
Current Perspectives on Synthetic Compartments for Biomedical Applications

 , , ,
, , ,
Abstract
:
1. Introduction
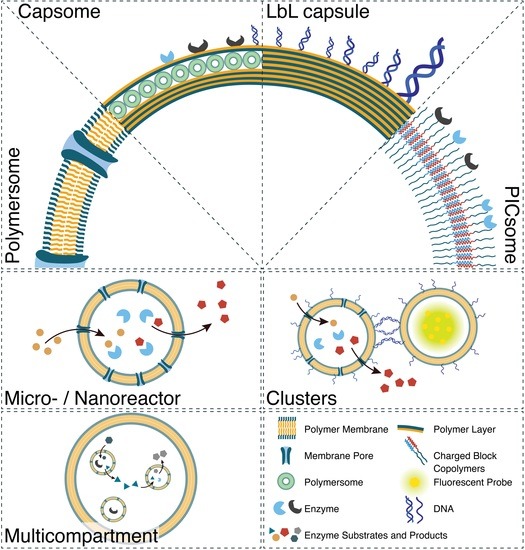
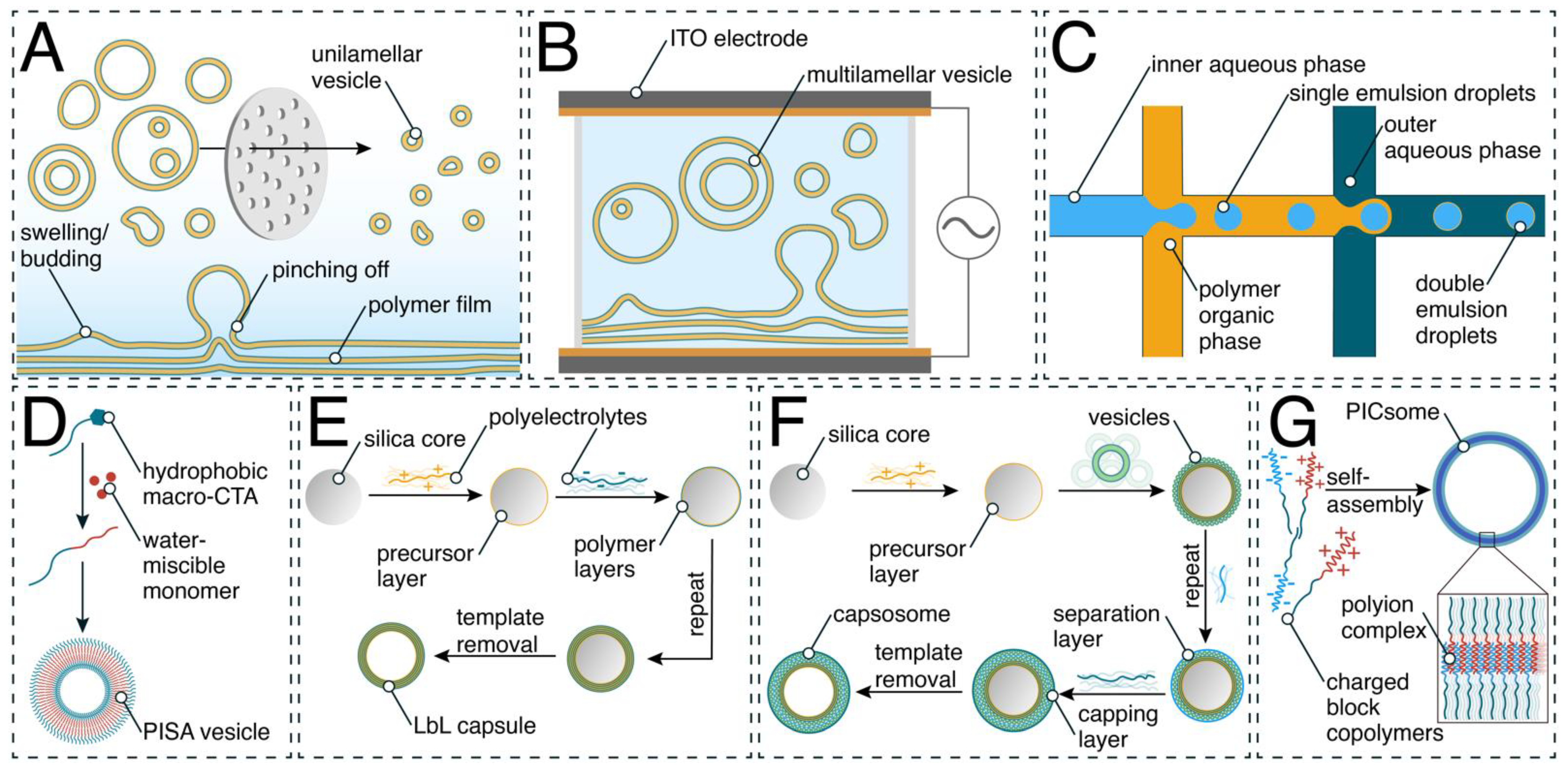
2. Generation of Synthetic Compartments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
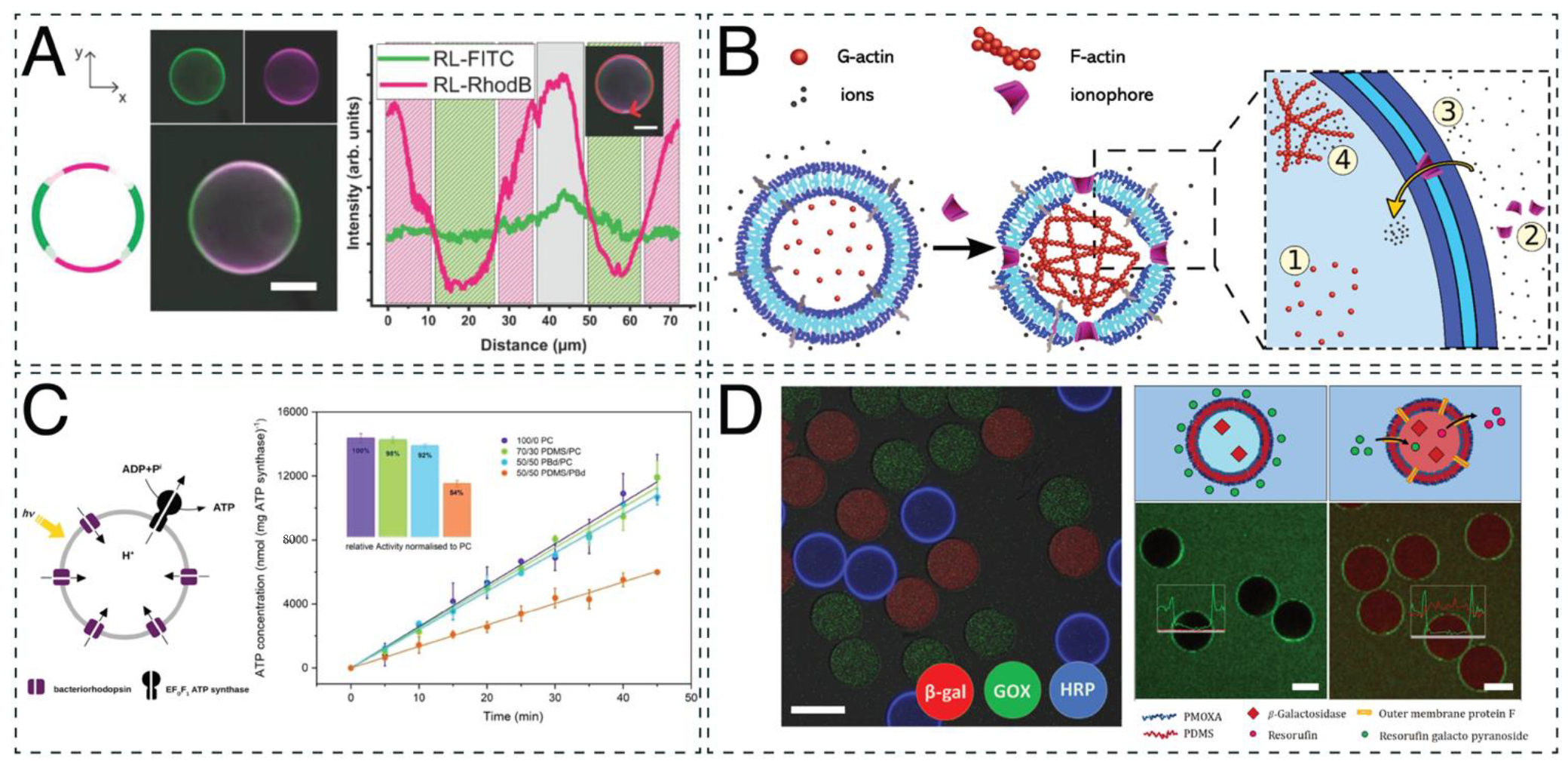{kind=link}
{kind=link}
| Polymer | Method of Self-Assembly | Characteristics |
|---|---|---|
| Carbohydrate-b-PPG | Direct hydration method [54] | Forms capsosomes, inherently permeable to low-molecular-weight compounds |
| Chitosan | Sonication-assisted mixing (capsules) [55], LbL [56] | Biocompatible, natural polymer |
| CTAB | LbL [56] | Surfactant, forms micelles in the absence of another polymer |
| PA/DEX | LbL [57] | Biocompatible polysaccharide (anionic) |
| P(OEGMA300-grad-HPMA) | PISA [43] | Biocompatible assembly, monomers and a macromolecular precursor need to be: (i) solvophilic and (ii) compatible with each other |
| PA/PLA | LbL [57] | Biocompatible cationic polyelectrolyte |
| PAA | LbL [58] | Anionic polyelectrolyte |
| PAH | LbL [59,60] | Cationic polyelectrolyte |
| PAMAM | Mixing (PICsomes) [61] | Dendrimer (branched structure) |
| P(Asp-AP) | Mixing (PICsomes) [62,63,64] | Anionic polyelectrolyte, forms PICsomes, cannot form vesicles on its own |
| PATK | Mixing (PICsomes) [65] | Cationic polyelectrolyte |
| PBd-b-PEG | Double emulsion microfluidics [37] | Biocompatible |
| PBd–b-PEO | Emulsion centrifugation [34], Electroformation [66], Film rehydration [67] | Pure or as hybrid (with POPC) polymersomes for membrane protein insertion, assembly of asymmetric polymer/lipid (POPC) hybrid membranes |
| PBO-b-PG | Microfluidic double emulsion, solvent switch [68] | Biocompatible |
| PCL-b-P[Glu-stat-(Glu-ADA)] | Solvent switch [69] | Biodegradable, bone-targeting |
| PCL-b-PTrp-b-P(Lys-statPhe) | Solvent switch [70] | Biocompatible, biodegradable, antibacterial |
| PDMS-b-heparin | Film rehydration [71] | Forms polymersomes in combination with PMOXA-b-PDMS-b-PMOXA, forms micelles by itself |
| PDMS-g-PEO | Electroformation, Film rehydration [72] | Pure or as hybrid (with PC) polymersomes and GUVs for membrane protein insertion |
| PEG-b-PCL | Electroformation [73], film rehydration [74] | Multidomain membrane formation with lipids (DPPC) |
| PEG-P(CLgTMC) | Direct hydration method [75] | Biodegradable, intrinsic fluorescence |
| PEG-b-P(CPTKMA-co-PEMA) | Solvent exchange method [76] | Biocompatible, conjugated with campthothecin |
| PEG-GPLGVRG-PCL-PGPMA | Film hydration method [77] | Biocompatible, MMP-cleavable peptide and CPP-mimicking polymer |
| PEG-b-PHPMA | PISA [78] | Highly hydrated membrane, size-selective transport of molecules |
| PEG-b-PIC | Solvent exchange [79] | Biocompatible, iodine-rich for SPECT/CT and radioisotope therapy |
| PEG-b-PLA | Film rehydration [74], double emulsion microfluidics [37] | Forms polymersomes with and without lipid mixing, biodegradable |
| PEG-b-polypeptide | Mixing (PICsomes) [80] | pH-responsive, biocompatible |
| PEG-b-PAsp | Mixing (PICsomes) [62,64] | Linear polymer, forms PICsomes, micelles or hydrogels, biocompatible |
| PEG-b-PS | Solvent switch method [81,82] | Biocompatible, formation of stomatocytes, rigid assemblies |
| PEI-b-PDLLA | Microfluidic double emulsion [83] | Biocompatible, cationic assemblies, can form polymer stomatocytes |
| PEO-b-PBO | Film rehydration [84] | Forms asymmetric polymersomes |
| PEO-b-PCL | Emulsification-induced assembly [85] | Low interfacial tension solvent or SDS is needed to control the assembly |
| PEO-b-PCL-b-PMOXA | Film rehydration [86] | Rehydration at 62 °C due to the semi crystalline nature of the PCL block |
| PEO-b-P(CMA-stat-DEA-stat-GEMA) | Solvent exchange method [87] | Biocompatible, CMA photocrosslinking stabilization |
| PEO-b-PEHOx-b-PEtOz | Solvent switch, film rehydration [26] | Asymmetric membrane, can be used for directed protein insertion |
| PEO-b-PPO-b-PEO (Pluronics L121) | Double emulsion microfluidics [37] | Assembly via DNA linkage |
| PiB-b-PEG | Freeze–thaw extrusion [88] | Biocompatible, high chemical and thermal stability |
| PLys | Mixing (PICsomes) [89] | Cationic polyelectrolyte |
| PMA | LbL [60] | Labor-intensive LbL assembly |
| PMOXA-b-PDMS | Film rehydration [90,91], microfluidic double emulsion [39] | Formation of nano- and micro-sized vesicles in biocompatible, aqueous conditions, various channels and proteins can be inserted |
| PMOXA-b-PDMS-b-PMOXA | Film rehydration [71,92] | Formation of nano and micro-sized vesicles in biocompatible, aqueous conditions, various channels and proteins can be inserted |
| PMPC-b-PDPA | Film rehydration [84,93] | Formation of (asymmetric) polymersomes, can be electroporated |
| POEGMA-b-P(ST-co-VBA) | PISA [41] | Biocompatible assembly, monomers and a macromolecular precursor need to be: (i) solvophilic and (ii) compatible with each other |
| Poly(dopamine) | LbL [94] | Simplified LbL capsule formation |
| PS-b-PEO | Emulsification [95] | High capacity of ammonia capture in bile salt-containing buffer |
| PSMA-PBzMA | PISA [42] | Biocompatible assembly, monomers and a macromolecular precursor need to be: (i) solvophilic and (ii) compatible with each other |
| PSS-b-PEO-b-PSS | Mixing (PICsomes) [61] | Forms PICsomes with loops within the membrane when combined with poly(amidoamine) dendrimers |
| PVP | LbL [96] | Work-intensive LbL assembly |
2.1. Surface Functionalization of Polymer Compartments
2.2. Assemblies of Compartments
3. Requirements for Compartments to Be Used in Biomedical Applications
4. Applications of Compartments in the Biomedical Field
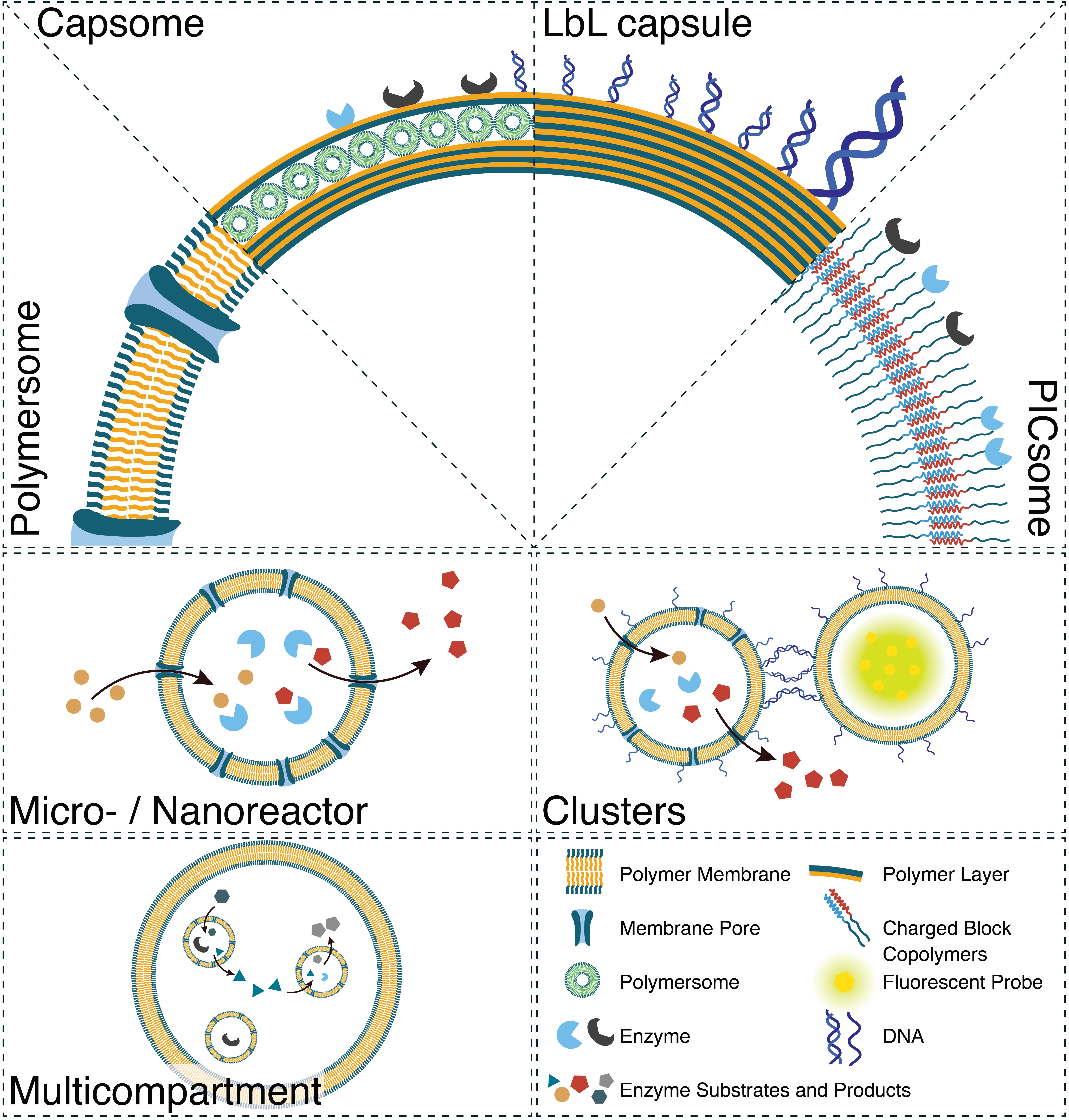
4.1. Imaging and Theranostic Applications
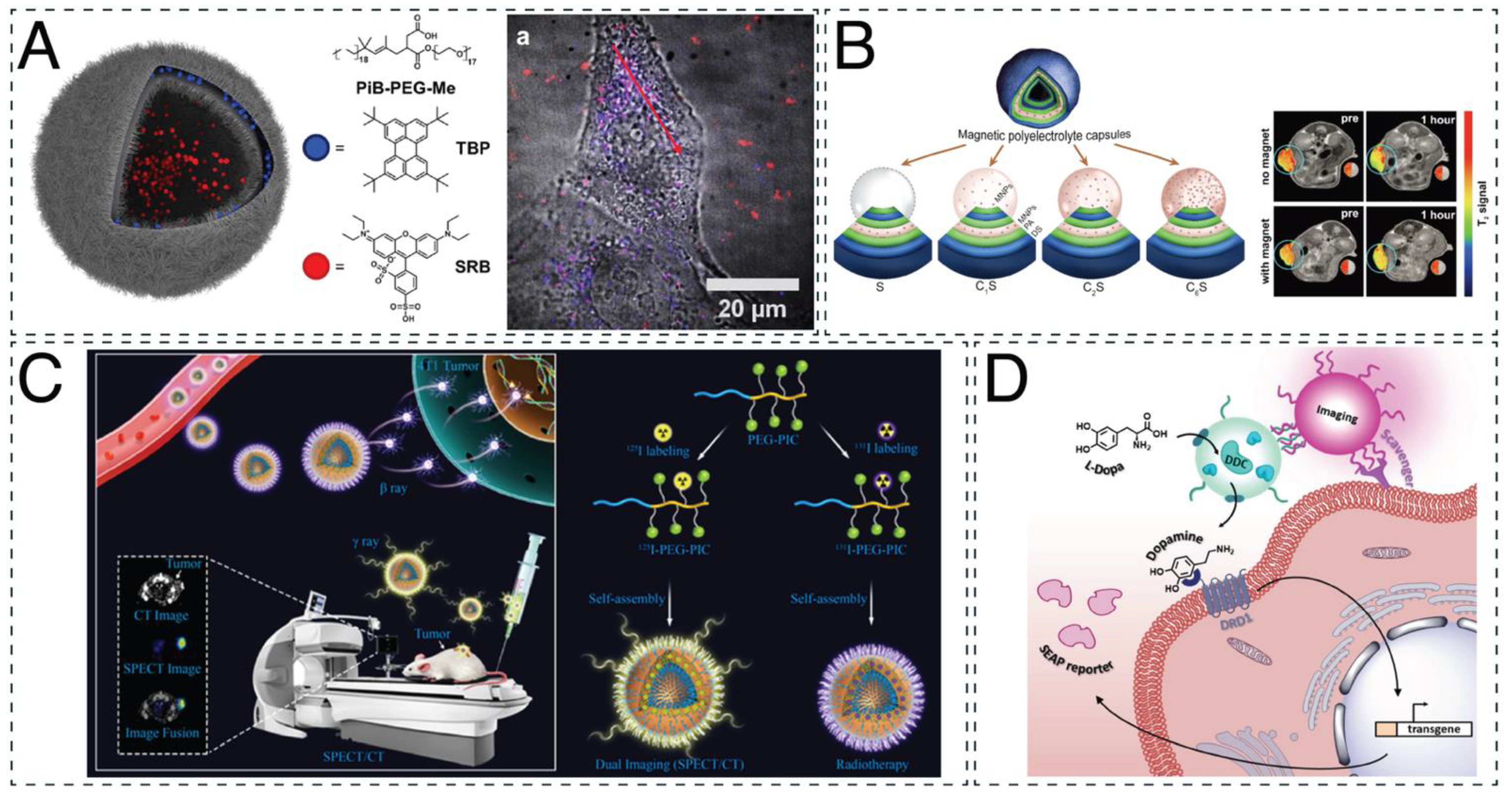
4.2. Therapeutic Applications: From In Vitro to In Vivo
4.2.1. In Vitro Studies
4.2.2. In Vivo Studies
4.3. Vesicles as Model Systems for Organelles and Cells
5. Conclusion and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADA | alendronic acid |
| ADP | adenosine diphosphate |
| AP | poly([5-aminopentyl]-α,β-aspartamide) |
| ATP | adenosine triphosphate |
| BDL | bile duct-ligated |
| BMD | bone mineral density |
| cGMP | cyclic guanosine monophosphate |
| CLSM | confocal laser scanning microscope |
| CMA | 7-(2-methacryloyloxyethoxy)-4-methylcoumarin |
| CNC | catalytic nanocompartment |
| CPP | cell-penetrating peptide |
| CPT | camptothecin |
| CPTKMA | thioketal-linked CPT methacrylate monomer |
| cRGD | cyclic arginine-glycine-aspartic |
| CT | computed tomography |
| CTAB | cetyltrimethylammonium bromide |
| DBCO | dibenzocyclooctyne |
| DDC | dopa decarboxylase |
| DEA | 2-(diethylamino)ethyl methacrylate |
| DEX | dextran sulfate |
| DMPC | 1,2-dimyristoyl-sn-glycero-3-phosphocholine |
| DNA | deoxyribonucleic acid |
| DPPC | 1,2-dipalmitoyl-sn-glycero-3-phosphocholine |
| DTT | dithiothreitol |
| E. coli | Escherichia coli |
| FGD | fluorescein di-β-galactopyranoside |
| FITC | fluorescein isothiocyanate |
| FRAP | fluorescence recovery after photobleaching |
| GEMA | (α-d-glucopyranosyl)ethyl methacrylate |
| Gox | glucose oxidase |
| GUV | giant unilamellar vesicle |
| HPTS | 8-hydroxypyrene-1,3,6-trisulfonate |
| HRP | horseradish peroxidase |
| iNOS | nitric oxide synthase |
| ITO | indium tin oxide |
| LbL | layer-by-layer |
| L-DOPA | levodopa/l-3,4-dihydroxyphenylalanine |
| LPO | lactoperoxidase |
| MMP | matrix metalloproteinase |
| mNSS | modified neurological severity scores |
| mPEG | methoxy-poly(ethylene glycol) |
| MRI | magnetic Resonance Imaging |
| NgR | Nogo-66 receptor |
| NIR | near-infrared |
| NP | nanoparticle |
| OEGMA | oligo(ethylene glycol) methyl ether methacrylate |
| OmpF | outer membrane protein F (from Escherichia coli) |
| PA | poly-L-arginine |
| PAA | poly(acrylic acid) |
| PAH | poly(allylamine hydrochloride) |
| PAMAM | poly(amidoamine) |
| PAsp | poly(α,β-aspartic acid) |
| PATK | poly([2-[[1-[(2-aminoethyl)thio]-1-methylethyl]thio]ethyl]-α,β-aspartamide) |
| PBd | poly(1,2-butadiene) |
| PBO | poly(butylene oxide) |
| PBzMA | poly(benzyl methacrylate) |
| PC | phosphatidylcholine |
| PCL | poly(ε-caprolactone) |
| PDA | polydopamine |
| PDLLA | poly(D,L-Lactic Acid) |
| PDMS | poly(dimethylsiloxane) |
| PDPA | poly [2-(diisopropylamino)ethyl methacrylate] |
| PEG | poly(ethylene glycol) |
| PEHO | poly(3-ethyl-3-hydroxymethyloxetane) |
| PEI | poly(ether imide) |
| PEMA | poly(ethyl methacrylate) |
| PEO | poly(ethylene oxide) |
| PEtOz | poly(2-ethyl-2-oxazoline) |
| PG | poly(glycidol) |
| PGPMA | poly(3-guanidinopropyl methacrylamide) |
| PHPMA | poly(N-(2-Hydroxypropyl) methacrylamide) |
| Pi | inorganic phosphate |
| PiB | polyisobutylene |
| PIC | polyion complex |
| PISA | polymerization-induced self-assembly |
| PL | phospholipids |
| PLA | polycaprolactone |
| PLys | poly-lysine |
| PMA | polymethyl acrylate |
| PMOXA | poly(2-methyl-2-oxazoline) |
| PMPC | poly(2-methacryloyloxyethyl phosphorylcholine) |
| POEGMA | poly(oligo(ethylene glycol) methyl ether methacrylate) |
| POPC | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine |
| PS | polystyrene |
| PSS | poly(styrene sulfonate) |
| PSMA | poly(stearyl methacrylate) |
| PPG | poly(propylene glycol) |
| PPO | poly(p-phenylene oxide) |
| PTMC | tetraphenylethylene pyridinium modified trimethylenecarbonate |
| PVP | polyvinylpyrrolidone |
| P(CLgTMC) | poly(caprolactone-gradient-trimethylene carbonate) |
| ROS | reactive oxygen species |
| S. aureus | Staphylococcus aureus |
| sGC | soluble guanylyl cyclase |
| siRNA | small interfering ribonucleic acid |
| SPAAC | strain-promoted azide-alkyne cycloaddition |
| SPECT | single-photon emission computed tomography |
| SPIONs | superparamagnetic iron oxide nanoparticles |
| ssDNA | single-stranded deoxyribonucleic acid |
| TA | tannic acid |
| TTA-UC | triplet–triplet annihilation based molecular photon upconversion |
| UOX | urate oxidase |
| USIONs | ultrasmall iron oxide nanoparticles |
| VBA | poly(vinyl benzaldehyde) |
References
- Palivan, C.G.; Goers, R.; Najer, A.; Zhang, X.; Car, A.; Meier, W. Bioinspired polymer vesicles and membranes for biological and medical applications. Chem. Soc. Rev. 2016, 45, 377–411. [Google Scholar] [CrossRef] [Green Version]
- Matoori, S.; Leroux, J.C. Twenty-five years of polymersomes: Lost in translation? Mater. Horiz. 2020, 7, 1297–1309. [Google Scholar] [CrossRef]
- Feng, H.; Lu, X.; Wang, W.; Kang, N.-G.; Mays, J.W. Block copolymers: Synthesis, self-assembly, and applications. Polymers 2017, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.T.; Meeuwissen, S.A.; Nolte, R.J.; van Hest, J.C. Smart nanocontainers and nanoreactors. Nanoscale 2010, 2, 844–858. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.K.; Leal, C. Polymer–lipid hybrid materials. Chem. Rev. 2021, 121, 13996–14030. [Google Scholar] [CrossRef]
- Thoma, J.; Belegrinou, S.; Rossbach, P.; Grzelakowski, M.; Kita-Tokarczyk, K.; Meier, W. Membrane protein distribution in composite polymer—Lipid thin films. Chem. Commun. 2012, 48, 8811–8813. [Google Scholar] [CrossRef]
- Fauquignon, M.; Ibarboure, E.; Meins, J.-F. Membrane reinforcement in giant hybrid polymer lipid vesicles achieved by controlling the polymer architecture. Soft Matter 2021, 17, 83–89. [Google Scholar] [CrossRef]
- Gilbert, R.J.; Serra, M.D.; Froelich, C.J.; Wallace, M.I.; Anderluh, G. Membrane pore formation at protein—Lipid interfaces. Trends Biochem. Sci. 2014, 39, 510–516. [Google Scholar] [CrossRef]
- Hu, Z.; Ho, J.C.; Nallani, M. Synthetic (polymer) biology (membrane): Functionalization of polymer scaffolds for membrane proteins. Curr. Opin. Biotechnol. 2017, 46, 51–56. [Google Scholar] [CrossRef]
- Hindley, J.W.; Law, R.V.; Ces, O. Membrane functionalization in artificial cell engineering. SN Appl. Sci. 2020, 2, 593. [Google Scholar] [CrossRef] [Green Version]
- Hoang, N.H.; Lim, C.; Sim, T.; Oh, K.T. Triblock copolymers for nano-sized drug delivery systems. J. Pharm. Investig. 2017, 47, 27–35. [Google Scholar] [CrossRef]
- Hasannia, M.; Aliabadi, A.; Abnous, K.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Synthesis of block copolymers used in polymersome fabrication: Application in drug delivery. J. Control. Release 2022, 341, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Mora-Huertas, C.E.; Fessi, H.; Elaissari, A. Polymer-based nanocapsules for drug delivery. Int. J. Pharm. 2010, 385, 113–142. [Google Scholar] [CrossRef] [PubMed]
- Reimhult, E.; Virk, M.M. Hybrid lipopolymer vesicle drug delivery and release systems. J. Biomed. Res. 2021, 35, 301–309. [Google Scholar] [CrossRef]
- Goers, R.; Thoma, J.; Ritzmann, N.; Di Silvestro, A.; Alter, C.; Gunkel-Grabole, G.; Fotiadis, D.; Müller, D.J.; Meier, W. Optimized reconstitution of membrane proteins into synthetic membranes. Commun. Chem. 2018, 1, 35. [Google Scholar] [CrossRef]
- Belluati, A.; Craciun, I.; Liu, J.; Palivan, C.G. Nanoscale enzymatic compartments in tandem support cascade reactions in vitro. Biomacromolecules 2018, 19, 4023–4033. [Google Scholar] [CrossRef]
- Rideau, E.; Wurm, F.R.; Landfester, K. Self-assembly of giant unilamellar vesicles by film hydration methodologies. Adv. Biosyst. 2019, 3, 1800324. [Google Scholar] [CrossRef] [Green Version]
- Cabukusta, B.; Neefjes, J. Mechanisms of lysosomal positioning and movement. Traffic 2018, 19, 761–769. [Google Scholar] [CrossRef]
- Blanazs, A.; Armes, S.P.; Ryan, A.J. Self-assembled block copolymer aggregates: From micelles to vesicles and their biological applications. Macromol. Rapid Commun. 2009, 30, 267–277. [Google Scholar] [CrossRef]
- Discher, D.E. Polymer vesicles. Science 2002, 297, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Antonietti, M.; Förster, S. Vesicles and liposomes: A self-assembly principle beyond lipids. Adv. Mater. 2003, 15, 1323–1333. [Google Scholar] [CrossRef]
- Daubian, D.; Gaitzsch, J.; Meier, W. Synthesis and complex self-assembly of amphiphilic block copolymers with a branched hydrophobic Poly(2-Oxazoline) into multicompartment micelles, pseudo-vesicles and Yolk/Shell nanoparticles. Polym. Chem. 2020, 11, 1237–1248. [Google Scholar] [CrossRef] [Green Version]
- Wehr, R.; Gaitzsch, J.; Daubian, D.; Fodor, C.; Meier, W. Deepening the insight into poly(butylene oxide)-Block-poly(Glycidol) synthesis and self-assemblies: Micelles, worms and vesicles. RSC Adv. 2020, 10, 22701–22711. [Google Scholar] [CrossRef] [PubMed]
- Li, B.-Y.; Li, Y.-C.; Lu, Z.-Y. The important role of cosolvent in the amphiphilic diblock copolymer self-assembly process. Polymer 2019, 171, 1–7. [Google Scholar] [CrossRef]
- Balasubramanian, V.; Herranz-Blanco, B.; Almeida, P.V.; Hirvonen, J.; Santos, H.A. Multifaceted polymersome platforms: Spanning from self-assembly to drug delivery and protocells. Prog. Polym. Sci. 2016, 60, 51–85. [Google Scholar] [CrossRef]
- Daubian, D.; Fillion, A.; Gaitzsch, J.; Meier, W. One-pot synthesis of an amphiphilic ABC triblock copolymer PEO- b -PEHOx- b -PEtOz and its self-assembly into nanoscopic asymmetric polymersomes. Macromolecules 2020, 53, 11040–11050. [Google Scholar] [CrossRef]
- Parnell, A.J.; Tzokova, N.; Topham, P.D.; Adams, D.J.; Adams, S.; Fernyhough, C.M.; Ryan, A.J.; Jones, R.A. The efficiency of encapsulation within surface rehydrated polymersomes. Faraday Discuss. 2009, 143, 29–46. [Google Scholar] [CrossRef]
- Belluati, A.; Craciun, I.; Meyer, C.E.; Rigo, S.; Palivan, C.G. Enzymatic reactions in polymeric compartments: Nanotechnology meets nature. Curr. Opin. Biotechnol. 2019, 60, 53–62. [Google Scholar] [CrossRef]
- Pachioni-Vasconcelos, J.D.; Apolinário, A.C.; Lopes, A.M.; Pessoa, A., Jr.; Barbosa, L.R.; Rangel-Yagui, C.D. Compartmentalization of therapeutic proteins into semi-crystalline PEG-PCL polymersomes. Soft Mater. 2021, 19, 222–230. [Google Scholar] [CrossRef]
- Lomora, M.; Garni, M.; Itel, F.; Tanner, P.; Spulber, M.; Palivan, C.G. Polymersomes with engineered ion selective permeability as stimuli-responsive nanocompartments with preserved architecture. Biomaterials 2015, 53, 406–414. [Google Scholar] [CrossRef]
- Ibarboure, E.; Fauquignon, M.; Le Meins, J.-F. Obtention of giant unilamellar hybrid vesicles by electroformation and measurement of their mechanical properties by micropipette aspiration. J. Vis. Exp. 2020, 155, 60199. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant vesicles: Preparations and applications. ChemBioChem 2010, 11, 848–865. [Google Scholar] [CrossRef] [PubMed]
- Garni, M.; Wehr, R.; Avsar, S.Y.; John, C.; Palivan, C.; Meier, W. Polymer membranes as templates for bio-applications ranging from artificial cells to active surfaces. Eur. Polym. J. 2019, 112, 346–364. [Google Scholar] [CrossRef]
- Peyret, A.; Ibarboure, E.; Le Meins, J.-F.; Lecommandoux, S. Asymmetric hybrid polymer—Lipid giant vesicles as cell membrane mimics. Adv. Sci. 2018, 5, 1700453. [Google Scholar] [CrossRef]
- dos Santos, E.C.; Angelini, A.; Hürlimann, D.; Meier, W.; Palivan, C.G. Giant polymer compartments for confined reactions. Chemistry 2020, 2, 470–489. [Google Scholar] [CrossRef]
- Shum, H.C.; Kim, J.-W.; Weitz, D.A. Microfluidic fabrication of monodisperse biocompatible and biodegradable polymersomes with controlled permeability. J. Am. Chem. Soc. 2008, 130, 9543–9549. [Google Scholar] [CrossRef]
- Luo, R.; Göpfrich, K.; Platzman, I.; Spatz, J.P. DNA-based assembly of multi-compartment polymersome networks. Adv. Funct. Mater. 2020, 30, 2003480. [Google Scholar] [CrossRef]
- Deshpande, S.; Caspi, Y.; Meijering, A.E.; Dekker, C. Octanol-assisted liposome assembly on chip. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- dos Santos, E.C.; Belluati, A.; Necula, D.; Scherrer, D.; Meyer, C.E.; Wehr, R.P.; Lörtscher, E.; Palivan, C.G.; Meier, W. Combinatorial strategy for studying biochemical pathways in double emulsion templated cell-sized compartments. Adv. Mater. 2020, 32, 2004804. [Google Scholar] [CrossRef]
- Weiss, M.; Frohnmayer, J.P.; Benk, L.T.; Haller, B.; Janiesch, J.W.; Heitkamp, T.; Börsch, M.; Lira, R.B.; Dimova, R.; Lipowsky, R.; et al. Sequential bottom-up assembly of mechanically stabilized synthetic cells by microfluidics. Nat. Mater. 2018, 17, 89–95. [Google Scholar] [CrossRef]
- Phan, H.; Taresco, V.; Penelle, J.; Couturaud, B. Polymerisation-induced self-assembly (PISA) as a straightforward formulation strategy for stimuli-responsive drug delivery systems and biomaterials: Recent advances. Biomater. Sci. 2021, 9, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Dorsman, I.R.; Derry, M.J.; Cunningham, V.J.; Brown, S.L.; Williams, C.N.; Armes, S.P. Tuning the vesicle-to-worm transition for thermoresponsive block copolymer vesicles prepared via polymerisation-induced self-assembly. Polym. Chem. 2021, 12, 1224–1235. [Google Scholar] [CrossRef]
- Xu, S.; Corrigan, N.; Boyer, C. Forced gradient copolymerisation: A simplified approach for polymerisation-induced self-assembly. Polym. Chem. 2021, 12, 57–68. [Google Scholar] [CrossRef]
- Canning, S.L.; Smith, G.N.; Armes, S.P. A critical appraisal of RAFT-mediated polymerization-induced self-assembly. Macromolecules 2016, 49, 1985–2001. [Google Scholar] [CrossRef]
- Donath, E.; Sukhorukov, G.B.; Caruso, F.; Davis, S.A.; Möhwald, H. Novel hollow polymer shells by colloid-templated assembly of polyelectrolytes. Angew. Chem. Int. Ed. 1998, 37, 2201–2205. [Google Scholar] [CrossRef]
- Eivazi, A.; Medronho, B.; Lindman, B.; Norgren, M. On the development of all-cellulose capsules by vesicle-templated layer-by-layer assembly. Polymers 2021, 13, 589. [Google Scholar] [CrossRef]
- Elizarova, I.S.; Luckham, P.F. Layer-by-layer adsorption: Factors affecting the choice of substrates and polymers. Adv. Colloid Interface Sci. 2018, 262, 1–20. [Google Scholar] [CrossRef]
- Kurapati, R.; Groth, T.W.; Raichur, A.M. Recent developments in layer-by-layer technique for drug delivery applications. ACS Appl. Bio Mater. 2019, 2, 5512–5527. [Google Scholar] [CrossRef]
- Campbell, J.; Vikulina, A.S. Layer-by-layer assemblies of biopolymers: Build-Up, mechanical stability and molecular dynamics. Polymers 2020, 12, 1949. [Google Scholar] [CrossRef]
- Francesch, M.B. Capsosomes: The revolutionary enzyme carriers. Eur. Pharm. Rev. 2019, 24, 34–36. [Google Scholar]
- Städler, B.; Chandrawati, R.; Goldie, K.; Caruso, F. Capsosomes: Subcompartmentalizing polyelectrolyte capsules using liposomes. Langmuir 2009, 25, 6725–6732. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Z. Polyion complexes via electrostatic interaction of oppositely charged block copolymers. Macromolecules 2020, 53, 8737–8740. [Google Scholar] [CrossRef]
- Liu, Y.; Maruyama, T.; Kc, B.; Mori, T.; Katayama, Y.; Kishimura, A. Inducible dynamic behavior of polyion complex vesicles by disrupting charge balance. Chem. Lett. 2021, 50, 1034–1037. [Google Scholar] [CrossRef]
- Nishimura, T.; Sasaki, Y.; Akiyoshi, K. Biotransporting self-assembled nanofactories using polymer vesicles with molecular permeability for enzyme prodrug cancer therapy. Adv. Mater. 2017, 29, 1702406. [Google Scholar] [CrossRef] [PubMed]
- Odrobińska, J.; Gumieniczek-Chłopek, E.; Szuwarzyński, M.; Radziszewska, A.; Fiejdasz, S.; Strączek, T.; Kapusta, C.; Zapotoczny, S. Magnetically navigated core—Shell polymer capsules as nanoreactors loadable at the oil/water interface. ACS Appl. Mater. Interfaces 2019, 11, 10905–10913. [Google Scholar] [CrossRef]
- Cai, H.; Wang, P.; Zhang, D. PH-responsive linkages-enabled layer-by-layer assembled antibacterial and antiadhesive multilayer films with polyelectrolyte nanocapsules as biocide delivery vehicles. J. Drug Deliv. Sci. Technol. 2019, 54, 101251. [Google Scholar] [CrossRef]
- Svenskaya, Y.; Garello, F.; Lengert, E.; Kozlova, A.; Verkhovskii, R.; Bitonto, V.; Ruggiero, M.R.; German, S.; Gorin, D.; Terreno, E. Biodegradable polyelectrolyte/magnetite capsules for mr imaging and magnetic targeting of tumors. Nanotheranostics 2021, 5, 362–377. [Google Scholar] [CrossRef]
- Seitz, S.; Ajiro, H. Self-assembling weak polyelectrolytes for the layer-by-layer encapsulation of paraffin-type phase change material icosane. Sol. Energy Mater. Sol. Cells 2019, 190, 57–64. [Google Scholar] [CrossRef]
- Larrañaga, A.; Isa, I.L.; Patil, V.; Thamboo, S.; Lomora, M.; Fernández-Yague, M.A.; Sarasua, J.-R.; Palivan, C.G.; Pandit, A. Antioxidant functionalized polymer capsules to prevent oxidative stress. Acta Biomater. 2018, 67, 21–31. [Google Scholar] [CrossRef]
- Sharma, V.; Vijay, J.; Ganesh, M.R.; Sundaramurthy, A. Multilayer capsules encapsulating nimbin and doxorubicin for cancer chemo-photothermal therapy. Int. J. Pharm. 2020, 582, 119350. [Google Scholar] [CrossRef]
- Huang, J.; Li, C.; Gao, Y.; Cai, Y.; Guo, X.; Cohen Stuart, M.A.; Wang, J. Dendrimer-based polyion complex vesicles: Loops make loose. Macromol. Rapid Commun. 2022, 43, e2100594. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Cabral, H.; Toh, K.; Kishimura, A.; Kataoka, K. Robust polyion complex vesicles (PICsomes) under physiological conditions reinforced by multiple hydrogen bond formation derived by guanidinium groups. Biomacromolecules 2018, 19, 4113–4121. [Google Scholar] [CrossRef] [PubMed]
- Koval, V.; Morozova, E.; Revtovich, S.; Lyfenko, A.; Chobanian, A.; Timofeeva, V.; Solovieva, A.; Anufrieva, N.; Kulikova, V.; Demidkina, T. Characteristics and stability assessment of therapeutic methionine γ-Lyase-loaded polyionic vesicles. ACS Omega 2022, 7, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Sueyoshi, D.; Anraku, Y.; Komatsu, T.; Urano, Y.; Kataoka, K. Enzyme-loaded polyion complex vesicles as in vivo nanoreactors working sustainably under the blood circulation: Characterization and functional evaluation. Biomacromolecules 2017, 18, 1189–1196. [Google Scholar] [CrossRef]
- Li, J.; Anraku, Y.; Kataoka, K. Self-boosting catalytic nanoreactors integrated with triggerable crosslinking membrane networks for initiation of immunogenic cell death by pyroptosis. Angew. Chem. Int. Ed. 2020, 59, 13526–13530. [Google Scholar] [CrossRef]
- Seneviratne, R.; Catania, R.; Rappolt, M.; Jeuken, L.J.; Beales, P.A. Membrane mixing and dynamics in hybrid POPC/poly(1,2-butadiene-Block-ethylene oxide) (PBd-b-PEO) lipid/block co-polymer giant vesicles. Soft Matter 2022, 18, 1294–1301. [Google Scholar] [CrossRef]
- Peyret, A.; Ibarboure, E.; Pippa, N.; Lecommandoux, S. Liposomes in polymersomes: Multicompartment System with temperature-triggered release. Langmuir 2017, 33, 7079–7085. [Google Scholar] [CrossRef] [Green Version]
- Wehr, R.; dos Santos, E.C.; Muthwill, M.S.; Chimisso, V.; Gaitzsch, J.; Meier, W. Fully amorphous atactic and isotactic block copolymers and their self-assembly into nano- and microscopic vesicles. Polym. Chem. 2021, 12, 5377–5389. [Google Scholar] [CrossRef]
- Zhou, X.; Cornel, E.J.; Fan, Z.; He, S.; Du, J. Bone-targeting polymer vesicles for effective therapy of osteoporosis. Nano Lett. 2021, 21, 7998–8007. [Google Scholar] [CrossRef]
- Yang, Y.-Y.; Chen, L.-S.; Sun, M.; Wang, C.-Y.; Fan, Z.; Du, J.-Z. Biodegradable polypeptide-based vesicles with intrinsic blue fluorescence for antibacterial visualization. Chin. J. Polym. Sci. 2021, 39, 1412–1420. [Google Scholar] [CrossRef]
- Belluati, A.; Thamboo, S.; Najer, A.; Maffeis, V.; von Planta, C.; Craciun, I.; Palivan, C.G.; Meier, W. Multicompartment polymer vesicles with artificial organelles for signal-triggered cascade reactions including cytoskeleton formation. Adv. Funct. Mater. 2020, 30, 2002949. [Google Scholar] [CrossRef]
- Marušič, N.; Otrin, L.; Zhao, Z.; Lira, R.B.; Kyrilis, F.L.; Hamdi, F.; Kastritis, P.L.; Vidaković-Koch, T.; Ivanov, I.; Sundmacher, K.; et al. Constructing artificial respiratory chain in polymer compartments: Insights into the interplay between Bo3 oxidase and the membrane. Proc. Natl. Acad. Sci. USA 2020, 117, 15006–15017. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.K.; Kambar, N.; Leal, C. Hybrid unilamellar vesicles of phospholipids and block copolymers with crystalline domains. Polymers 2020, 12, 1232. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; McCabe, J.; Hill, K.; Beales, P.A. Biodegradable hybrid block copolymer—Lipid vesicles as potential drug delivery systems. J. Colloid Interface Sci. 2020, 562, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Xia, Y.; Shao, J.; Guo, B.; Dong, Y.; Pijpers, I.A.; Zhong, Z.; Meng, F.; Abdelmohsen, L.K.; Williams, D.S.; et al. Biodegradable polymersomes with structure inherent fluorescence and targeting capacity for enhanced photo-dynamic therapy. Angew. Chem. Int. Ed. 2021, 60, 17629–17637. [Google Scholar] [CrossRef]
- Ke, W.; Li, J.; Mohammed, F.; Wang, Y.; Tou, K.; Liu, X.; Wen, P.; Kinoh, H.; Anraku, Y.; Chen, H.; et al. Therapeutic polymersome nanoreactors with tumor-specific activable cascade reactions for cooperative cancer therapy. ACS Nano 2019, 13, 2357–2369. [Google Scholar] [CrossRef]
- Li, J.; Ge, Z.; Toh, K.; Liu, X.; Dirisala, A.; Ke, W.; Wen, P.; Zhou, H.; Wang, Z.; Xiao, S.; et al. Enzymatically transformable polymersome-based nanotherapeutics to eliminate minimal relapsable cancer. Adv. Mater. 2021, 33, 2105254. [Google Scholar] [CrossRef]
- Blackman, L.D.; Varlas, S.; Arno, M.C.; Houston, Z.H.; Fletcher, N.L.; Thurecht, K.J.; Hasan, M.; Gibson, M.I.; O’Reilly, R.K. Confinement of therapeutic enzymes in selectively permeable polymer vesicles by polymerization-induced self-assembly (pisa) reduces antibody binding and proteolytic susceptibility. ACS Cent. Sci. 2018, 4, 718–723. [Google Scholar] [CrossRef]
- Cao, J.; Wei, Y.; Zhang, Y.; Wang, G.; Ji, X.; Zhong, Z. Iodine-rich polymersomes enable versatile SPECT/CT imaging and potent radioisotope therapy for tumor in vivo. ACS Appl. Mater. Interfaces 2019, 11, 18953–18959. [Google Scholar] [CrossRef]
- Kim, B.S.; Naito, M.; Chaya, H.; Hori, M.; Hayashi, K.; Min, H.S.; Yi, Y.; Kim, H.J.; Nagata, T.; Anraku, Y.; et al. Noncovalent stabilization of vesicular polyion complexes with chemically modified/single-stranded oligonucleotides and peg-b-guanidinylated polypeptides for intracavity encapsulation of effector enzymes aimed at cooperative gene knockdown. Biomacromolecules 2020, 21, 4365–4376. [Google Scholar] [CrossRef]
- Choi, H.; Lee, G.-H.; Kim, K.S.; Hahn, S.K. Light-guided nanomotor systems for autonomous photothermal cancer therapy. ACS Appl. Mater. Interfaces 2018, 10, 2338–2346. [Google Scholar] [CrossRef]
- Sun, J.; Mathesh, M.; Li, W.; Wilson, D.A. Enzyme-powered nanomotors with controlled size for biomedical applications. ACS Nano 2019, 13, 10191–10200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pijpers, I.A.; Cao, S.; Llopis-Lorente, A.; Zhu, J.; Song, S.; Joosten, R.R.; Meng, F.; Friedrich, H.; Williams, D.S.; Sánchez, S.; et al. Hybrid biodegradable nanomotors through compartmentalized synthesis. Nano Lett. 2020, 20, 4472–4480. [Google Scholar] [CrossRef] [PubMed]
- Bueno, C.Z.; Apolinário, A.C.; Duro-Castano, A.; Poma, A.; Pessoa, A.; Rangel-Yagui, C.O.; Battaglia, G. L-asparaginase encapsulation into asymmetric permeable polymersomes. ACS Macro Lett. 2020, 9, 1471–1477. [Google Scholar] [CrossRef]
- Jin, S.-M.; Jeon, J.; Park, M.-K.; Kim, G.H.; Lee, E. Multicompartment vesicles formation by emulsification-induced assembly of poly(ethylene oxide)-block-poly(ε-Caprolactone) and their dual-loading capability. Macromol. Rapid Commun. 2018, 39, 1700545. [Google Scholar] [CrossRef] [PubMed]
- Konishcheva, E.V.; Daubian, D.; Rigo, S.; Meier, W.P. Probing membrane asymmetry of abc polymersomes. Chem. Commun. 2019, 55, 1148–1151. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Sun, H.; Du, J. Sugar-breathing glycopolymersomes for regulating glucose level. J. Am. Chem. Soc. 2017, 139, 7640–7647. [Google Scholar] [CrossRef]
- Askes, S.H.; Pomp, W.; Hopkins, S.L.; Kros, A.; Wu, S.; Schmidt, T.; Bonnet, S. Imaging upconverting polymersomes in cancer cells: Biocompatible antioxidants brighten triplet-triplet annihilation upconversion. Small Weinh. Bergstr. Ger. 2016, 12, 5579–5590. [Google Scholar] [CrossRef] [Green Version]
- Kulikova, V.V.; Morozova, E.A.; Anufrieva, N.V.; Koval, V.S.; Lyfenko, A.D.; Lesnova, E.I.; Kushch, A.A.; Revtovich, S.V.; Demidkina, T.V. Kinetic and pharmacokinetic characteristics of therapeutic methinonine γ-Lyase encapsulated in polyion complex vesicles. Biochimie 2022, 194, 13–18. [Google Scholar] [CrossRef]
- Garni, M.; Einfalt, T.; Goers, R.; Palivan, C.G.; Meier, W. Live follow-up of enzymatic reactions inside the cavities of synthetic giant unilamellar vesicles equipped with membrane proteins mimicking cell architecture. ACS Synth. Biol. 2018, 7, 2116–2125. [Google Scholar] [CrossRef]
- Maffeis, V.; Belluati, A.; Craciun, I.; Wu, D.; Novak, S.; Schoenenberger, C.-A.; Palivan, C.G. Clustering of catalytic nanocompartments for enhancing an extracellular non-native cascade reaction. Chem. Sci. 2021, 12, 12274–12285. [Google Scholar] [CrossRef] [PubMed]
- Thamboo, S.; Najer, A.; Belluati, A.; von Planta, C.; Wu, D.; Craciun, I.; Meier, W.; Palivan, C.G. Mimicking cellular signaling pathways within synthetic multicompartment vesicles with triggered enzyme activity and induced ion channel recruitment. Adv. Funct. Mater. 2019, 29, 1904267. [Google Scholar] [CrossRef]
- Messager, L.; Burns, J.R.; Kim, J.; Cecchin, D.; Hindley, J.; Pyne, A.L.; Gaitzsch, J.; Battaglia, G.; Howorka, S. Biomimetic hybrid nanocontainers with selective permeability. Angew. Chem. Int. Ed. 2016, 55, 11106–11109. [Google Scholar] [CrossRef] [PubMed]
- Hosta-Rigau, L.; York-Duran, M.J.; Zhang, Y.; Goldie, K.N.; Städler, B. Confined Multiple enzymatic (cascade) reactions within poly(dopamine)-based capsosomes. ACS Appl. Mater. Interfaces 2014, 6, 12771–12779. [Google Scholar] [CrossRef]
- Matoori, S.; Bao, Y.; Schmidt, A.; Fischer, E.J.; Ochoa-Sanchez, R.; Tremblay, M.; Oliveira, M.M.; Rose, C.F.; Leroux, J.-C. An investigation of PS-b-PEO polymersomes for the oral treatment and diagnosis of hyperammonemia. Small 2019, 15, 1902347. [Google Scholar] [CrossRef]
- Kulygin, O.; Price, A.D.; Chong, S.-F.; Städler, B.; Zelikin, A.N.; Caruso, F. Subcompartmentalized polymer hydrogel capsules with selectively degradable carriers and subunits. Small 2010, 6, 1558–1564. [Google Scholar] [CrossRef]
- Nishimura, T.; Toh, W.-L.; Akiyoshi, K. Synthesis and characterization of shell-cross-linked glycopolymer bilayer vesicles. Macromol. Rapid Commun. 2018, 39, 1800384. [Google Scholar] [CrossRef]
- Liu, D.; Sun, H.; Xiao, Y.; Chen, S.; Cornel, E.J.; Zhu, Y.; Du, J. Design principles, synthesis and biomedical applications of polymer vesicles with inhomogeneous membranes. J. Control. Release 2020, 326, 365–386. [Google Scholar] [CrossRef]
- Zhong, Y.; Meng, F.; Zhang, W.; Li, B.; van Hest, J.C.; Zhong, Z. CD44-targeted vesicles encapsulating granzyme B as artificial killer cells for potent inhibition of human multiple myeloma in mice. J. Control. Release 2020, 320, 421–430. [Google Scholar] [CrossRef]
- Dinda, S.; Sarkar, S.; Das, P.K. Glucose oxidase mediated targeted cancer-starving therapy by biotinylated self-assembled vesicles. Chem. Commun. 2018, 54, 9929–9932. [Google Scholar] [CrossRef] [Green Version]
- van Beek, L.F.; Welzen, P.L.; Teufel, L.U.; Joosten, I.; Diavatopoulos, D.A.; van Hest, J.; de Jonge, M.I. Bimodal targeting of human leukocytes by Fc- and CpG-decorated polymersomes to tune immune induction. Biomacromolecules 2021, 22, 4422–4433. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Sun, Y.; Guo, B.; Wei, Y.; Xia, Y.; Huangfu, Z.; Meng, F.; van Hest, J.C.; Yuan, J.; Zhong, Z. A3β1 integrin-targeting polymersomal docetaxel as an advanced nanotherapeutic for nonsmall cell lung cancer treatment. ACS Appl. Mater. Interfaces 2020, 12, 14905–14913. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.E.; Liu, J.; Craciun, I.; Wu, D.; Wang, H.; Xie, M.; Fussenegger, M.; Palivan, C.G. Segregated nanocompartments containing therapeutic enzymes and imaging compounds within dna-zipped polymersome clusters for advanced nanotheranostic platform. Small 2020, 16, 1906492. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jia, H.; Tham, H.P.; Qu, Q.; Xing, P.; Zhao, J.; Phua, S.Z.; Chen, G.; Zhao, Y. Theranostic prodrug vesicles for imaging guided codelivery of camptothecin and sirna in synergetic cancer therapy. ACS Appl. Mater. Interfaces 2017, 9, 23536–23543. [Google Scholar] [CrossRef] [PubMed]
- Egli, S.; Schlaad, H.; Bruns, N.; Meier, W. Functionalization of block copolymer vesicle surfaces. Polymers 2011, 3, 252–280. [Google Scholar] [CrossRef] [Green Version]
- van Dongen, S.F.; Nallani, M.; Cornelissen, J.J.; Nolte, R.J.; van Hest, J.C. A three-enzyme cascade reaction through positional assembly of enzymes in a polymersome nanoreactor. Chem. Eur. J. 2009, 15, 1107–1114. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Qian, Y.; Yu, Y.; Liu, G.; Hu, J.; Zhang, G.; Liu, S. Engineering intracellular delivery nanocarriers and nanoreactors from oxidation-responsive polymersomes via synchronized bilayer cross-linking and permeabilizing inside live cells. J. Am. Chem. Soc. 2016, 138, 10452–10466. [Google Scholar] [CrossRef]
- Anajafi, T.; Yu, J.; Sedigh, A.; Haldar, M.K.; Muhonen, W.W.; Oberlander, S.; Wasness, H.; Froberg, J.; Molla, M.S.; Katti, K.S.; et al. Nuclear localizing peptide-conjugated, redox-sensitive polymersomes for delivering curcumin and doxorubicin to pancreatic cancer microtumors. Mol. Pharm. 2017, 14, 1916–1928. [Google Scholar] [CrossRef]
- Scheerstra, J.; Wauters, A.; Tel, J.; Abdelmohsen, L.; van Hest, J. Polymersomes as a potential platform for cancer immunotherapy. Mater. Today Adv. 2022, 13, 100203. [Google Scholar] [CrossRef]
- Fujita, S.; Motoda, Y.; Kigawa, T.; Tsuchiya, K.; Numata, K. Peptide-based polyion complex vesicles that deliver enzymes into intact plants to provide antibiotic resistance without genetic modification. Biomacromolecules 2021, 22, 1080–1090. [Google Scholar] [CrossRef]
- Gunkel-Grabole, G.; Palivan, C.; Meier, W. Nanostructured surfaces through immobilization of self-assembled polymer architectures using thiol–ene chemistry. Macromol. Mater. Eng. 2017, 302, 1600363. [Google Scholar] [CrossRef]
- Rigo, S.; Gunkel-Grabole, G.; Meier, W.; Palivan, C.G. Surfaces with dual functionality through specific coimmobilization of self-assembled polymeric nanostructures. Langmuir 2019, 35, 4557–4565. [Google Scholar] [CrossRef] [PubMed]
- Simon-Gracia, L.; Hunt, H.; Scodeller, P.D.; Gaitzsch, J.; Braun, G.B.; Willmore, A.-M.; Ruoslahti, E.; Battaglia, G.; Teesalu, T. Paclitaxel-loaded polymersomes for enhanced intraperitoneal chemotherapy. Mol. Cancer Ther. 2016, 15, 670–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leong, J.; Teo, J.Y.; Aakalu, V.K.; Yang, Y.Y.; Kong, H. Engineering polymersomes for diagnostics and therapy. Adv. Healthc. Mater. 2018, 7, 1701276. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.; Boye, S.; Ajeilat, H.G.; Michen, S.; Tietze, S.; Voit, B.; Lederer, A.; Temme, A.; Appelhans, D. Multivalent protein-loaded ph-stable polymersomes: First step toward protein targeted therapeutics. Macromol. Biosci. 2021, 21, 2100102. [Google Scholar] [CrossRef]
- Egli, S.; Nussbaumer, M.G.; Balasubramanian, V.; Chami, M.; Bruns, N.; Palivan, C.; Meier, W. Biocompatible functionalization of polymersome surfaces: A new approach to surface immobilization and cell targeting using polymersomes. J. Am. Chem. Soc. 2011, 133, 4476–4483. [Google Scholar] [CrossRef]
- Rijpkema, S.J.; Langens, S.G.; van der Kolk, M.R.; Gavriel, K.; Toebes, B.J.; Wilson, D.A. Modular approach to the functionalization of polymersomes. Biomacromolecules 2020, 21, 1853–1864. [Google Scholar] [CrossRef]
- Moulahoum, H.; Ghorbanizamani, F.; Zihnioglu, F.; Timur, S. Surface biomodification of liposomes and polymersomes for efficient targeted drug delivery. Bioconjug. Chem. 2021, 32, 1491–1502. [Google Scholar] [CrossRef]
- Craciun, I.; Denes, A.S.; Gunkel-Grabole, G.; Belluati, A.; Palivan, C.G. Surfaces decorated with polymeric nanocompartments for PH reporting. Helv. Chim. Acta 2018, 101, e1700290. [Google Scholar] [CrossRef]
- Liu, J.; Craciun, I.; Belluati, A.; Wu, D.; Sieber, S.; Einfalt, T.; Witzigmann, D.; Chami, M.; Huwyler, J.; Palivan, C.G. DNA-directed arrangement of soft synthetic compartments and their behavior in vitro and in vivo. Nanoscale 2020, 12, 9786–9799. [Google Scholar] [CrossRef] [Green Version]
- Chandrawati, R. Layer-by-layer engineered polymer capsules for therapeutic delivery. In Biomaterials for Tissue Engineering: Methods and Protocols; Chawla, K., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; pp. 73–84. ISBN 978-1-4939-7741-3. [Google Scholar]
- Kawamura, W.; Miura, Y.; Kokuryo, D.; Toh, K.; Yamada, N.; Nomoto, T.; Matsumoto, Y.; Sueyoshi, D.; Liu, X.; Aoki, I.; et al. Density-tunable conjugation of cyclic rgd ligands with polyion complex vesicles for the neovascular imaging of orthotopic glioblastomas. Sci. Technol. Adv. Mater. 2015, 16, 035004. [Google Scholar] [CrossRef] [PubMed]
- Hammer, D.A.; Robbins, G.P.; Haun, J.B.; Lin, J.J.; Qi, W.; Smith, L.A.; Peter Ghoroghchian, P.; Therien, M.J.; Bates, F.S. Leuko-polymersomes. Faraday Discuss. 2008, 139, 129. [Google Scholar] [CrossRef] [PubMed]
- Di Leone, S.; Vallapurackal, J.; Yorulmaz Avsar, S.; Kyropolou, M.; Ward, T.R.; Palivan, C.G.; Meier, W. Expanding the potential of the solvent-assisted method to create bio-interfaces from amphiphilic block copolymers. Biomacromolecules 2021, 22, 3005–3016. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Postupalenko, V.; Lörcher, S.; Wu, D.; Chami, M.; Meier, W.; Palivan, C.G. DNA-mediated self-organization of polymeric nanocompartments leads to interconnected artificial organelles. Nano Lett. 2016, 16, 7128–7136. [Google Scholar] [CrossRef]
- Deng, N.-N.; Yelleswarapu, M.; Zheng, L.; Huck, W.T. Microfluidic assembly of monodisperse vesosomes as artificial cell models. J. Am. Chem. Soc. 2017, 139, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Rasoulianboroujeni, M.; Kupgan, G.; Moghadam, F.; Tahriri, M.; Boughdachi, A.; Khoshkenar, P.; Ambrose, J.J.; Kiaie, N.; Vashaee, D.; Ramsey, J.D.; et al. Development of a DNA-liposome complex for gene delivery applications. Mater. Sci. Eng. C 2017, 75, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Shi, Z.; Zhang, Y.; Chen, X.-J.; Han, S.-Y.; Baumgart, T.; Chenoweth, D.M.; Park, S.-J. DNA island formation on binary block copolymer vesicles. J. Am. Chem. Soc. 2016, 138, 10157–10162. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.E.; Schoenenberger, C.-A.; Liu, J.; Craciun, I.; Palivan, C.G. DNA-tethered polymersome clusters as nanotheranostic platform. CHIMIA 2021, 75, 296. [Google Scholar] [CrossRef]
- Arun, Y.; Ghosh, R.; Domb, A.J. Biodegradable hydrophobic injectable polymers for drug delivery and regenerative medicine. Adv. Funct. Mater. 2021, 31, 2010284. [Google Scholar] [CrossRef]
- Yorulmaz Avsar, S.; Kyropoulou, M.; Di Leone, S.; Schoenenberger, C.-A.; Meier, W.P.; Palivan, C.G. Biomolecules turn self-assembling amphiphilic block co-polymer platforms into biomimetic interfaces. Front. Chem. 2019, 6, 645. [Google Scholar] [CrossRef]
- De Leo, V.; Milano, F.; Agostiano, A.; Catucci, L. Recent advancements in polymer/liposome assembly for drug delivery: From surface modifications to hybrid vesicles. Polymers 2021, 13, 1027. [Google Scholar] [CrossRef]
- Rideau, E.; Dimova, R.; Schwille, P.; Wurm, F.R.; Landfester, K. Liposomes and polymersomes: A comparative review towards cell mimicking. Chem. Soc. Rev. 2018, 47, 8572–8610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itel, F.; Najer, A.; Palivan, C.G.; Meier, W. Dynamics of membrane proteins within synthetic polymer membranes with large hydrophobic mismatch. Nano Lett. 2015, 15, 3871–3878. [Google Scholar] [CrossRef] [PubMed]
- Lomora, M.; Itel, F.; Dinu, I.A.; Palivan, C.G. Selective ion-permeable membranes by insertion of biopores into polymersomes. Phys. Chem. Chem. Phys. 2015, 17, 15538–15546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garni, M.; Thamboo, S.; Schoenenberger, C.-A.; Palivan, C.G. Biopores/membrane proteins in synthetic polymer membranes. Biochim. Biophys. Acta BBA Biomembr. 2017, 1859, 619–638. [Google Scholar] [CrossRef]
- Draghici, C.; Kowal, J.; Darjan, A.; Meier, W.; Palivan, C.G. “Active surfaces” formed by immobilization of enzymes on solid-supported polymer membranes. Langmuir 2014, 30, 11660–11669. [Google Scholar] [CrossRef]
- Einfalt, T.; Witzigmann, D.; Edlinger, C.; Sieber, S.; Goers, R.; Najer, A.; Spulber, M.; Onaca-Fischer, O.; Huwyler, J.; Palivan, C.G. Biomimetic artificial organelles with in vitro and in vivo activity triggered by reduction in microenvironment. Nat. Commun. 2018, 9, 1127. [Google Scholar] [CrossRef] [Green Version]
- Edlinger, C.; Einfalt, T.; Spulber, M.; Car, A.; Meier, W.; Palivan, C.G. Biomimetic strategy to reversibly trigger functionality of catalytic nanocompartments by the insertion of PH-responsive biovalves. Nano Lett. 2017, 17, 5790–5798. [Google Scholar] [CrossRef]
- Sun, X.; Liu, C.; Liu, D.; Li, P.; Zhang, N. Novel biomimetic vectors with endosomal-escape agent enhancing gene transfection efficiency. Int. J. Pharm. 2012, 425, 62–72. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Hoekstra, D.; Zuhorn, I.S. Mechanism of polyplex- and lipoplex-mediated delivery of nucleic acids: Real-time visualization of transient membrane destabilization without endosomal lysis. ACS Nano 2013, 7, 3767–3777. [Google Scholar] [CrossRef]
- Wong, A.S.; Mann, S.K.; Czuba, E.; Sahut, A.; Liu, H.; Suekama, T.C.; Bickerton, T.; Johnston, A.P.; Such, G.K. Self-assembling dual component nanoparticles with endosomal escape capability. Soft Matter 2015, 11, 2993–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaur, D.; Dubey, N.C.; Tripathi, B.P. Biocatalytic self-assembled synthetic vesicles and coacervates: From single compartment to artificial cells. Adv. Colloid Interface Sci. 2022, 299, 102566. [Google Scholar] [CrossRef] [PubMed]
- Belluati, A.; Mikhalevich, V.; Yorulmaz Avsar, S.; Daubian, D.; Craciun, I.; Chami, M.; Meier, W.P.; Palivan, C.G. How do the properties of amphiphilic polymer membranes influence the functional insertion of peptide pores? Biomacromolecules 2020, 21, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Ma, W.; Zhao, X.; Wang, J.; Yao, L.; Jiang, X.; Wu, Z. Interaction between surface charge-modified gold nanoparticles and phospholipid membranes. Langmuir 2018, 34, 12583–12589. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 5577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, N.; Cao, J.; Lee, J.; Hlaing, S.P.; Oshi, M.A.; Naeem, M.; Ki, M.-H.; Lee, B.L.; Jung, Y.; Yoo, J.-W. Bacteria-targeted clindamycin loaded polymeric nanoparticles: Effect of surface charge on nanoparticle adhesion to MRSA, antibacterial activity, and wound healing. Pharmaceutics 2019, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Clavadetscher, J.; Lee, D.-K.; Chankeshwara, S.; Bradley, M.; Cho, W.-S. Surface charge-dependent cellular uptake of polystyrene nanoparticles. Nanomaterials 2018, 8, 1028. [Google Scholar] [CrossRef] [Green Version]
- Du, X.-J.; Wang, J.-L.; Iqbal, S.; Li, H.-J.; Cao, Z.-T.; Wang, Y.-C.; Du, J.-Z.; Wang, J. The effect of surface charge on oral absorption of polymeric nanoparticles. Biomater. Sci. 2018, 6, 642–650. [Google Scholar] [CrossRef]
- Kohli, A.K.; Alpar, H.O. Potential use of nanoparticles for transcutaneous vaccine delivery: Effect of particle size and charge. Int. J. Pharm. 2004, 275, 13–17. [Google Scholar] [CrossRef]
- El Badawy, A.M.; Silva, R.G.; Morris, B.; Scheckel, K.G.; Suidan, M.T.; Tolaymat, T.M. Surface charge-dependent toxicity of silver nanoparticles. Environ. Sci. Technol. 2011, 45, 283–287. [Google Scholar] [CrossRef]
- Deprey, K.; Becker, L.; Kritzer, J.; Plückthun, A. Trapped! A critical evaluation of methods for measuring total cellular uptake versus cytosolic localization. Bioconjug. Chem. 2019, 30, 1006–1027. [Google Scholar] [CrossRef]
- Sharma, A.R.; Kundu, S.K.; Nam, J.-S.; Sharma, G.; Priya Doss, C.G.; Lee, S.-S.; Chakraborty, C. Next generation delivery system for proteins and genes of therapeutic purpose: Why and how? BioMed Res. Int. 2014, 2014, e327950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleineberg, C.; Wölfer, C.; Abbasnia, A.; Pischel, D.; Bednarz, C.; Ivanov, I.; Heitkamp, T.; Börsch, M.; Sundmacher, K.; Vidaković-Koch, T. Light-driven ATP regeneration in diblock/grafted hybrid vesicles. ChemBioChem 2020, 21, 2149–2160. [Google Scholar] [CrossRef]
- Khan, S.; Li, M.; Muench, S.P.; Jeuken, L.J.C.; Beales, P.A. Durable proteo-hybrid vesicles for the extended functional lifetime of membrane proteins in bionanotechnology. Chem. Commun. 2016, 52, 11020–11023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otrin, L.; Witkowska, A.; Marušič, N.; Zhao, Z.; Lira, R.B.; Kyrilis, F.L.; Hamdi, F.; Ivanov, I.; Lipowsky, R.; Kastritis, P.L.; et al. En route to dynamic life processes by SNARE-mediated fusion of polymer and hybrid membranes. Nat. Commun. 2021, 12, 4972. [Google Scholar] [CrossRef] [PubMed]
- Artificial Melanogenesis by Confining Melanin/Polydopamine Production inside Polymersomes-Meyer-2021-Macromolecular Bioscience-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/mabi.202100249 (accessed on 27 April 2022).
- de Freitas, C.F.; Calori, I.R.; da Silva, A.C.P.; de Castro, L.V.; Sato, F.; Pellosi, D.S.; Tessaro, A.L.; Caetano, W.; Hioka, N. PEG-coated vesicles from pluronic/lipid mixtures for the carrying of photoactive erythrosine derivatives. Colloids Surf. B Biointerfaces 2019, 175, 530–544. [Google Scholar] [CrossRef]
- Belluati, A.; Craciun, I.; Palivan, C.G. Bioactive catalytic nanocompartments integrated into cell physiology and their amplification of a native signaling cascade. ACS Nano 2020, 14, 12101–12112. [Google Scholar] [CrossRef]
- Catalytic Polymersomes to Produce Strong and Long-Lasting Bioluminescence—Nanoscale (RSC Publishing). Available online: https://pubs.rsc.org/en/content/articlelanding/2021/NR/D0NR07178A#!divAbstract (accessed on 27 April 2022).
- Korpidou, M.; Maffeis, V.; Dinu, I.A.; Schoenenberger, C.-A.; Meier, W.P.; Palivan, C.G. Inverting glucuronidation of hymecromone in situ by catalytic nanocompartments. J. Mater. Chem. B 2022. [Google Scholar] [CrossRef]
- Langowska, K.; Palivan, C.G.; Meier, W. Polymer nanoreactors shown to produce and release antibiotics locally. Chem. Commun. 2012, 49, 128–130. [Google Scholar] [CrossRef]
- Meyer, C.E.; Schoenenberger, C.-A.; Wehr, R.P.; Wu, D.; Palivan, C.G. Artificial melanogenesis by confining melanin/polydopamine production inside polymersomes. Macromol. Biosci. 2021, 21, 2100249. [Google Scholar] [CrossRef]
- Wang, X.; Zhong, X.; Li, J.; Liu, Z.; Cheng, L. Inorganic nanomaterials with rapid clearance for biomedical applications. Chem. Soc. Rev. 2021, 50, 8669–8742. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Yang, N.; Wang, X.; Zhao, Q.; Chen, Q.; Liu, Z.; Cheng, L. Tumor microenvironment-responsive intelligent nanoplatforms for cancer theranostics. Nano Today 2020, 32, 100851. [Google Scholar] [CrossRef]
- Cheng, P.; Pu, K. Molecular imaging and disease theranostics with renal-clearable optical agents. Nat. Rev. Mater. 2021, 6, 1095–1113. [Google Scholar] [CrossRef]
- Imaging Upconverting Polymersomes in Cancer Cells: Biocompatible Antioxidants Brighten Triplet–Triplet Annihilation Upconversion-Askes-2016-Small-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/10.1002/smll.201601708 (accessed on 31 March 2022).
- Askes, S.H.; Bossert, N.; Bussmann, J.; Talens, V.S.; Meijer, M.S.; Kieltyka, R.E.; Kros, A.; Bonnet, S.; Heinrich, D. Dynamics of dual-fluorescent polymersomes with durable integrity in living cancer cells and zebrafish embryos. Biomaterials 2018, 168, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Camblin, M.; Detampel, P.; Kettiger, H.; Wu, D.; Balasubramanian, V.; Huwyler, J. Polymersomes containing quantum dots for cellular imaging. Int. J. Nanomed. 2014, 9, 2287–2298. [Google Scholar] [CrossRef] [Green Version]
- Iodine-Rich Polymersomes Enable Versatile SPECT/CT Imaging and Potent Radioisotope Therapy for Tumor in Vivo. ACS Applied Materials & Interfaces. Available online: https://pubs.acs.org/doi/10.1021/acsami.9b04294 (accessed on 31 March 2022).
- Lu, L.; Wang, Y.; Zhang, F.; Chen, M.; Lin, B.; Duan, X.; Cao, M.; Zheng, C.; Mao, J.; Shuai, X.; et al. MRI-Visible SiRNA nanomedicine directing neuronal differentiation of neural stem cells in stroke. Adv. Funct. Mater. 2018, 28, 1706769. [Google Scholar] [CrossRef]
- Meeuwissen, S.A.; Kim, K.T.; Chen, Y.; Pochan, D.J.; van Hest, J.C.M. Controlled shape transformation of polymersome stomatocytes. Angew. Chem. Int. Ed. 2011, 50, 7070–7073. [Google Scholar] [CrossRef]
- de Martino, M.T.; Tonin, F.; Bloemendal, V.R.; Hanefeld, U.; Rutjes, F.P.; van Hest, J.C. Compartmentalized cross-linked enzyme nano aggregates (c-CLEnAs) toward pharmaceutical transformations. RSC Adv. 2021, 11, 21857–21861. [Google Scholar] [CrossRef]
- Abdelmohsen, L.K.; Nijemeisland, M.; Pawar, G.M.; Janssen, G.-J.; Nolte, R.J.; van Hest, J.C.; Wilson, D.A. Dynamic loading and unloading of proteins in polymeric stomatocytes: Formation of an enzyme-loaded supramolecular nanomotor. ACS Nano 2016, 10, 2652–2660. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Tu, Y.; Wilson, D.A. Micro/nanomotors towards in vivo application: Cell, tissue and biofluid. Chem. Soc. Rev. 2017, 46, 5289–5310. [Google Scholar] [CrossRef]
- Lee, K.Y.; Park, S.-J.; Lee, K.A.; Kim, S.-H.; Kim, H.; Meroz, Y.; Mahadevan, L.; Jung, K.-H.; Ahn, T.K.; Parker, K.K.; et al. Photosynthetic artificial organelles sustain and control ATP-dependent reactions in a protocellular system. Nat. Biotechnol. 2018, 36, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fei, J.; Li, G.; Yuan, T.; Li, J. Compartmentalized assembly of motor protein reconstituted on protocell membrane toward highly efficient photophosphorylation. ACS Nano 2017, 11, 10175–10183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Kong, W.; Wang, S. Reconstitution of Caveolin-1 into artificial lipid membrane: Characterization by transmission electron microscopy and solid-state nuclear magnetic resonance. Molecules 2021, 26, 6201. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fei, J.; Xu, Y.; Hong, J.-D.; Li, J. Proton-consumed nanoarchitectures toward sustainable and efficient photophosphorylation. J. Colloid Interface Sci. 2019, 535, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, X.; Mu, W.; Han, X. Chemical signal communication between two protoorganelles in a lipid-based artificial cell. Anal. Chem. 2019, 91, 6859–6864. [Google Scholar] [CrossRef]
- Simons, K.; Sampaio, J.L. Membrane organization and lipid rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef]
- Devaux, P.F. Static and dynamic lipid asymmetry in cell membranes. Biochemistry 1991, 30, 1163–1173. [Google Scholar] [CrossRef]
- Rothman, J.E.; Lenard, J. Membrane asymmetry. Science 1977, 195, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Arriaga, L.R.; Huang, Y.; Kim, S.H.; Aragones, J.L.; Ziblat, R.; Koehler, S.A.; Weitz, D.A. Single-step assembly of asymmetric vesicles. Lab. Chip 2019, 19, 749–756. [Google Scholar] [CrossRef]
- Kamiya, K.; Kawano, R.; Osaki, T.; Akiyoshi, K.; Takeuchi, S. Cell-sized asymmetric lipid vesicles facilitate the investigation of asymmetric membranes. Nat. Chem. 2016, 8, 881–889. [Google Scholar] [CrossRef]
- Lu, L.; Schertzer, J.W.; Chiarot, P.R. Continuous microfluidic fabrication of synthetic asymmetric vesicles. Lab. Chip 2015, 15, 3591–3599. [Google Scholar] [CrossRef] [PubMed]
- Heberle, F.A.; Feigenson, G.W. Phase Separation in lipid membranes. Cold Spring Harb. Perspect. Biol. 2011, 3, a004630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Cabrera, L.A.; Pardo-López, L. Membrane remodeling and organization: Elements common to prokaryotes and eukaryotes. IUBMB Life 2017, 69, 55–62. [Google Scholar] [CrossRef] [Green Version]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- McConnell, H.M.; Kornberg, R.D. Inside-outside transitions of phospholipids in vesicle membranes. Biochemistry 1971, 10, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Pagano, R.E. Measurement of spontaneous transfer and transbilayer movement of BODIPY-labeled lipids in lipid vesicles. Biochemistry 1997, 36, 8840–8848. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.H.; Silver, P.A. Designing biological compartmentalization. Trends Cell Biol. 2012, 22, 662–670. [Google Scholar] [CrossRef]
- von Ballmoos, C.; Wiedenmann, A.; Dimroth, P. Essentials for ATP synthesis by F1F0 ATP synthases. Annu. Rev. Biochem. 2009, 78, 649–672. [Google Scholar] [CrossRef] [Green Version]
- Itel, F.; Chami, M.; Najer, A.; Lörcher, S.; Wu, D.; Dinu, I.A.; Meier, W. Molecular organization and dynamics in polymersome membranes: A lateral diffusion study. Macromolecules 2014, 47, 7588–7596. [Google Scholar] [CrossRef]
- Heuberger, L.; Palivan, C. Block copolymer giant unilamellar vesicles for high-throughput screening. CHIMIA 2022, 76, 350. [Google Scholar] [CrossRef]





| Biomolecule | Polymer | Location in Assembly | Application/Function |
|---|---|---|---|
| Actin | PMOXA-b-PDMS-b-PMOXA [71] | Encapsulated (in GUVs) | Polymerization to form a cytoskeleton |
| ATP synthase | PDMS-g-PEO, PBd-b-PEO [72,154] | Incorporated within membrane (GUVs) | ATP generation |
| Bacteriorhodopsin | PDMS-g-PEO, PBd- b-PEO [154] | Incorporated within membrane (GUVs) | Pumping protons across membrane |
| Catalase | PEG-b-PS [82], PAH and DEX [59] | Encapsulated with the stomata of polymer stomatocytes and LbL capsules | Conversion of hydrogen peroxide to oxygen and water for self-propelled movement |
| Cholesterol–DNA | PEO-b-PPO-b-PEO (Pluronics L121), PBd-b-PEG, PLA-b-PEG [37] | Incorporated within membrane (GUVs) | Clustering of polymersomes |
| Cytochrome bo3 ubiquinol oxidase (Cyt bo3) | PBd–PEO:POPC hybrid [155], PDMS-g-PEO and PDMS-g-PEO/PC hybrid [72,156] | Incorporated within membrane (polymersomes, GUVs) | Pumping protons across membrane |
| DNA nanopore NP-3c | PMPC-b-PDPA [93] | Incorporated within membrane (GUVs) | Pore formation for cross-membrane diffusion |
| Dopa decarboxylase (DDC) | PMOXA-b-PDMS [157] | Encapsulated (in polymersomes) | Production of dopamine |
| Erythrosine B (and its ester derivatives) | F127 Pluronic (mixed with DPPC lipids) [158] | Incorporated within membrane (polymersomes) | Photodynamic therapy |
| Glucose oxidase (Gox) | PMOXA-b-PDMS [39,91], PEG-b-P(CPTKMA-co-PEMA) [76], PATK and PEG-b-Pasp [65] | Encapsulated (in GUVs, polymersomes, and PICsomes) | Catalysis of glucose oxidation to hydrogen peroxide and D-glucono-δ-lactone |
| Gramicidin | PMOXA-b-PDMS [39], PMOXA-b-PDMS-b-PMOXA [71] | Incorporated within membrane | Membrane permeabilization towards ions |
| Horseradish peroxidase (HRP) | PMOXA-b-PDMS [39], PMOXA-b-PDMS-b-PMOXA [90,138], carbohydrate-b-PPG [54] | Encapsulated (in GUVs, polymersomes, capsosomes) | Catalysis of oxidation of organic substrates by hydrogen peroxide |
| Icosane | PAA and PAH (LbL) [58] | Encapsulated (in capsules) | Acting as a phase change material for thermal energy storage |
| Inducible nitric oxide synthase (iNOS) | PMOXA-b-PDMS-b-PMOXA [159] | Encapsulated (in polymersomes) | Oxidation of l-arginine to l-citrulline and nitric oxide (NO) |
| Ionomycin | PMOXA-b-PDMS-b-PMOXA [71] | Incorporated within membrane | Membrane permeabilization towards ions |
| Laccase | PMOXA-b-PDMS [120] | Encapsulated (in polymersomes) | Oxidation of phenolic and nonphenolic compounds |
| Lactoperoxidase (LPO) | PMOXA-b-PDMS [91] | Encapsulated (in polymersomes) | Oxidation of Amplex red using hydrogen peroxide |
| L-asparaginase | PMPC-b-PDPA and PEO-b-PBO [84], PEG-b-Pasp and P(Asp-AP) [64], PEG-b-PHPMA [78] | Encapsulated (in polymersomes, PICsomes) | Catalysis of L-asparagine to l-aspartic acid and ammonia |
| Lipase | PMOXA-b-PDMS-b-PMOXA [71] | Encapsulated (in polymersomes) | Catalysis of the hydrolysis of fats |
| Luciferase | PMOXA-b-PDMS [160] | Encapsulated (in polymersomes) | Bioluminescence |
| Melittin | PMOXA-b-PDMS [91,144,161] | Incorporated within membrane (polymersomes, GUVs) | Pore formation for cross-membrane diffusion |
| Methionine γ-lyase (MGL) | PEG-P(Asp) and PLys [63,89] | Encapsulated (in PICsomes) | Cancer therapy |
| Outer membrane protein F from E. coli (OmpF) | PMOXA-b-PDMS [39], PMOXA-b-PDMS-b-PMOXA [90,138] | Incorporated within membrane (GUVs, polymersomes) | Pore formation for cross-membrane diffusion |
| Penicillin acylase | PMOXA-b-PDMS-b-PMOXA [162] | Encapsulated (in polymersomes) | Production of antibiotic cephalexin |
| Rnase H | PEG-b-polypeptide (with single-stranded oligonucleotides) [80] | Encapsulated (in PICsomes) | Gene knockout therapy |
| β-galactosidase | PMOXA-b-PDMS [39] carbohydrate-b-PPG [54] | Encapsulated (in GUVs, capsosomes) | Catalysis of the hydrolysis of β-galactosides into monosaccharides |
| β-glucuronidase | PMOXA-b-PDMS [161] | Encapsulated (in polymersomes) | Cleavage of the glucuronide moiety from glucuronide-conjugates |
| Soluble guanylyl cyclase (sGC) | PMOXA-b-PDMS-b-PMOXA [159] | Encapsulated (in polymersomes) | Production of cyclic 3,5-guanosine monophosphate (cGMP) |
| Trypsin | PMPC-b-PDPA [93] | Encapsulated (in polymersomes) | Hydrolyzation of proteins |
| Tyrosinase | PMOXA-b-PDMS [163] | Encapsulated (in polymersomes) | Oxidation of L-DOPA |
| Urate oxidase (UOX) | PMOXA-b-PDMS-b-PMOXA [16] | Encapsulated (in polymersomes) | Production of hydrogen peroxide for a cascade reaction |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heuberger, L.; Korpidou, M.; Eggenberger, O.M.; Kyropoulou, M.; Palivan, C.G. Current Perspectives on Synthetic Compartments for Biomedical Applications. Int. J. Mol. Sci. 2022, 23, 5718. https://doi.org/10.3390/ijms23105718
Heuberger L, Korpidou M, Eggenberger OM, Kyropoulou M, Palivan CG. Current Perspectives on Synthetic Compartments for Biomedical Applications. International Journal of Molecular Sciences. 2022; 23(10):5718. https://doi.org/10.3390/ijms23105718
Chicago/Turabian StyleHeuberger, Lukas, Maria Korpidou, Olivia M. Eggenberger, Myrto Kyropoulou, and Cornelia G. Palivan. 2022. "Current Perspectives on Synthetic Compartments for Biomedical Applications" International Journal of Molecular Sciences 23, no. 10: 5718. https://doi.org/10.3390/ijms23105718
APA StyleHeuberger, L., Korpidou, M., Eggenberger, O. M., Kyropoulou, M., & Palivan, C. G. (2022). Current Perspectives on Synthetic Compartments for Biomedical Applications. International Journal of Molecular Sciences, 23(10), 5718. https://doi.org/10.3390/ijms23105718