
Characterization of the Cell Wall Component through Thermogravimetric Analysis and Its Relationship with an Expansin-like Protein in Deschampsia antarctica

 ,
, 
 , ,
, , 
Abstract
:1. Introduction
2. Result and Discussion
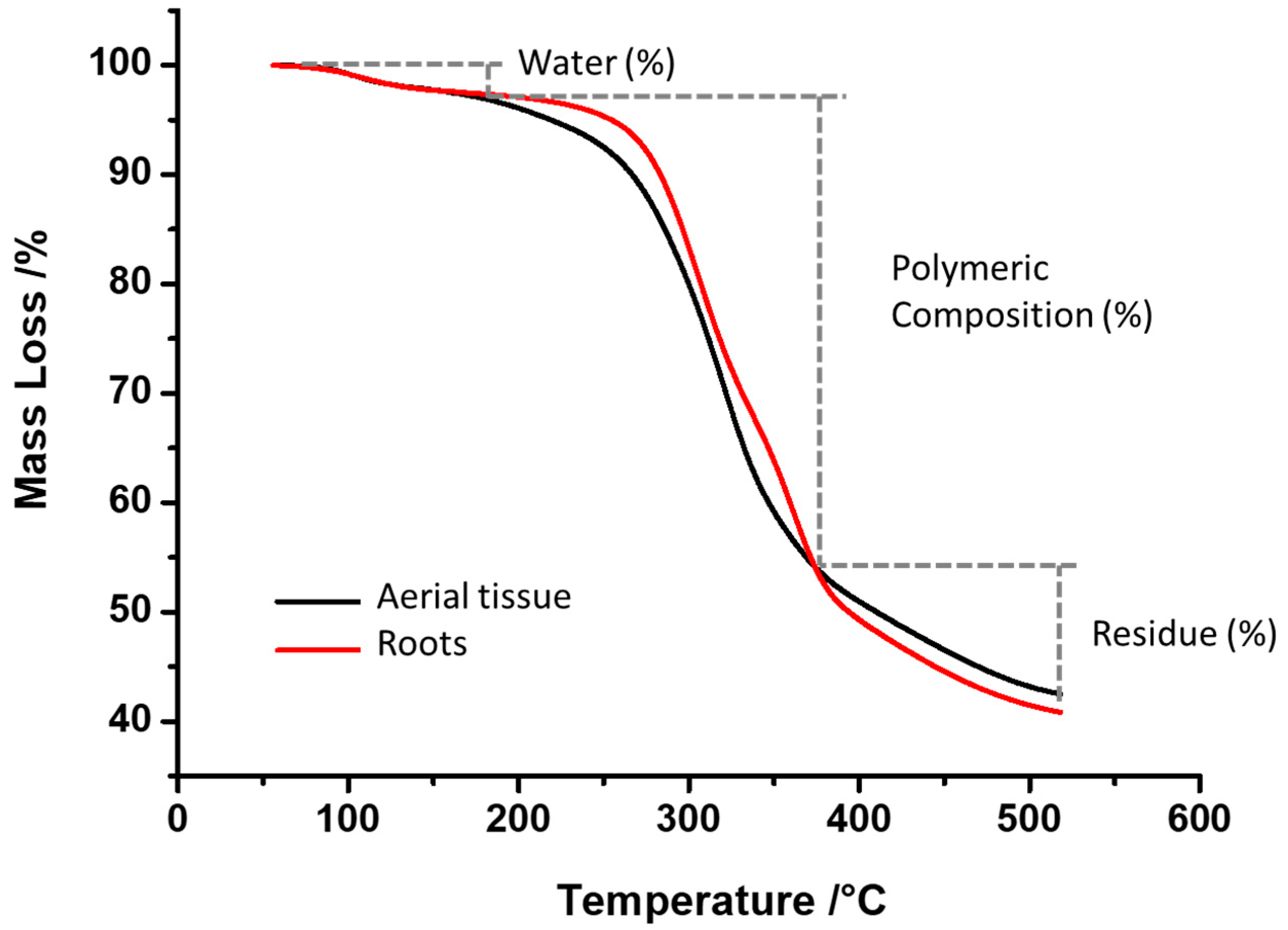
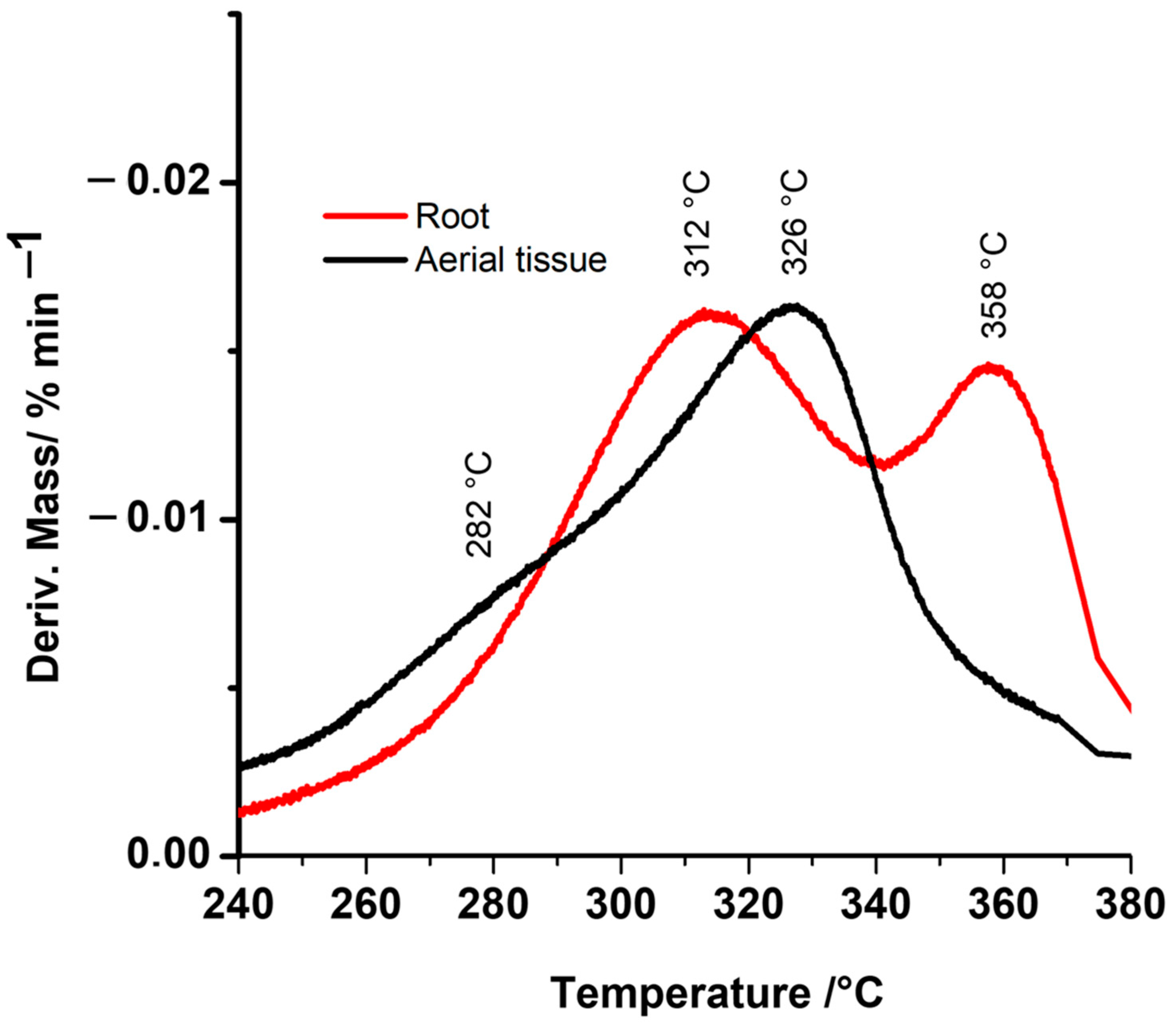
2.1. Thermogravimetric (TGA and DTG) Characterization
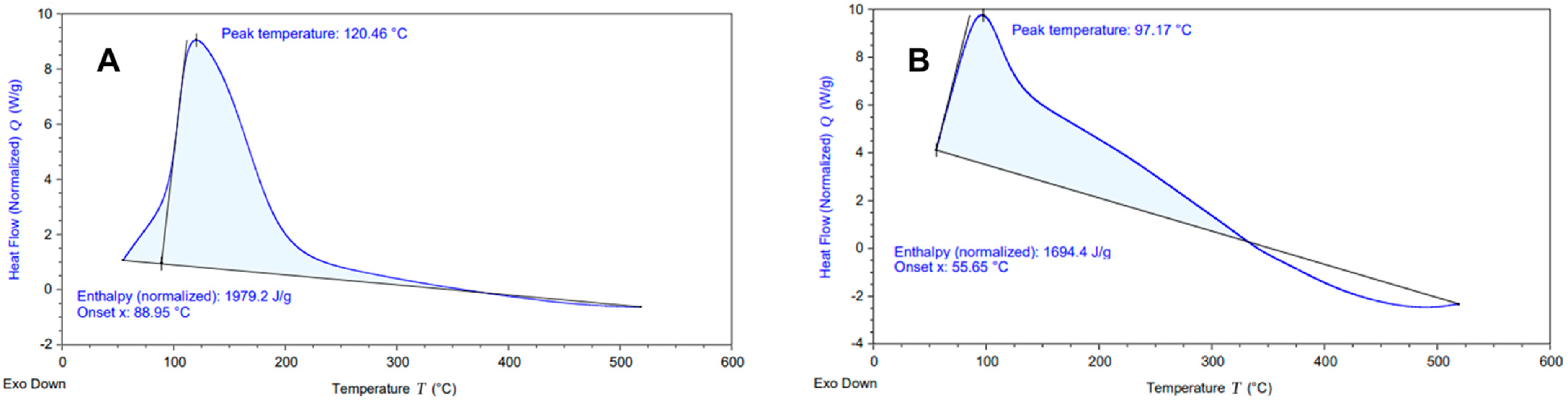
2.2. Differential Scanning Calorimetry (DSC) Analysis
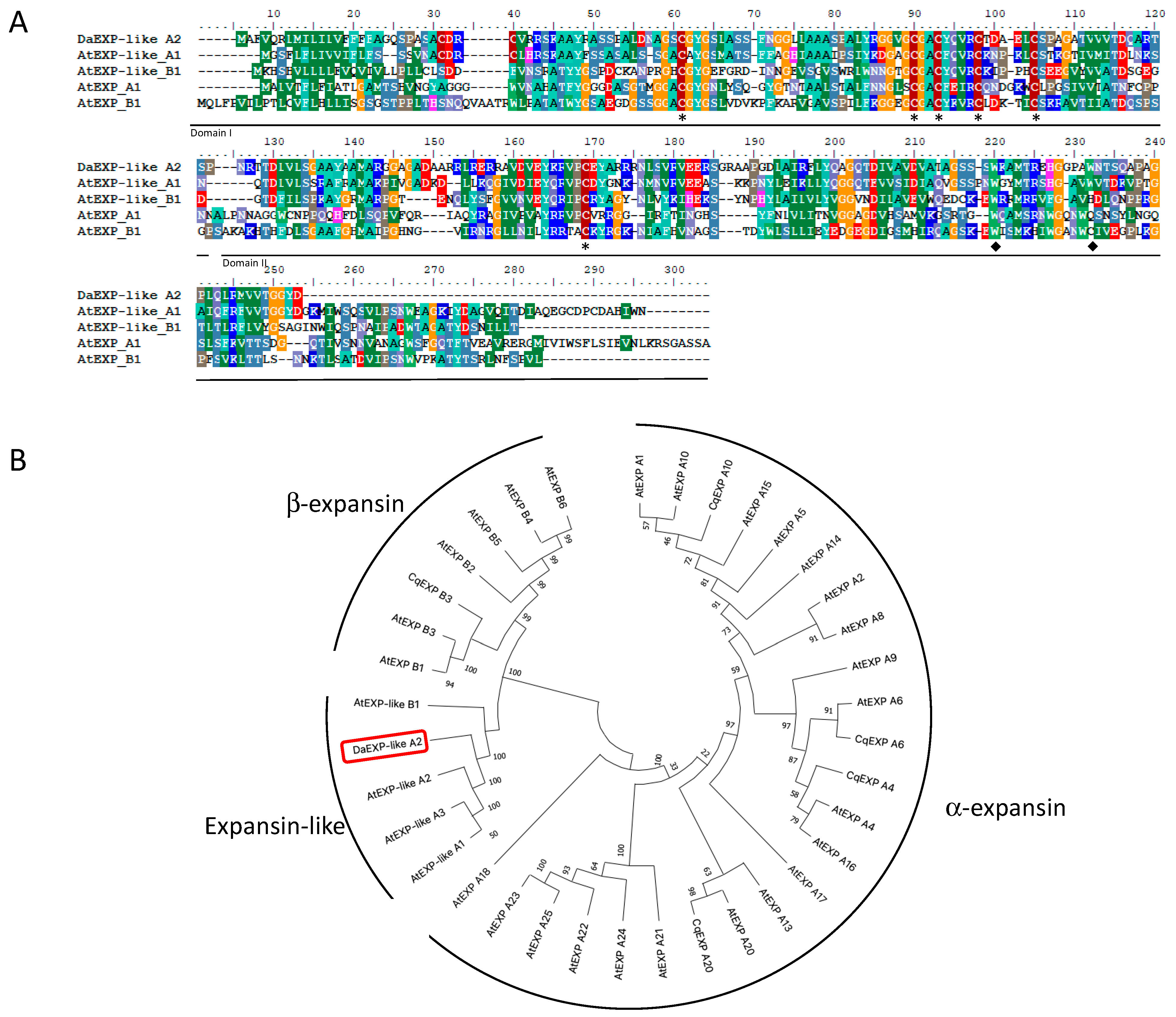
2.3. Isolation of DaEXLA2 and Phylogenetic Analysis
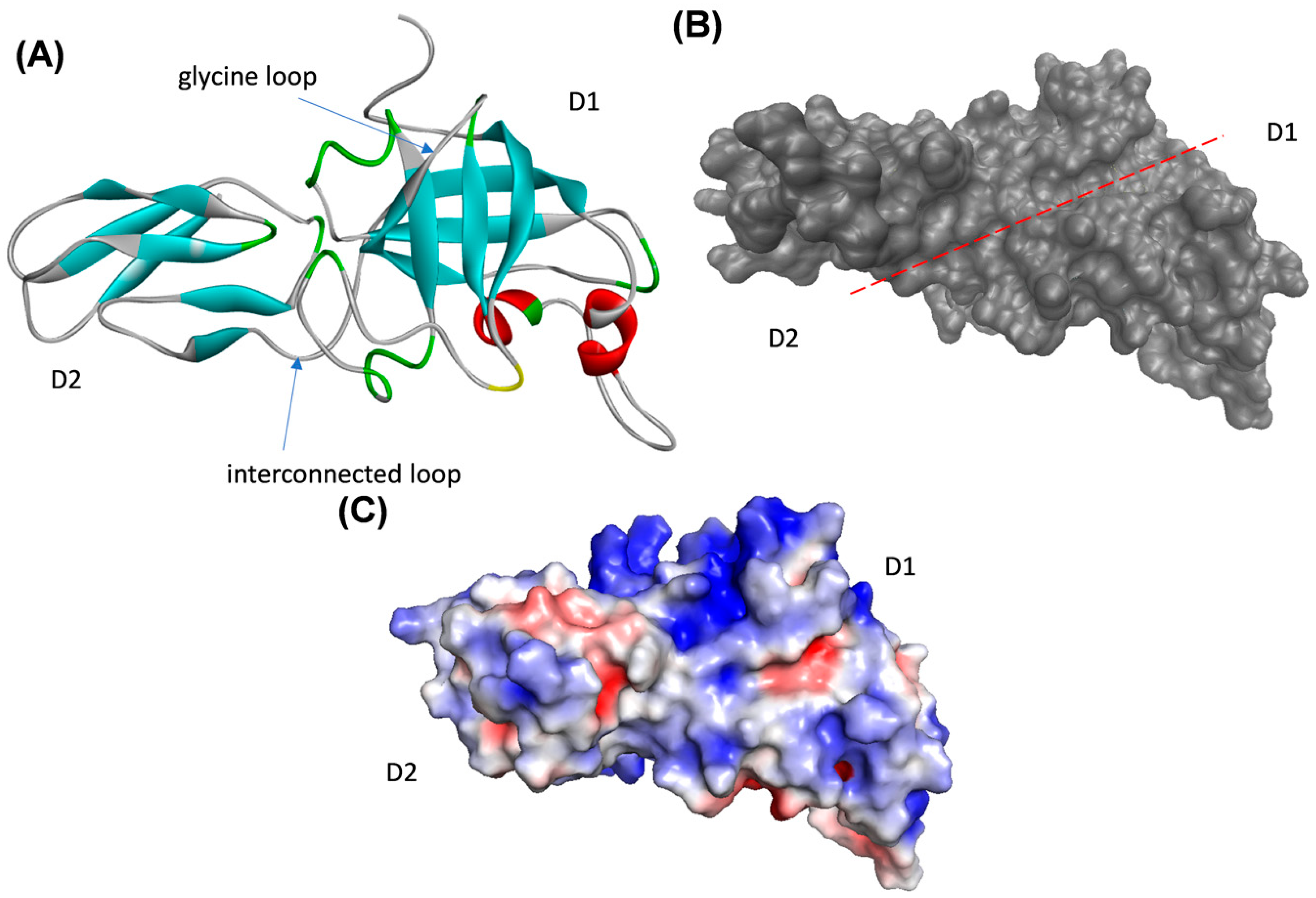
2.4. DaEXLA2 Protein Model Obtention
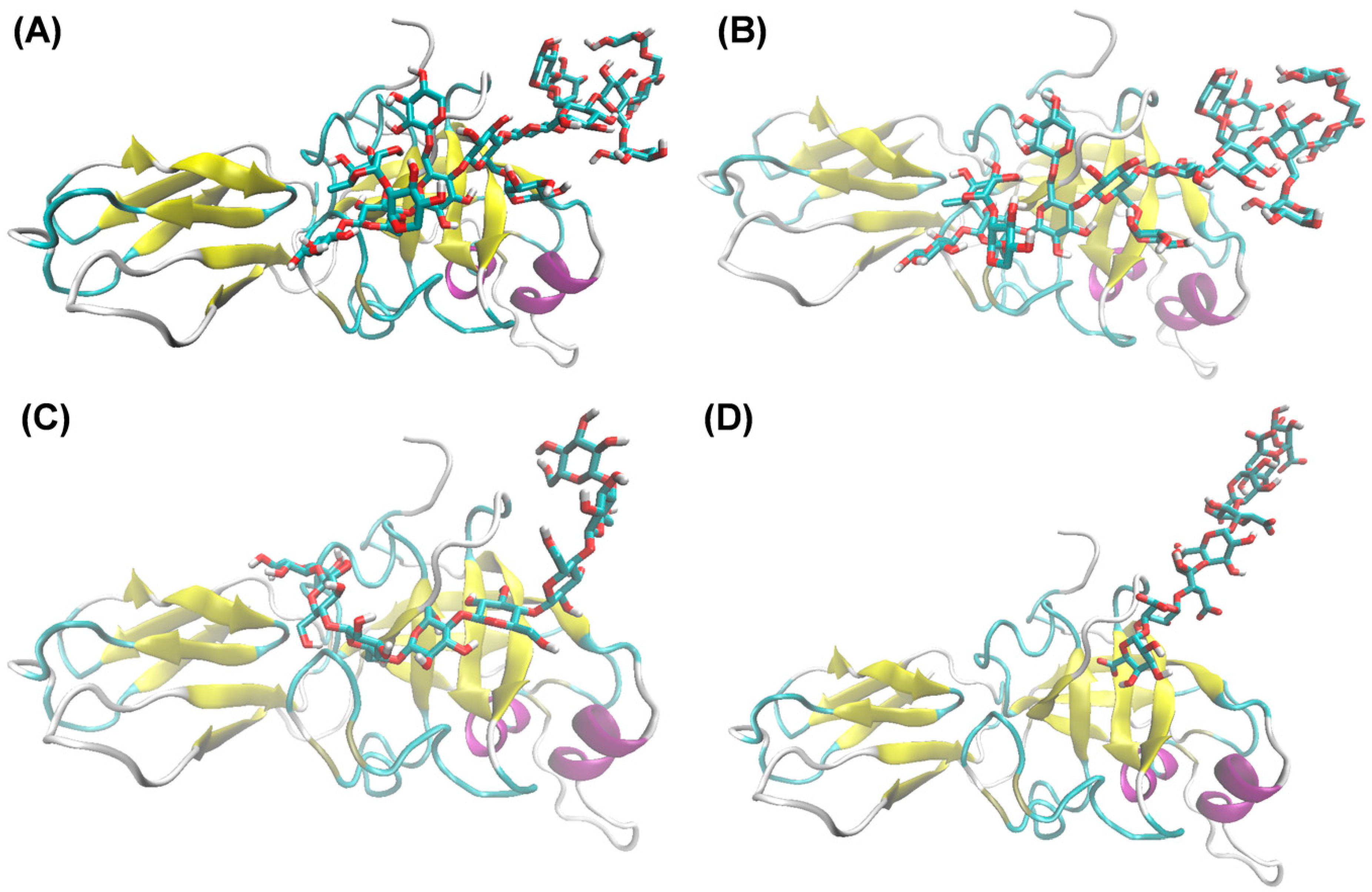
2.5. Evaluation of the Protein–Ligand Interaction Mode
3. Material and Methods
3.1. Plant Samples
3.2. Thermogravimetric Analysis (TGA)
3.3. Sequence Obtention and Analysis
3.4. DaEXLA2 Structural Model Construction
3.5. Determination of the DaEXLA2–Ligand Interaction by Molecular Dynamics Simulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alberdi, M.; Bravo, L.A.; Gutiérrez, A.; Gidekel, M.; Corcuera, L.J. Ecophysiology of Antarctic vascular plants. Physiol. Plantarum. 2002, 115, 479–486. [Google Scholar] [CrossRef]
- Gallardo-Cerda, J.; Levihuan, J.; Lavín, P.; Oses, R.; Atala, C.; Torres-Díaz, C.; Cuba-Díaz, M.; Barrera, A.; Molina-Montenegro, M.A. Antarctic rhizobacteria improve salt tolerance and physiological performance of the Antarctic vascular plants. Polar Biol. 2018, 41, 1973–1982. [Google Scholar] [CrossRef]
- Cannone, N.; Guglielmin, M.; Convey, P.; Worland, M.R.; Longo, S.F. Vascular plant changes in extreme environments: Effects of multiple drivers. Clim. Chang. 2016, 134, 651–665. [Google Scholar] [CrossRef] [Green Version]
- Barrientos-Díaz, L.; Gidekel, M.; Gutiérrez-Moraga, A. Characterization of rhizospheric bacteria isolated from Deschampsia antarctica Desv. World J. Microbiol. Biotechnol. 2008, 24, 2289–2296. [Google Scholar] [CrossRef]
- Gielwanowska, I.; Szczuka, E.; Bednara, J.; Górecki, R. Anatomical features and ultrastructure of Deschampsia antarctica (poaceae) leaves from different growing habitats. Ann. Bot. 2005, 96, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Parnikoza, I.; Kozeretska, I.; Kunakh, V. Vascular plants of the maritime antarctic: Origin and adaptation. Am. J. Plant Sci. 2011, 2, 381–395. [Google Scholar] [CrossRef] [Green Version]
- Rosa, L.H.; Vaz, A.B.M.; Caligiorne, R.B.; Campolina, S.; Rosa, C.A. Endophytic fungi associated with the Antarctic grass Deschampsia antarctica Desv. (Poaceae). Polar Biol. 2008, 32, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Bhatt, K.; Baumann, K. Shaping in plant cells. Curr. Opin. Plant Biol. 2001, 4, 540–549. [Google Scholar] [CrossRef]
- Smith, L.G. Cytoskeletal control of plant cell shape: Getting the fine points. Curr. Opin. Plant Biol. 2003, 6, 63–73. [Google Scholar] [CrossRef]
- Pilling, E.; Höfte, H. Feedback from the wall. Curr. Opin. Plant Biol. 2003, 6, 611–616. [Google Scholar] [CrossRef]
- Nardi, C.F.; Villareal, N.M.; Rossi, F.R.; Martinez, S.; Martinez, G.A.; Civello, P.M. Overexpression of the carbohydrate binding module of strawberry expansin2 in Arabidopsis thaliana modifies plant growth and cell wall metabolism. Plant Mol. Biol. 2015, 88, 101–117. [Google Scholar] [CrossRef]
- Castro, R.I.; Morales-Quintana, L. Study of the cell wall components produced during different ripening stages through thermogravimetric analysis. Cellulose 2019, 26, 3009–3020. [Google Scholar] [CrossRef]
- Castro, R.I.; Gonzalez-Feliu, A.; Muñoz-Vera, M.; Valenzuela-Riffo, F.; Parra-Palma, C.; Morales-Quintana, L. Effect of the exogenous auxin treatment in strawberry fruit over cell wall polymers. Int. J. Mol. Sci. 2021, 22, 6294. [Google Scholar] [CrossRef]
- Castro, R.I.; Gonzalez-Feliu, A.; Valenzuela-Riffo, F.; Parra-Palma, C.; Morales-Quintana, L. Changes in the cell wall components produced by exogenous abscisic acid treatment in strawberry fruit. Cellulose 2021, 28, 1555–1570. [Google Scholar] [CrossRef]
- Zenoni, S.; Reale, L.; Tornielli, G.B.; Lanfaloni, L.; Porceddu, A.; Ferrarini, A.; Moretti, C.; Zamboni, A.; Speghini, A.; Ferranti, F.; et al. Downregulation of the Petunia hybrida α-expansin gene PhEXP1 reduces the amount of crystalline cellulose in cell walls and leads to phenotypic changes in petal limbs. Plant Cell 2004, 16, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, L.; Hao, W.; Zhang, L.; Liu, Y.; Chen, L. Expression of Two α-Type Expansins from Ammopiptanthus nanus in Arabidopsis thaliana Enhance Tolerance to Cold and Drought Stresses. Int. J. Mol. Sci. 2019, 20, 5255. [Google Scholar] [CrossRef] [Green Version]
- Morales-Quintana, L.; Barrera, A.; Hereme, R.; Jara, K.; Rivera-Mora, C.; Valenzuela-Riffo, F.; Gundel, P.E.; Pollmann, S.; Ramos, P. Molecular and structural characterization of expansins modulated by fungal endophytes in the Antarctic Colobanthus quitensis (Kunth) Bartl. Exposed to drought stress. Plant Physiol. Biochem. 2021, 168, 465–476. [Google Scholar] [CrossRef]
- McQueen-Mason, S.; Cosgrove, D.J. Disruption of hydrogen bonding between plant cell wall polymers by proteins that induce wall extension. Proc. Natl. Acad. Sci. USA 1994, 91, 6574–6578. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Choi, D.; Kende, H. Expansins: Ever-expanding numbers and functions. Curr. Opin. Plant Biol. 2001, 4, 527–532. [Google Scholar] [CrossRef]
- Saloheimo, M.; Paloheimo, M.; Hakola, S.; Pere, J.; Swanson, B.; Nyyssönen, E.; Bhatia, A.; Ward, M.; Penttilä, M. Swollenin, a Trichoderma reesei protein with sequence similarity to the plant expansins, exhibits disruption activity on cellulosic materials. Eur. J. Biochem. 2002, 269, 4202–4211. [Google Scholar] [CrossRef]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, D.J. Plant expansins: Diversity and interactions with plant cell walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Bunterngsook, B.; Eurwilaichitr, L.; Thamchaipenet, A.; Champreda, V. Binding characteristics and synergistic effects of bacterial expansins on cellulosic and hemicellulosic substrates. Bioresour. Technol. 2015, 176, 129–135. [Google Scholar] [CrossRef]
- Cosgrove, D.J. New genes and new biological roles for expansins. Curr. Opin. Plant Biol. 2000, 3, 73–78. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.; Shi, Y.; Xu, J.; Huang, B. Transgenic tobacco plants overexpressing a grass PpEXP1 gene exhibit enhanced tolerance to heat stress. PLoS ONE 2014, 9, e100792. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Ding, A.; Marowa, P.; Kong, Y. Genome-wide identification of the expansin gene family in tobacco (Nicotiana tabacum). Mol. Genet. Genom. 2016, 291, 1891–1907. [Google Scholar] [CrossRef]
- Kuluev, B.; Avalbaev, A.; Mikhaylova, E.; Nikonorov, Y.; Berezhneva, Z.; Chemeris, A. Expression profiles and hormonal regulation of tobacco expansin genes and their involvement in abiotic stress response. J. Plant. Physiol. 2016, 206, 1–12. [Google Scholar] [CrossRef]
- Tan, J.; Wang, M.; Shi, Z.; Miao, X. OsEXPA10 mediates the balance between growth and resistance to biotic stress in rice. Plant Cell Rep. 2018, 37, 993–1002. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Kong, C.; Yang, Y.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLoS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef] [Green Version]
- Muthusamy, M.; Kim, J.Y.; Yoon, E.K.; Kim, J.A.; Lee, S.I. BrEXLB1, a Brassica rapa Expansin-Like B1 Gene is Associated with Root Development, Drought Stress Response, and Seed Germination. Genes 2020, 11, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from enzymatic degradation of cellulose and hemicellulose to fermentable sugars—A review. Biomass Bioenerg. 2020, 134, 10548. [Google Scholar] [CrossRef]
- Grønli, M.G.; Várhegyi, G.; Di Blasi, C. Thermogravimetric Analysis and Devolatilization Kinetics of Wood. Ind. Eng. Chem. Res. 2002, 41, 4201–4208. [Google Scholar] [CrossRef]
- Johnson, K.G.; Silva, M.C.; Mackenzie, C.R.; Schneider, H.; Fontana, J.D. Microbial degradation of hemicellulosic materials. Appl. Biochem. Biotechnol. 1989, 20–21, 245–258. [Google Scholar] [CrossRef]
- Shen, D.K.; Gu, S.; Bridgwater, A.V. Study on the pyrolytic behaviour of xylan-based hemicellulose using TG–FTIR and Py–GC–FTIR. J. Anal. Appl. Pyrolysis. 2010, 87, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Tao, P.; Zhang, N.; Nie, S. Preparation and thermal stability evaluation of cellulose nanofibrils from bagasse pulp with differing hemicelluloses contents. Carbohydr. Polym. 2020, 245, 116463. [Google Scholar] [CrossRef]
- Zhou, X.; Li, W.; Mabon, R.; Broadbelt, L.J. A Critical Review on Hemicellulose Pyrolysis. Energy Technol. 2016, 5, 52–79. [Google Scholar] [CrossRef] [Green Version]
- Nam, S.; Hillyer, M.B.; Condon, B.D. Method for identifying the triple transition (glass transition-dehydration-crystallization) of amorphous cellulose in cotton. Carbohydr. Polym. 2020, 228, 115374. [Google Scholar] [CrossRef]
- Bigger, S.W.; Scheirs, J.; Camino, G. An investigation of the kinetics of cellulose degradation under non-isothermal conditions. Polym. Degrad. Stab. 1998, 62, 33–40. [Google Scholar] [CrossRef]
- Wu, Y.; Dollimore, D. Kinetic studies of thermal degradation of natural cellulosic materials. Thermochim. Acta 1998, 324, 49–57. [Google Scholar] [CrossRef]
- Hattori, K.; Arai, A. Preparation and Hydrolysis of Water-Stable Amorphous Cellulose. ACS Sustain. Chem. Eng. 2016, 4, 1180–1186. [Google Scholar] [CrossRef]
- Jones, L.; McQueen-Mason, S. A role for expansins in dehydration and rehydration of the resurrection plant Craterostigma Plantagineum. FEBS Lett. 2004, 559, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Mateluna, P.; Valenzuela-Riffo, F.; Morales-Quintana, L.; Herrera, R.; Ramos, P. Transcriptional and computational study of expansins differentially expressed in the response to inclination in radiata pine. Plant Physiol. Biochem. 2017, 115, 12–24. [Google Scholar] [CrossRef]
- Gaete-Eastman, C.; Morales-Quintana, L.; Herrera, R.; Moya-León, M.A. In silico Analysis of structure and binding site feature of expansin protein from mountain papaya fruit (VpEXPA2), through molecular modeling, docking and dynamics simulation studies. J. Mol. Model. 2015, 21, 115. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Ramos, P.; Morales-Quintana, L. Computational study of FaEXPA1, a strawberry alpha expansin protein, through molecular modeling and molecular dynamics simulation studies. Comput. Biol. Chem. 2018, 76, 79–86. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Gaete-Eastman, C.; Stappung, Y.; Lizana, R.; Herrera, R.; Moya-León, M.A.; Morales-Quintana, L. Comparative in silico study of the differences in the structure and ligand interaction properties of three alpha-expansin proteins from Fragaria chiloensis fruit. J. Biomol. Struct. Dynam. 2019, 37, 3245–3258. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Parra-Palma, C.; Ramos, P.; Morales-Quintana, L. Molecular and structural insights into FaEXPA5, an alpha-expansin protein related with cell wall disassembly during ripening of strawberry fruit. Plant Physiol. Biochem. 2020, 154, 581–589. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Morales-Quintana, L. Study of the structure and binding site features of FaEXPA2, an α-expansin protein involved in strawberry fruit softening. Comput. Biol. Chem. 2020, 87, 107279. [Google Scholar] [CrossRef]
- Georgelis, N.; Tabuchi, A.; Nikolaidis, N.; Cosgrove, D.J. Structure-Function Analysis of the Bacterial Expansin EXLX1. J. Biol. Chem. 2011, 286, 16814–16823. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Valdebenito, D.; Bravo Ramirez, L.A.; Arce–Johnson, P.; Gutiérrez-Moraga, A. Salt tolerance traits in Deschampsia antarctica Desv. Antarctic Sci. 2016, 28, 462–472. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95⁄98⁄NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Šali, A.; Blundell, T.L. Comparative protein modeling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Morales-Quintana, L.; Fuentes, L.; Gaete-Eastman, C.; Herrera, R.; Moya-León, M.A. Structural characterization and substrate specificity of VpAAT1 protein related to ester biosynthesis in mountain papaya fruit. J. Mol. Graph. Model. 2011, 29, 635–642. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins. Proteins 1993, 17, 355–362. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in threedimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [Green Version]
- DeLano, W. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDockVina, Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Vergara-Jaque, A.; Comer, J.; Monsalve, L.; González-Nilo, F.D.; Sandoval, C. Computationally efficient methodology for atomic-level characterization of dendrimer−drug complexes: A comparison of amine- and acetyl-terminated PAMAM. J. Phys. Chem. B 2013, 117, 6801–6813. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Temperature °C | Average Percentage of Weight Loss (%) | |
|---|---|---|---|
| Aerial Tissue | Root Tissue | ||
| 1st degradation stage (Water loss) | 50 to 180 | 3.07 | 2.75 |
| 2nd degradation stage (polymeric composition) | 180 to 380 | 43.73 | 45.96 |
| Structure | Verify3D (Score) | ProSA (Z-Score) | PROCHECK | |||
|---|---|---|---|---|---|---|
| Core (%) a | Allow (%) b | Gener (%) c | Disall (%) d | |||
| DaEXLA2 | 85.2% | −4.09 | 80.2% | 18.9% | 0.8% | 0.0% |
| Protein | Ligand Name | Affinity Energy (kcal mol−1) |
|---|---|---|
| DaEXLA2 | Cellodextrin 8-mer | −6.9 b ± 0.35 |
| XXXGXXXG | −8.2 a ± 0.11 | |
| XXFGXXFG | −6.1 c ± 0.43 | |
| GAX | −5.4 d ± 0.11 |
| Ligand | ΔHvdWMM (kcal mol−1) | ΔHelecMM (kcal mol−1) | ΔGsol–pol (kcal mol−1) | ΔGsol–npol (kcal mol−1) | ΔGbind (kcal mol−1) |
|---|---|---|---|---|---|
| Cellodextrin 8-mer | −60.2 | 22.1 | 0.0 | −41.0 | –70.6 ± 0.7 |
| XXXGXXXG | −65.7 | 27.3 | 0.0 | −31.4 | –69.8 ± 1.5 |
| XXFGXXFG | −67.3 | 31.1 | 0.3 | −48.2 | –84.1 ± 0.8 |
| GAX | −39.3 | 30.5 | 0.8 | −41.2 | –49.2 ± 3.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Quintana, L.; Tapia-Valdebenito, D.; Castro, R.I.; Rabert, C.; Larama, G.; Gutiérrez, A.; Ramos, P. Characterization of the Cell Wall Component through Thermogravimetric Analysis and Its Relationship with an Expansin-like Protein in Deschampsia antarctica. Int. J. Mol. Sci. 2022, 23, 5741. https://doi.org/10.3390/ijms23105741
Morales-Quintana L, Tapia-Valdebenito D, Castro RI, Rabert C, Larama G, Gutiérrez A, Ramos P. Characterization of the Cell Wall Component through Thermogravimetric Analysis and Its Relationship with an Expansin-like Protein in Deschampsia antarctica. International Journal of Molecular Sciences. 2022; 23(10):5741. https://doi.org/10.3390/ijms23105741
Chicago/Turabian StyleMorales-Quintana, Luis, Daisy Tapia-Valdebenito, Ricardo I. Castro, Claudia Rabert, Giovanni Larama, Ana Gutiérrez, and Patricio Ramos. 2022. "Characterization of the Cell Wall Component through Thermogravimetric Analysis and Its Relationship with an Expansin-like Protein in Deschampsia antarctica" International Journal of Molecular Sciences 23, no. 10: 5741. https://doi.org/10.3390/ijms23105741
APA StyleMorales-Quintana, L., Tapia-Valdebenito, D., Castro, R. I., Rabert, C., Larama, G., Gutiérrez, A., & Ramos, P. (2022). Characterization of the Cell Wall Component through Thermogravimetric Analysis and Its Relationship with an Expansin-like Protein in Deschampsia antarctica. International Journal of Molecular Sciences, 23(10), 5741. https://doi.org/10.3390/ijms23105741