Med. Sci. 2020, 23, 1603–1609. [Google Scholar]
- González-Pleiter, M.; Gonzalo, S.; Rodea-Palomares, I.; Leganés, F.; Rosal, R.; Boltes, K.; Marco, E.; Fernández-Piñas, F. Toxicity of five antibiotics and their mixtures towards photosynthetic aquatic organisms: Implications for environmental risk assessment. Water Res. 2013, 47, 2050–2064. [Google Scholar] [CrossRef]
- Fang, S.; Wang, D.; Zhang, X.; Long, X.; Qin, M.M.; Lin, Z.; Liu, Y. Similarities and differences in combined toxicity of sulfonamides and other antibiotics towards bacteria for environmental risk assessment. Environ. Monit. Assess. 2016, 188, 429. [Google Scholar] [CrossRef]
- Rider, C.V.; Furr, J.; Wilson, V.S.; Gray, L.E., Jr. A mixture of seven antiandrogens induces reproductive malformations in rats. Int. J. Androl. 2008, 31, 249–262. [Google Scholar] [CrossRef]
- Meling, D.D.; Warner, G.R.; Szumski, J.R.; Gao, L.; Gonsioroski, A.V.; Rattan, S.; Flaws, J.A. The effects of a phthalate metabolite mixture on antral follicle growth and sex steroid synthesis in mice. Toxicol. Appl. Pharmacol. 2020, 388, 114875. [Google Scholar] [CrossRef]
- Zhou, C.; Gao, L.; Flaws, J.A. Prenatal exposure to an environmentally relevant phthalate mixture disrupts reproduction in F1 female mice. Toxicol. Appl. Pharmacol. 2017, 318, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Hamid, N.; Deng, S.; Jia, P.P.; Pei, D.S. Individual and combined toxicogenetic effects of microplastics and heavy metals (Cd, Pb, and Zn Oryzias melastigma). J. Hazard. Mater. 2020, 397, 122795. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, W.; Liu, J.; Sun, Y.; Li, Y.; Li, H.; Xiao, S.; Shen, X. Metabolomic changes in follicular fluid induced by soy isoflavones administered to rats from weaning until sexual maturity. Toxicol. Appl. Pharmacol. 2013, 269, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, S.; Ploessl, F.; Bracher, F.; Laforsch, C. Single and combined toxicity of pharmaceuticals at environmentally relevant concentrations in Daphnia magna—A multigenerational study. Chemosphere 2010, 79, 60–66. [Google Scholar] [CrossRef]
- Mathieu-Denoncourt, J.; Wallace, S.J.; de Solla, S.R.; Langlois, V.S. Plasticizer endocrine disruption: Highlighting developmental and reproductive effects in mammals and non-mammalian aquatic species. Gen. Comp. Endocrinol. 2015, 219, 74–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikkam, M.; Tracey, R.; Guerrero-Bosagna, C.; Skinner, M.K. Plastics derived endocrine disruptors (BPA, DEHP and DBP) induce epigenetic transgenerational inheritance of obesity, reproductive disease and sperm epimutations. PLoS ONE 2013, 8, e55387. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.; Larsen, G.; Manikkam, M.; Guerrero-Bosagna, C.; Savenkova, M.I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of ovarian disease. PLoS ONE 2012, 7, e36129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, T.; Hiromi, J. Toxicity of 17alpha-ethynylestradiol and norethindrone, constituents of an oral contraceptive pill to the swimming and reproduction of cladoceran Daphnia magna, with special reference to their synergetic effect. Mar. Pollut. Bull. 2003, 47, 139–142. [Google Scholar] [CrossRef]
- Schmitz, M.; Beghin, M.; Mandiki, S.N.M.; Nott, K.; Gillet, M.; Ronkart, S.; Robert, C.; Baekelandt, S.; Kestemont, P. Environmentally-relevant mixture of pharmaceutical drugs stimulates sex-steroid hormone production and modulates the expression of candidate genes in the ovary of juvenile female rainbow trout. Aquat. Toxicol. 2018, 205, 89–99. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guarnotta, V.; Amodei, R.; Frasca, F.; Aversa, A.; Giordano, C. Impact of Chemical Endocrine Disruptors and Hormone Modulators on the Endocrine System. Int. J. Mol. Sci. 2022, 23, 5710. https://doi.org/10.3390/ijms23105710
Guarnotta V, Amodei R, Frasca F, Aversa A, Giordano C. Impact of Chemical Endocrine Disruptors and Hormone Modulators on the Endocrine System. International Journal of Molecular Sciences. 2022; 23(10):5710. https://doi.org/10.3390/ijms23105710
Chicago/Turabian StyleGuarnotta, Valentina, Roberta Amodei, Francesco Frasca, Antonio Aversa, and Carla Giordano. 2022. "Impact of Chemical Endocrine Disruptors and Hormone Modulators on the Endocrine System" International Journal of Molecular Sciences 23, no. 10: 5710. https://doi.org/10.3390/ijms23105710
APA StyleGuarnotta, V., Amodei, R., Frasca, F., Aversa, A., & Giordano, C. (2022). Impact of Chemical Endocrine Disruptors and Hormone Modulators on the Endocrine System. International Journal of Molecular Sciences, 23(10), 5710. https://doi.org/10.3390/ijms23105710