
Mutation of Protoporphyrinogen IX Oxidase Gene Causes Spotted and Rolled Leaf and Its Overexpression Generates Herbicide Resistance in Rice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
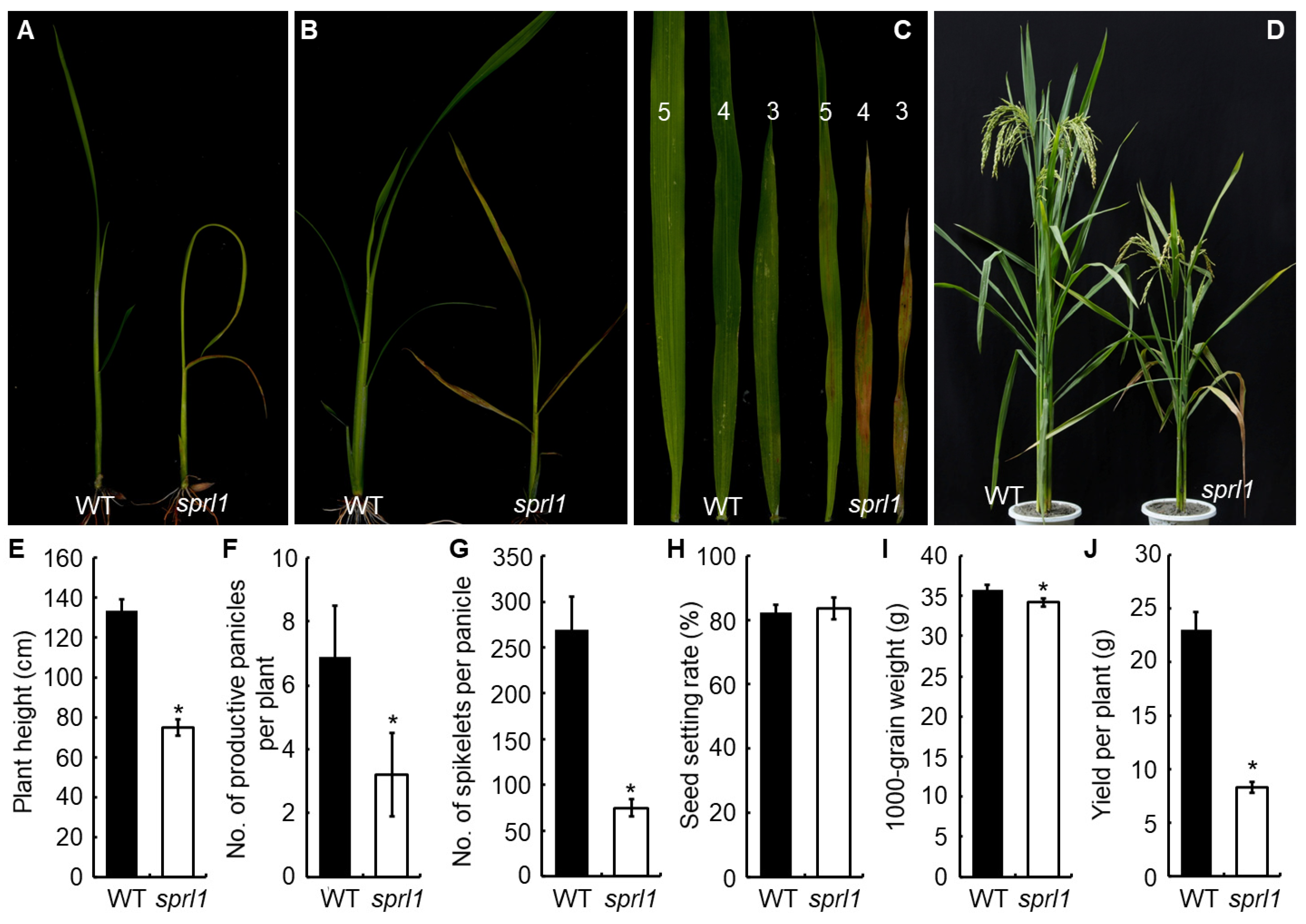
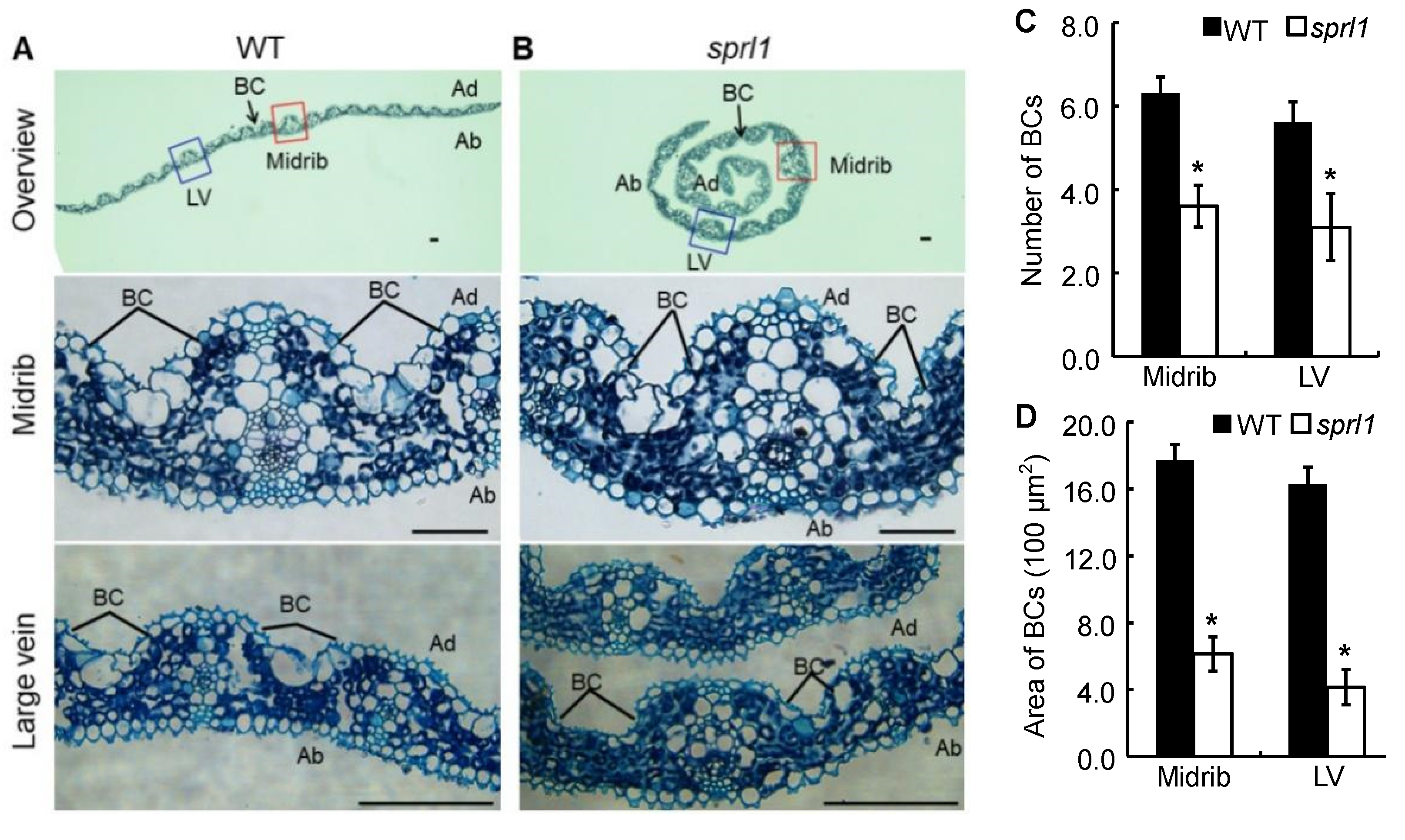
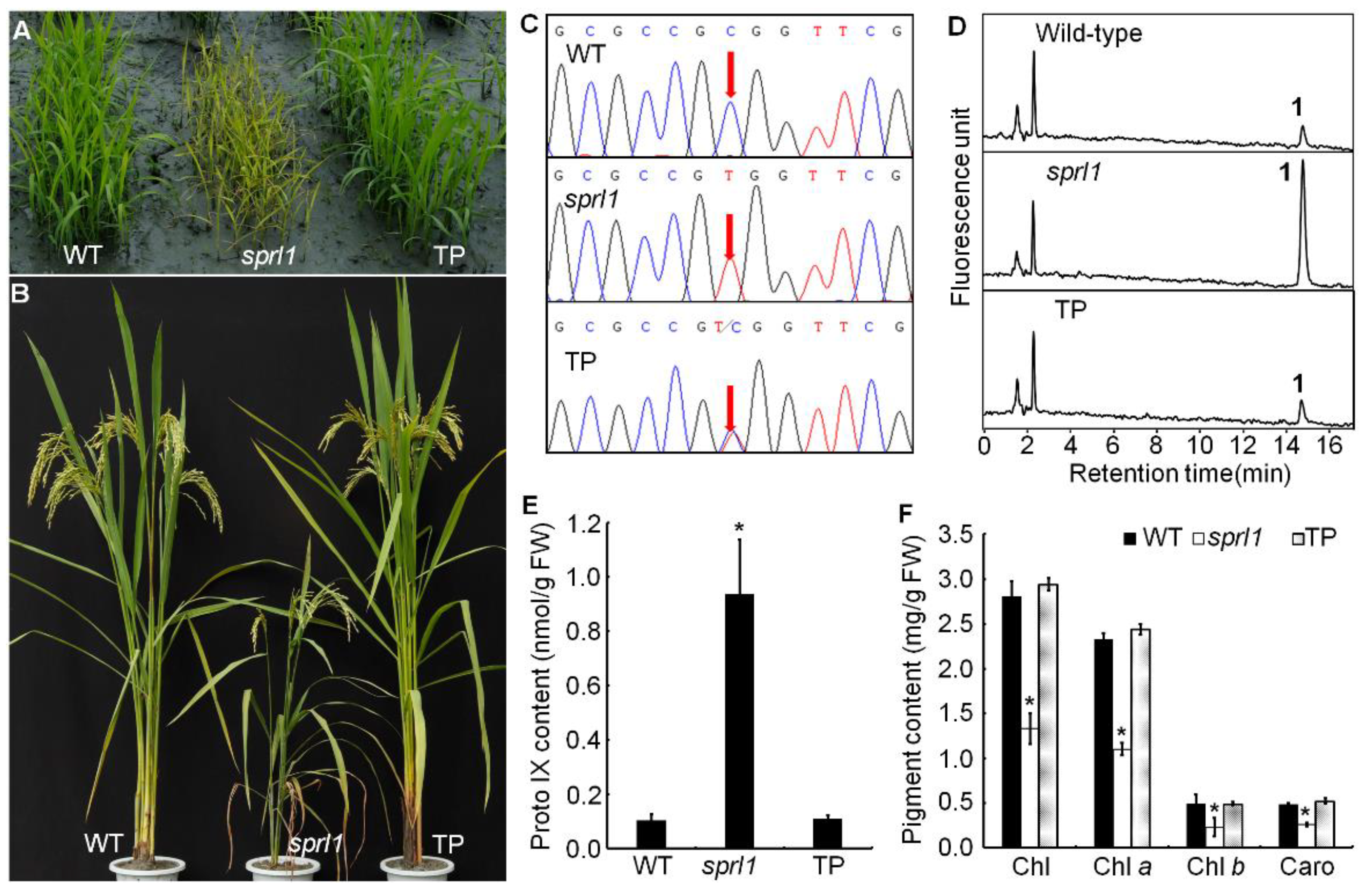
2.1. Phenotypic Characterization of the sprl1 Mutant
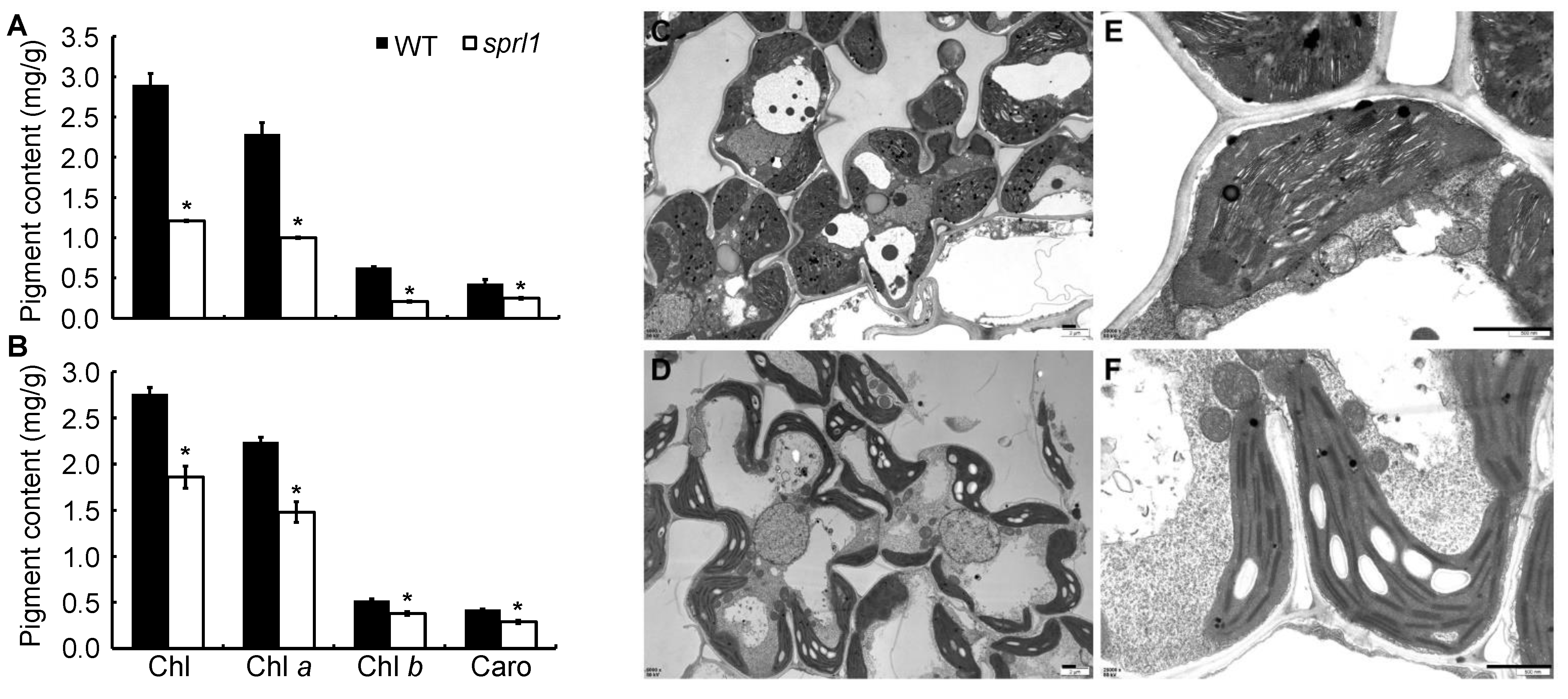
2.2. Physiological Characterization of the sprl1 Mutant
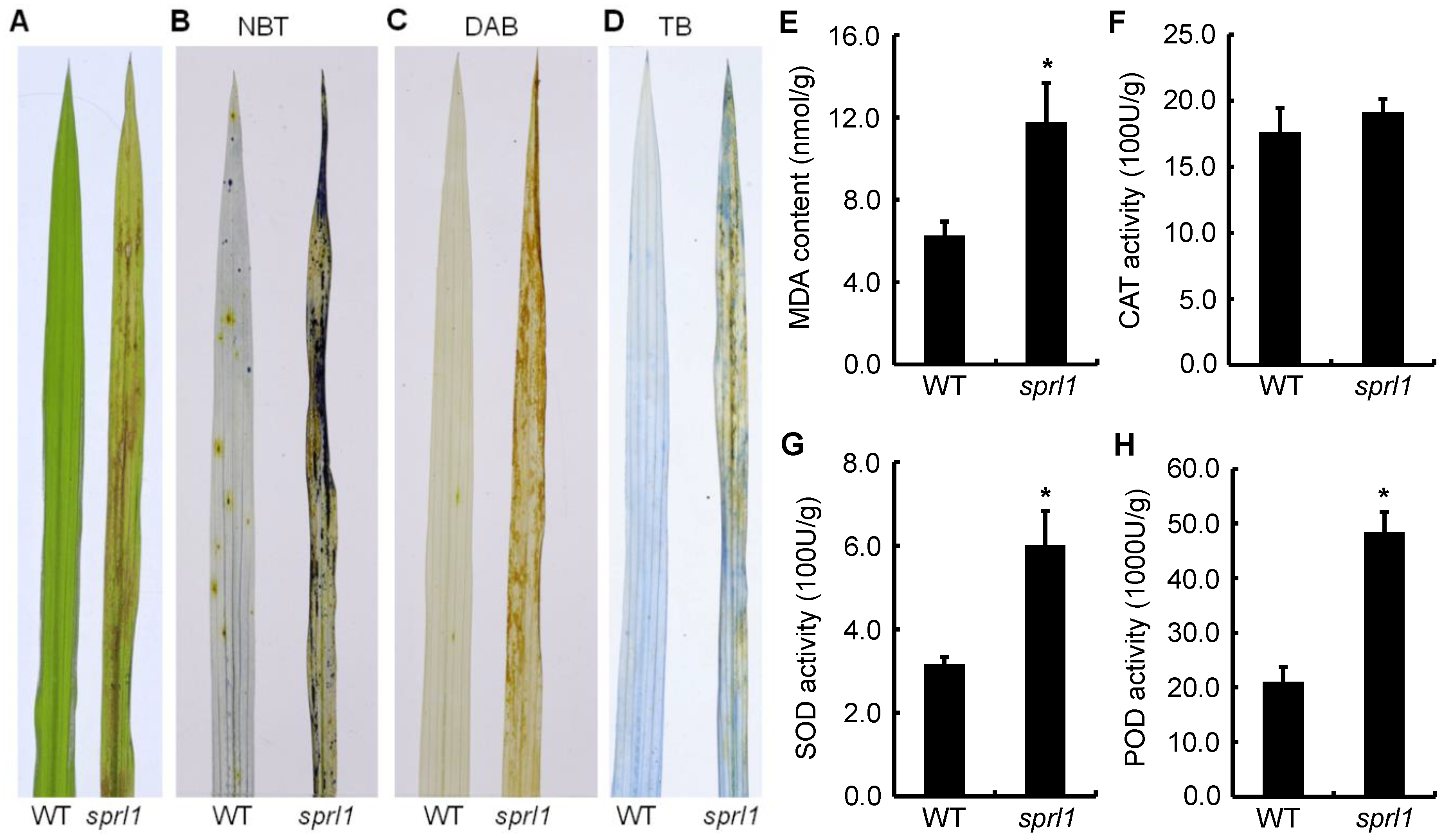
2.3. ROS Accumulation in the sprl1 Mutant
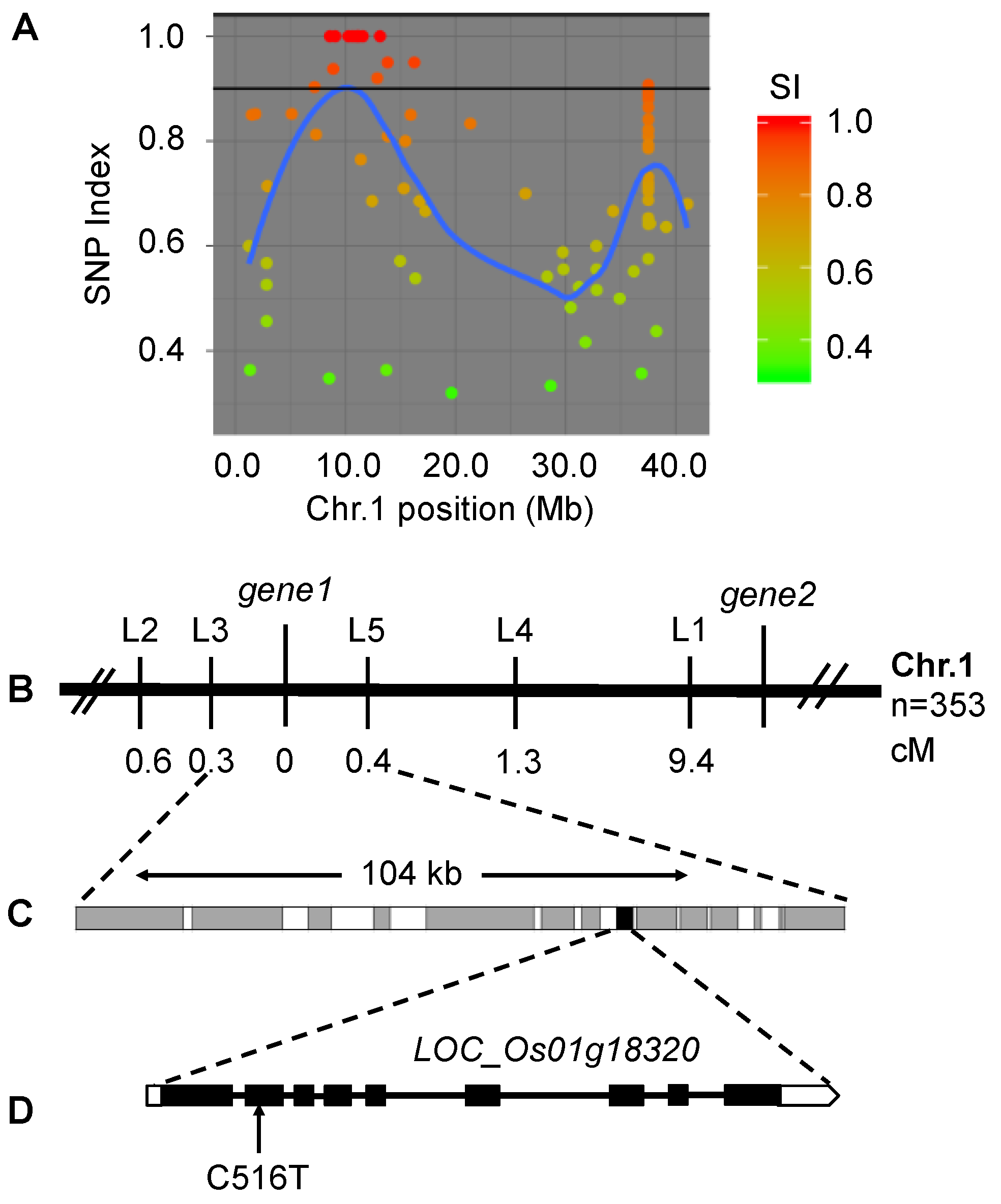
2.4. The sprl1 Locus Mapped to a Putative Gene Encoding Protogen IX Oxidase
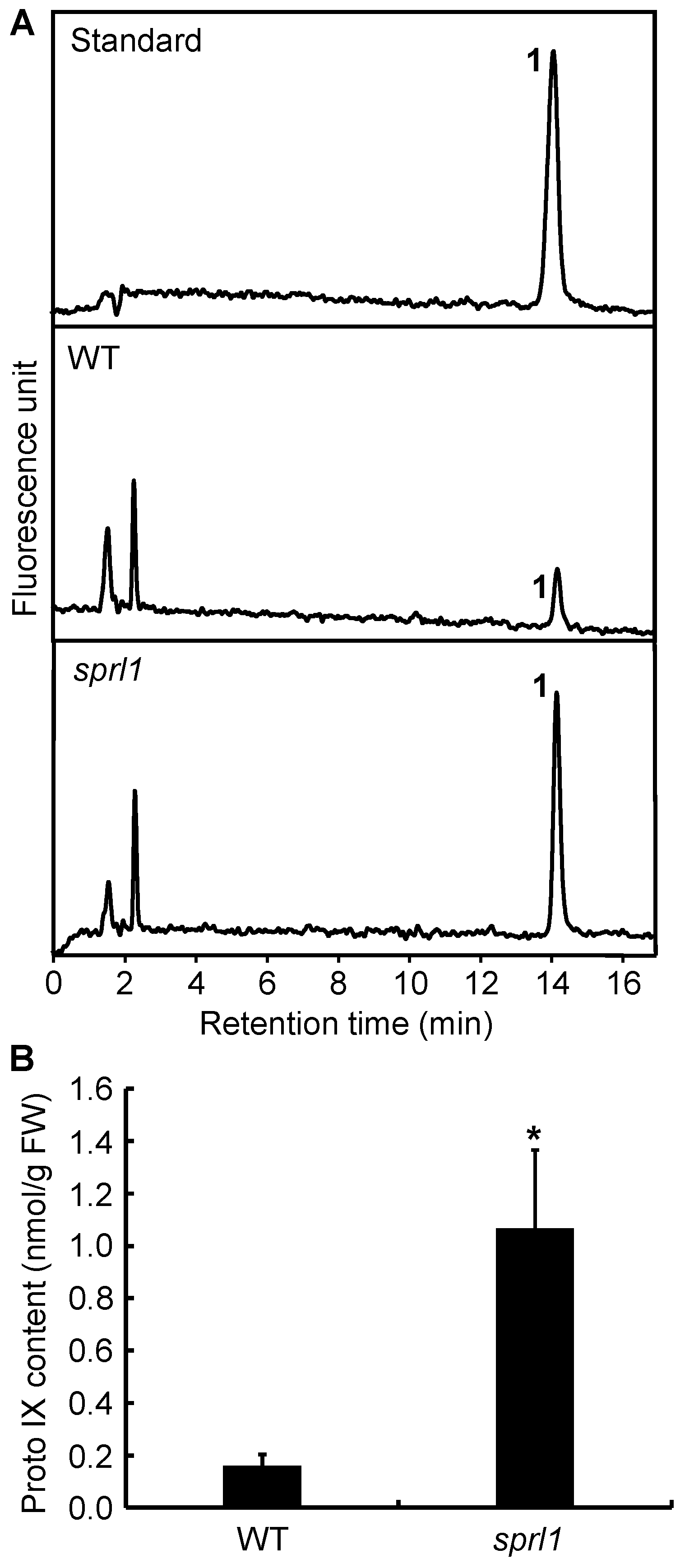
2.5. Excessive Accumulation of Proto IX in sprl1 Mutant
2.6. OsPPO1 Gene Rescued the Mutant Phenotype of sprl1
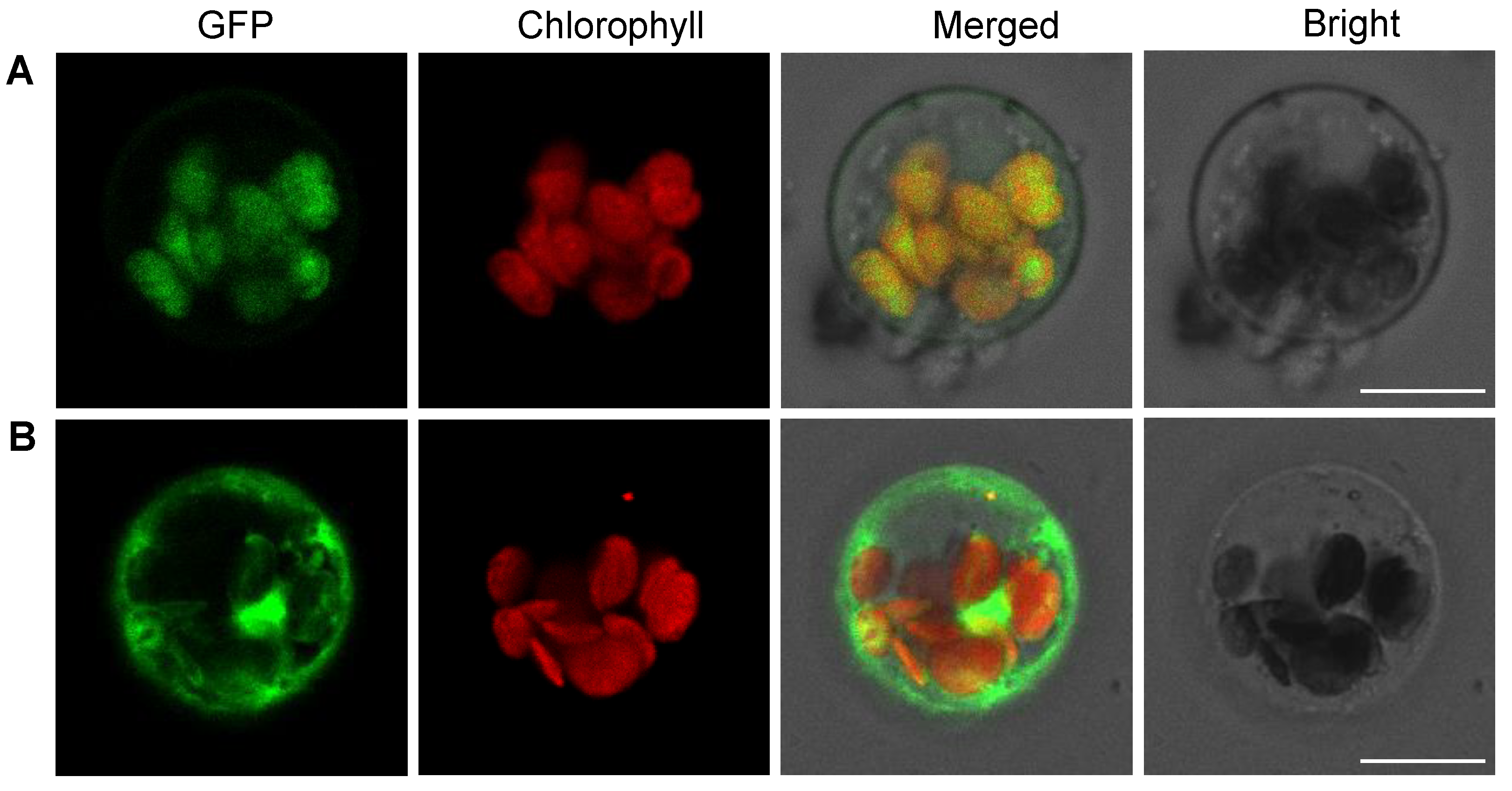
2.7. OsPPO1 Gets Localized to Chloroplast
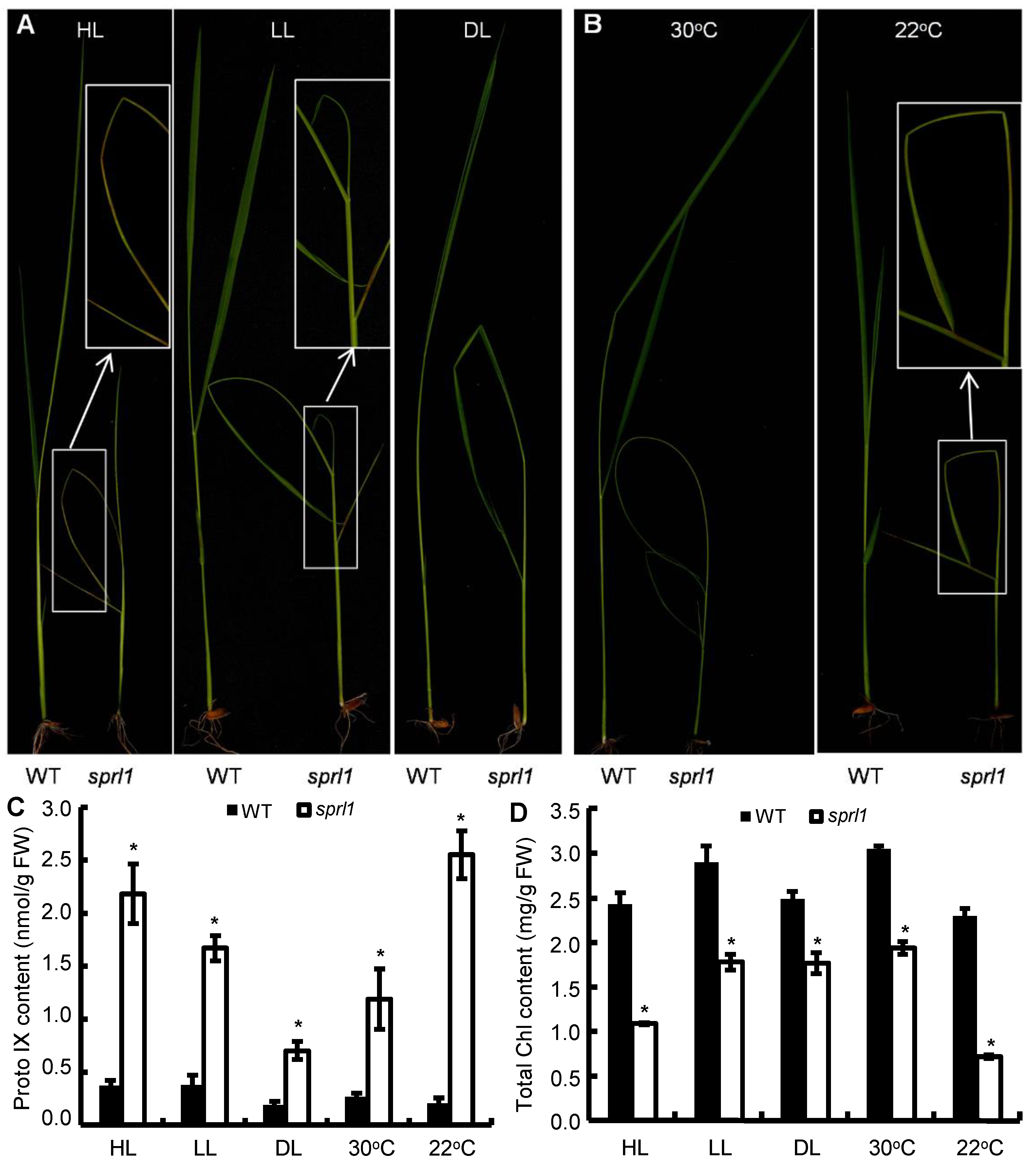
2.8. Light and Temperature Sensitivity of sprl1 Mutant
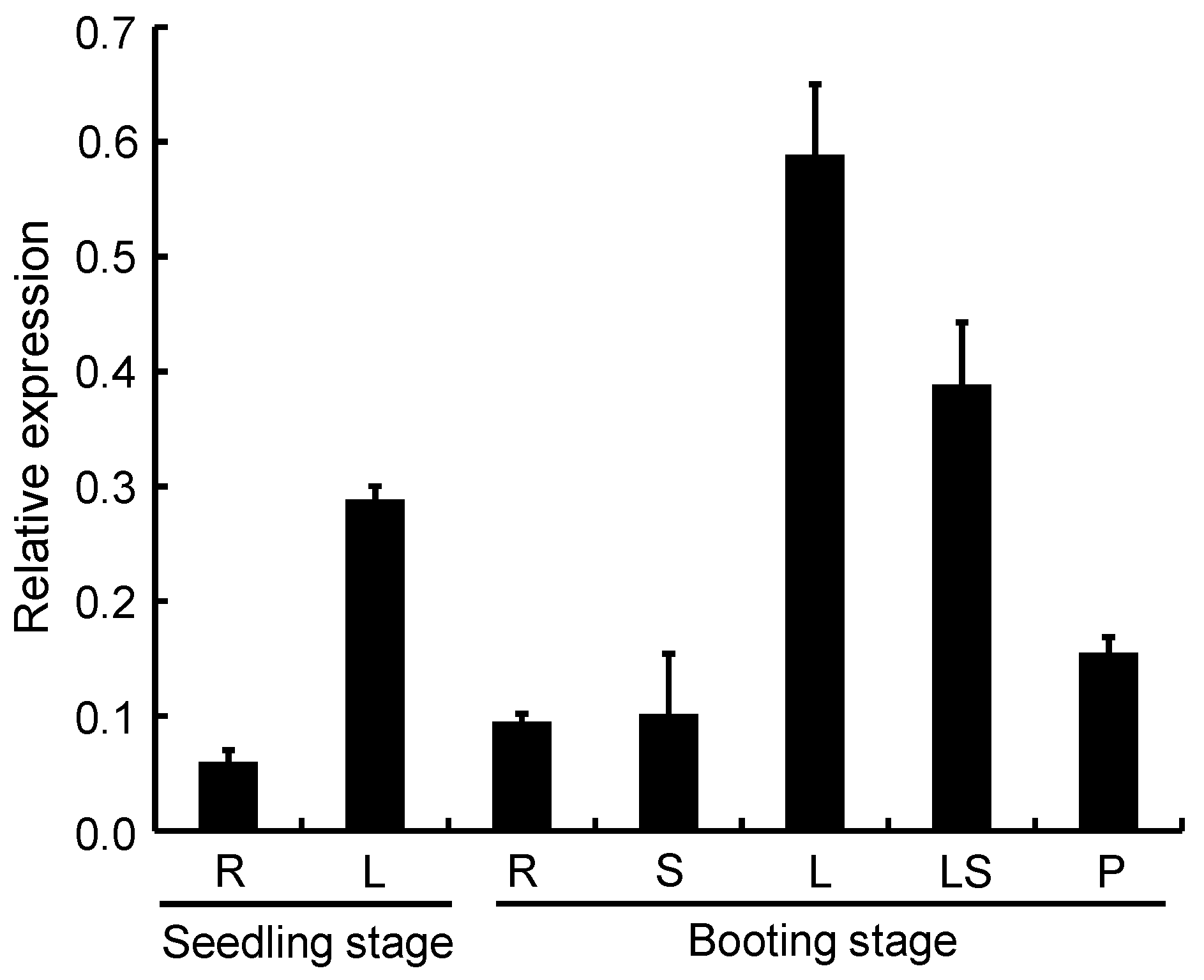
2.9. Expression Pattern Analysis of OsPPO1
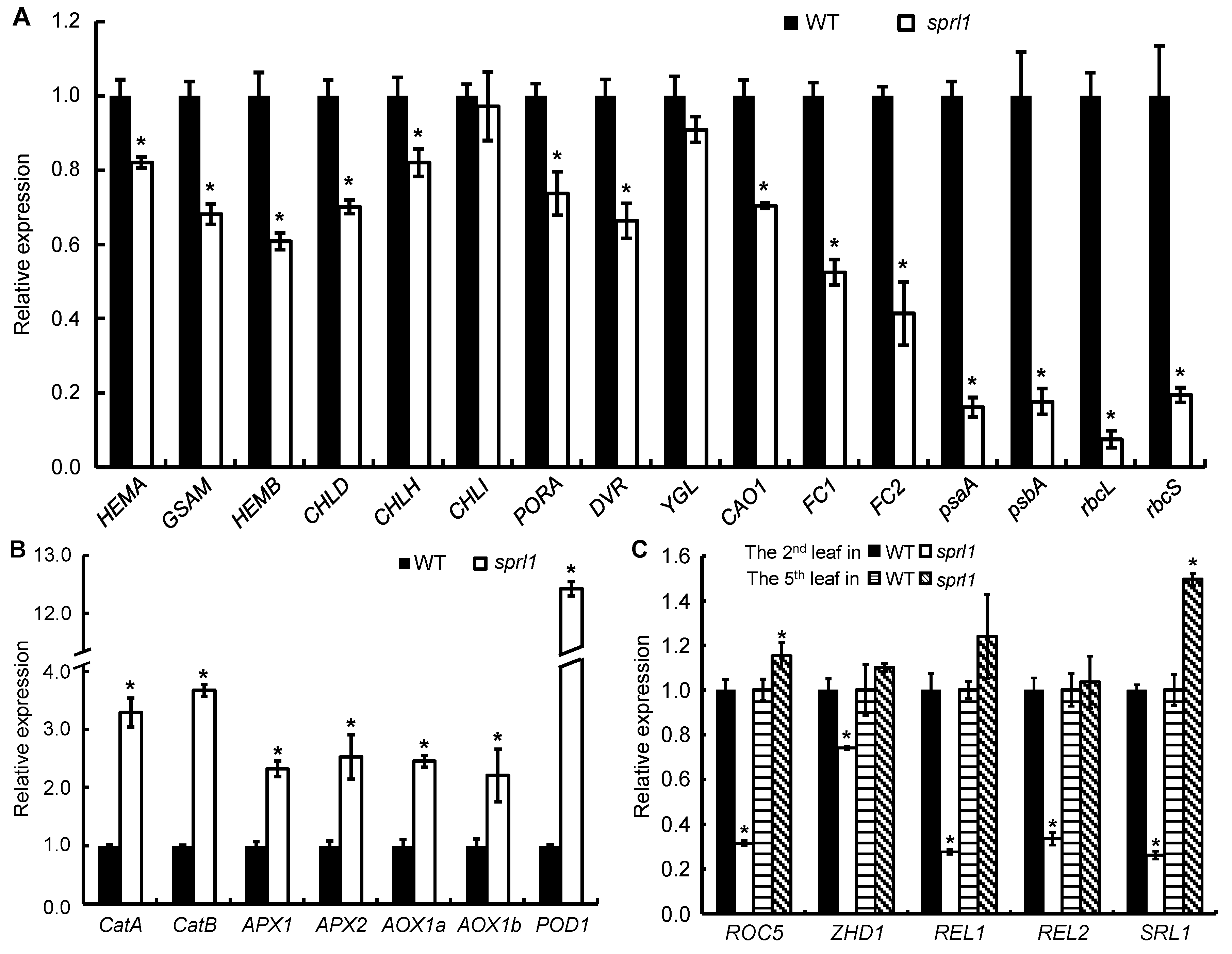
2.10. Expression Analysis of the Genes Associated with Tetrapyrrole Synthesis, Photosynthesis, ROS Accumulation, and Rolled Leaf
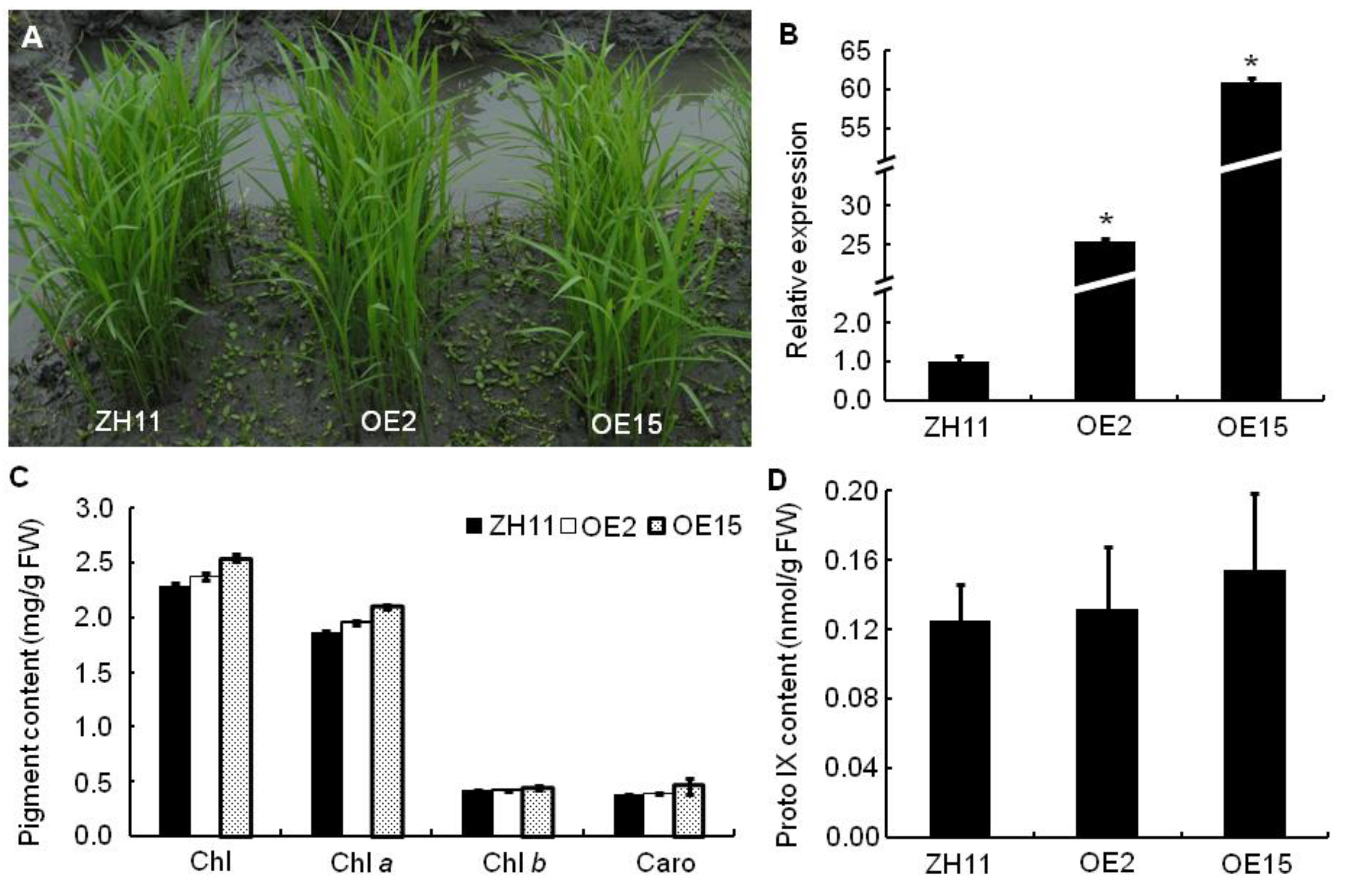
2.11. Overexpression Analyses of OsPPO1
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Paraffin Sectioning and Transmission Electron Microscopy Analyses
4.3. Histochemical Analysis
4.4. Photosynthetic Pigment and Proto IX Quantification
4.5. Vector Construction and Rice Transformation
4.6. Subcellular Localization of the OsPPO1 Protein
4.7. Gene Expression Analysis
4.8. Herbicide Treatment
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhu, B.Y.; Chen, C.P.; Yuan, Z.D.; Guo, J.; Yang, X.R.; Wang, S.; Lv, Y.; Liu, Q.S.; Yang, B.; et al. A single nucleotide substitution of GSAM gene causes massive accumulation of glutamate 1-semialdehyde and yellow leaf phenotype in rice. Rice 2021, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.H.; Liu, L.C.; Tang, J.Y.; Lin, A.H.; Zhang, F.T.; Fang, J.; Zhang, G.F.; Chu, C.C. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J. Genet. Genom. 2011, 38, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Hur, J.; Ryu, C.H.; Choi, Y.; Chung, Y.Y.; Miyao, A.; Hirochika, H.; An, G. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol. 2003, 44, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.T.; Li, J.J.; Yoo, J.H.; Yoo, S.C.; Cho, S.H.; Koh, H.J.; Seo, H.S.; Paek, N.C. Rice Chlorina-1 and Chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol. Biol. 2006, 62, 325–337. [Google Scholar] [CrossRef]
- Kong, W.Y.; Yu, X.W.; Chen, H.Y.; Liu, L.L.; Xiao, Y.J.; Wang, Y.L.; Wang, C.L.; Lin, Y.; Yu, Y.; Wang, C.M.; et al. The catalytic subunit of magnesium-protoporphyrin IX monomethyl ester cyclase forms a chloroplast complex to regulate chlorophyll biosynthesis in rice. Plant Mol. Biol. 2016, 92, 177–191. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Rahman, M.L.; Cho, S.H.; Kim, Y.S.; Koh, H.J.; Yoo, S.C.; Peak, N.C. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J. 2013, 74, 122–133. [Google Scholar] [CrossRef]
- Wang, P.R.; Gao, J.X.; Wan, C.M.; Zhang, F.T.; Xu, Z.J.; Huang, X.Q.; Sun, X.Q.; Deng, X.J. Divinyl chlorophyll(ide) α can be converted to monovinyl chlorophyll(ide) α by a divinyl reductase in rice. Plant Physiol. 2010, 153, 994–1003. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.M.; Zhang, X.; He, B.; Diao, L.P.; Sheng, S.L.; Wang, J.L.; Guo, X.P.; Su, N.; Wang, L.F.; Jiang, L.; et al. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol. 2007, 145, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, J.H.; Yoo, E.S.; Lee, C.H.; Hirochika, H.; An, G. Differential regulation of chlorophyll α oxygenase genes in rice. Plant Mol. Biol. 2005, 57, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Xu, J.; Huang, L.C.; Leng, Y.J.; Dai, L.P.; Rao, Y.C.; Chen, L.; Wang, Y.Q.; Tu, Z.J.; Hu, J.; et al. PGL, encoding chlorophyllide a oxygenase 1, impacts leaf senescence and indirectly affects grain yield and quality in rice. J. Exp. Bot. 2016, 67, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- Izawa, T.; Oikawa, T.; Tokutomi, S.; Okuno, K.; Shimamoto, K. Phytochromes confer the photoperiodic control of flowering in rice (a short-day plant). Plant J. 2000, 22, 391–399. [Google Scholar] [CrossRef]
- Li, Q.Z.; Zhu, F.Y.; Gao, X.L.; Sun, Y.; Li, S.J.; Tao, Y.Z.; Lo, C.; Liu, H.J. Young Leaf Chlorosis 2 encodes the stroma-localized heme oxygenase 2 which is required for normal tetrapyrrole biosynthesis in rice. Planta 2014, 240, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Okumoto, Y.; Yoshitake, Y.; Inoue, H.; Yuan, Q.B.; Teraishi, M.; Tsukiyama, T.; Nishida, H.; Tanisaka, T. Complete loss of photoperiodic response in the rice mutant line X61 is caused by deficiency of phytochrome chromophore biosynthesis gene. Theor. Appl. Genet. 2011, 122, 109–118. [Google Scholar] [CrossRef]
- Narita, S.I.; Tanaka, R.; Ito, T.; Okada, K.; Taketani, S.; Inokuchi, H. Molecular cloning and characterization of a cDNA that encodes protoporphyrinogen oxidase of Arabidopsis thaliana. Gene 1996, 182, 169–175. [Google Scholar] [CrossRef]
- Lermontova, I.; Kruse, E.; Mock, H.P.; Grimm, B. Cloning and characterization of a plastidal and a mitochondrial isoform of tobacco protoporphyrinogen IX oxidase. Proc. Natl. Acad. Sci. USA 1997, 94, 8895–8900. [Google Scholar] [CrossRef] [Green Version]
- Che, F.S.; Watanabe, N.; Iwano, M.; Inokuchi, H.; Takayama, S.; Yoshida, S.; Isogai, A. Molecular characterization and subcellular localization of protoporphyrinogen oxidase in spinach chloroplasts. Plant Physiol. 2000, 124, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Che, F.S.; Iwano, M.; Takayama, S.; Yoshida, S.; Isogai, A. Dual targeting of spinach protoporphyrinogen oxidase II to mitochondria and chloroplasts by alternative use of two in-frame initiation codons. J. Biol. Chem. 2001, 276, 20474–20481. [Google Scholar] [CrossRef] [Green Version]
- Sasarman, A.; Letowski, J.; Czaika, G.; Ramirez, V.; Nead, M.A.; Jacobs, J.M.; Morais, R. Nucleotide sequence of the hemG gene involved in the protoporphyrinogen oxidase activity of Escherichia coli K12. Can. J. Microbiol. 1993, 39, 1155–1161. [Google Scholar] [CrossRef]
- Nishimura, K.; Taketani, S.; Inokuchi, H. Cloning of a human cDNA for protoporphyrinogen oxidase by complementation in vivo of a hemG mutant of Escherichia coli. J. Biol. Chem. 1995, 270, 8076–8080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camadro, J.M.; Labbe, P. Cloning and characterization of the yeast HEM14 gene coding for protoporphyrinogen oxidase, the molecular target of diphenyl ether-type herbicides. J. Biol. Chem. 1996, 271, 9120–9128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, A.; Volrath, S.; Guyer, D.; Maleck, K.; Ryals, J.; Ward, E. Inhibition of protoporphyrinogen oxidase expression in Arabidopsis causes a lesion-mimic phenotype that induces systemic acquired resistance. Plant J. 1999, 17, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Tang, W.J.; Hedtke, B.; Zhong, L.L.; Liu, L.; Peng, L.W.; Lu, C.M.; Grimm, B.; Lin, R.C. Tetrapyrrole biosynthetic enzyme protoporphyrinogen IX oxidase 1 is required for plastid RNA editing. Proc. Natl. Acad. Sci. USA 2014, 111, 2023–2028. [Google Scholar] [CrossRef] [Green Version]
- Li, J.L.; Zhang, F.; Li, Y.H.; Yang, W.C.; Lin, R.C. Chloroplast-localized protoporphyrinogen IX oxidase1 is involved in the mitotic cell cycle in Arabidopsis. Plant Cell Physiol. 2019, 60, 2436–2448. [Google Scholar] [CrossRef]
- Lermontova, I.; Grimm, B. Reduced activity of plastid protoporphyrinogen oxidase causes attenuated photodynamic damage during high-light compared to low-light exposure. Plant J. 2006, 48, 499–510. [Google Scholar] [CrossRef]
- Jung, S.; Lee, H.J.; Lee, Y.; Kang, K.; Kim, Y.S.; Grimm, B.; Back, K. Toxic tetrapyrrole accumulation in protoporphyrinogen IX oxidase-overexpressing transgenic rice plants. Plant Mol. Biol. 2008, 67, 535–546. [Google Scholar] [CrossRef]
- Lermontova, I.; Grimm, B. Overexpression of plastidic protoporphyrinogen IX oxidase leads to resistance to the diphenyl-ether herbicide acifluorfen. Plant Physiol. 2000, 122, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Kuk, Y.I.; Chung, J.S.; Lee, S.B.; Choi, K.W.; Han, O.S.; Guh, J.O. Physiology and growth of transgenic tobacco plants containing Bacillus subtilis protoporphyrinogen oxidase gene in response to oxyfluorfen treatment. Korean J. Weed Sci. 1998, 18, 237–245. [Google Scholar]
- Lee, H.J.; Lee, S.B.; Chung, J.S.; Han, S.U.; Han, O.; Guh, J.O.; Jeon, J.S.; An, G.; Back, K. Transgenic rice plants expressing a Bacillus subtilis protoporphyrinogen oxidase gene are resistant to diphenyl ether herbicide oxyfluorfen. Plant Cell Physiol. 2000, 41, 743–749. [Google Scholar] [CrossRef]
- Kuk, Y.I.; Lee, H.J.; Chung, J.S.; Kim, K.M.; Lee, S.B.; Ha, S.B.; Back, K.; Guh, J.O. Expression of a Bacillus subtilis protoporphyrinogen oxidase gene in rice plants reduces sensitivity to peroxidizing herbicides. Biol. Plant. 2005, 49, 577–583. [Google Scholar] [CrossRef]
- Ha, S.B.; Lee, S.B.; Lee, Y.; Yang, K.; Lee, N.; Jang, S.M.; Chung, J.S.; Jung, S.; Kim, Y.S.; Wi, S.G.; et al. The plastidic Arabidopsis protoporphyrinogen IX oxidase gene, with or without the transit sequence, confers resistance to the diphenyl ether herbicide in rice. Plant Cell Environ. 2003, 27, 79–88. [Google Scholar] [CrossRef]
- Lee, Y.; Jung, S.; Back, K. Expression of human protoporphyrinogen oxidase in transgenic rice induces both a photodynamic response and oxyfluorfen resistance. Pestic. Biochem. Physiol. 2004, 80, 65–74. [Google Scholar] [CrossRef]
- Jung, S.; Lee, Y.; Yang, K.; Lee, S.B.; Jang, S.M.; Ha, S.B.; Back, K. Dual targeting of Myxococcus xanthus protoporphyrinogen oxidase into chloroplasts and mitochondria and high level oxyfluorfen resistance. Plant Cell Environ. 2004, 27, 1436–1446. [Google Scholar] [CrossRef]
- Park, J.; Tran, L.H.; Jung, S. A protoporphyrinogen oxidase gene expression influences responses of transgenic rice to oxyfluorfen. Biol. Plant. 2017, 61, 659–666. [Google Scholar] [CrossRef]
- Cui, Y.J.; Peng, Y.L.; Zhang, Q.; Xia, S.S.; Ruan, B.P.; Xu, Q.K.; Yu, X.Q.; Zhou, T.T.; Liu, H.; Zeng, D.L.; et al. Disruption of EARLY LESION LEAF 1, encoding a cytochrome P450 monooxygenase, induces ROS accumulation and cell death in rice. Plant J. 2021, 105, 942–956. [Google Scholar] [CrossRef]
- Wierenga, R.K.; Terpstra, P.; Hol, W.G.J. Prediction of the occurrence of the ADP-binding βαβ-fold in proteins, using an amino acid sequence fingerprint. J. Mol. Biol. 1986, 187, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- Apitz, J.; Nishimura, K.; Schmied, J.; Wolf, A.; Hedtke, B.; van Wijk, K.J.; Grimm, B. Posttranslational control of ALA synthesis includes GluTR degradation by Clp protease and stabilization by GluTR-binding protein. Plant Physiol. 2016, 170, 2040–2051. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.J.; Wang, W.Q.; Chen, D.Q.; Ji, Q.; Jing, Y.J.; Wang, H.Y.; Lin, R.C. Transposase-derived proteins FHY3/FAR1 interact with Phytochrome-Interacting factor1 to regulate chlorophyll biosynthesis by modulating HEMB1 during deetiolation in Arabidopsis. Plant Cell 2012, 24, 1984–2000. [Google Scholar] [CrossRef] [Green Version]
- Li, C.M.; Liu, X.; Pan, J.H.; Guo, J.; Wang, Q.; Chen, C.P.; Li, N.; Zhang, K.; Yang, B.; Sun, C.H.; et al. A lil3 chlp double mutant with exclusive accumulation of geranylgeranyl chlorophyll displays a lethal phenotype in rice. BMC Plant Biol. 2019, 19, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Higo, K.I.; Higo, H.; Minobe, Y.; Matsui, H.; Chiba, S. Nucleotide and derived amino acid sequence of a catalase cDNA isolated from rice immature seeds. Plant Mol. Biol. 1992, 18, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Morita, S.; Tasaka, M.; Fujisawa, H.; Ushimaru, T.; Tsuji, H. A cDNA clone encoding a rice catalase isozyme. Plant Physiol. 1994, 105, 1015–1016. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Jwa, N.S.; Iwahashi, H.; Rakwal, R. Importance of ascorbate peroxidases OsAPX1 and OsAPX2 in the rice pathogen response pathways and growth and reproduction revealed by their transcriptional profiling. Gene 2003, 322, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Saisho, D.; Nakazono, M.; Tsutsumi, N.; Hirai, A. Transcript levels of tandem-arranged alternative oxidase genes in rice are increased by low temperature. Gene 1997, 203, 121–129. [Google Scholar] [CrossRef]
- Zou, L.P.; Sun, X.H.; Zhang, Z.G.; Liu, P.; Wu, J.X.; Tian, C.J.; Qiu, J.L.; Lu, T.G. Leaf rolling controlled by the homeodomain leucine zipper class IV gene Roc5 in rice. Plant Physiol. 2011, 156, 1589–1602. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, Y.H.; Long, Q.Z.; Huang, J.X.; Wang, Y.L.; Zhou, K.N.; Zheng, M.; Sun, J.; Chen, H.; Chen, S.H.; et al. Overexpression of OsZHD1, a zinc finger homeodomain class homeobox transcription factor, induces abaxially curled and drooping leaf in rice. Planta 2014, 239, 803–816. [Google Scholar] [CrossRef]
- Chen, Q.L.; Xie, Q.J.; Gao, J.; Wang, W.Y.; Sun, B.; Liu, B.H.; Zhu, H.T.; Peng, H.F.; Zhao, H.B.; Liu, C.H.; et al. Characterization of Rolled and Erect Leaf 1 in regulating leave morphology in rice. J. Exp. Bot. 2015, 66, 6047–6058. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Q.; Li, W.Q.; Miao, H.; Gan, P.F.; Qiao, L.; Chang, Y.L.; Shi, C.H.; Chen, K.M. REL2, a gene encoding an unknown function protein which contains DUF630 and DUF632 domains controls leaf rolling in rice. Rice 2016, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Xiang, J.J.; Zhang, G.H.; Qian, Q.; Xue, H.W. SEMI-ROLLED LEAF1 encodes a putative glycosylphosphatidylinositol-anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells. Plant Physiol. 2012, 159, 1488–1500. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.C.; Chen, J.; Zeng, L.R.; Goh, M.L.; Leung, H.; Khush, G.S.; Wang, G.L. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol. Plant Microbe Interact. 2000, 13, 869–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.R.; Wan, C.M.; Xu, Z.J.; Wang, P.Y.; Wang, W.M.; Sun, C.H.; Ma, X.Z.; Xiao, Y.H.; Zhu, J.Q.; Gao, X.L.; et al. One divinyl reductase reduces the 8-vinyl groups in various intermediates of chlorophyll biosynthesis in a given higher plant species, but the isozyme differs between species. Plant Physiol. 2013, 161, 521–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Su, J.B.; Duan, S.; Ao, Y.; Dai, J.R.; Liu, J.; Wang, P.; Li, Y.G.; Liu, B.; Feng, D.R.; et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 2011, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, N.; Hu, M.L.; Wu, D.X.; Wu, F.Q.; Fei, G.L.; Lan, Y.; Chen, X.L.; Shu, X.L.; Zhang, X.; Guo, X.P.; et al. Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production. Plant Physiol. 2012, 159, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.M.; Hu, Y.; Huang, R.; Ma, X.Z.; Wang, Y.; Liao, T.T.; Zhong, P.; Xiao, F.L.; Sun, C.H.; Xu, Z.J.; et al. Mutation of FdC2 gene encoding a ferreoxin-like protein with C-terminal extension causes yellow-green leaf phenotype in rice. Plant Sci. 2015, 238, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Inkagai, N.; Kinoshita, K.; Kagawa, T.; Tanaka, A.; Ueno, O.; Shimada, H.; Takano, M. Phytochrome B mediates the regulation of chlorophyll biosynthesis through transcriptional regulation of ChlH and GUN4 in rice seedlings. PLoS ONE 2015, 10, e0135408. [Google Scholar] [CrossRef]
- Yang, S.L.; Fang, G.N.; Zhang, A.P.; Ruan, B.P.; Jiang, H.Z.; Ding, S.L.; Liu, C.L.; Zhang, Y.; Jaha, N.; Hu, P.; et al. Rice EARLY SENESCENCE 2, encoding an inositol polyphosphate kinase, is involved in leaf senescence. BMC Plant Biol. 2020, 20, 393. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Deng, X.-J.; Li, C.-Y.; Xiao, Y.-K.; Zhao, K.; Guo, J.; Yang, X.-R.; Zhang, H.-S.; Chen, C.-P.; Luo, Y.-T.; et al. Mutation of Protoporphyrinogen IX Oxidase Gene Causes Spotted and Rolled Leaf and Its Overexpression Generates Herbicide Resistance in Rice. Int. J. Mol. Sci. 2022, 23, 5781. https://doi.org/10.3390/ijms23105781
Liu X, Deng X-J, Li C-Y, Xiao Y-K, Zhao K, Guo J, Yang X-R, Zhang H-S, Chen C-P, Luo Y-T, et al. Mutation of Protoporphyrinogen IX Oxidase Gene Causes Spotted and Rolled Leaf and Its Overexpression Generates Herbicide Resistance in Rice. International Journal of Molecular Sciences. 2022; 23(10):5781. https://doi.org/10.3390/ijms23105781
Chicago/Turabian StyleLiu, Xin, Xiao-Jian Deng, Chun-Yan Li, Yong-Kang Xiao, Ke Zhao, Jia Guo, Xiao-Rong Yang, Hong-Shan Zhang, Cong-Ping Chen, Ya-Ting Luo, and et al. 2022. "Mutation of Protoporphyrinogen IX Oxidase Gene Causes Spotted and Rolled Leaf and Its Overexpression Generates Herbicide Resistance in Rice" International Journal of Molecular Sciences 23, no. 10: 5781. https://doi.org/10.3390/ijms23105781
APA StyleLiu, X., Deng, X. -J., Li, C. -Y., Xiao, Y. -K., Zhao, K., Guo, J., Yang, X. -R., Zhang, H. -S., Chen, C. -P., Luo, Y. -T., Tang, Y. -L., Yang, B., Sun, C. -H., & Wang, P. -R. (2022). Mutation of Protoporphyrinogen IX Oxidase Gene Causes Spotted and Rolled Leaf and Its Overexpression Generates Herbicide Resistance in Rice. International Journal of Molecular Sciences, 23(10), 5781. https://doi.org/10.3390/ijms23105781