
Anti-Inflammatory Effects of Psoralen Derivatives on RAW264.7 Cells via Regulation of the NF-κB and MAPK Signaling Pathways



Abstract
:1. Introduction
2. Results
2.1. Effect of Psoralen Derivatives on the Viability of RAW 264.7 Cells
2.2. Effect of Psoralen Derivatives on NO Production of RAW 264.7 Cells
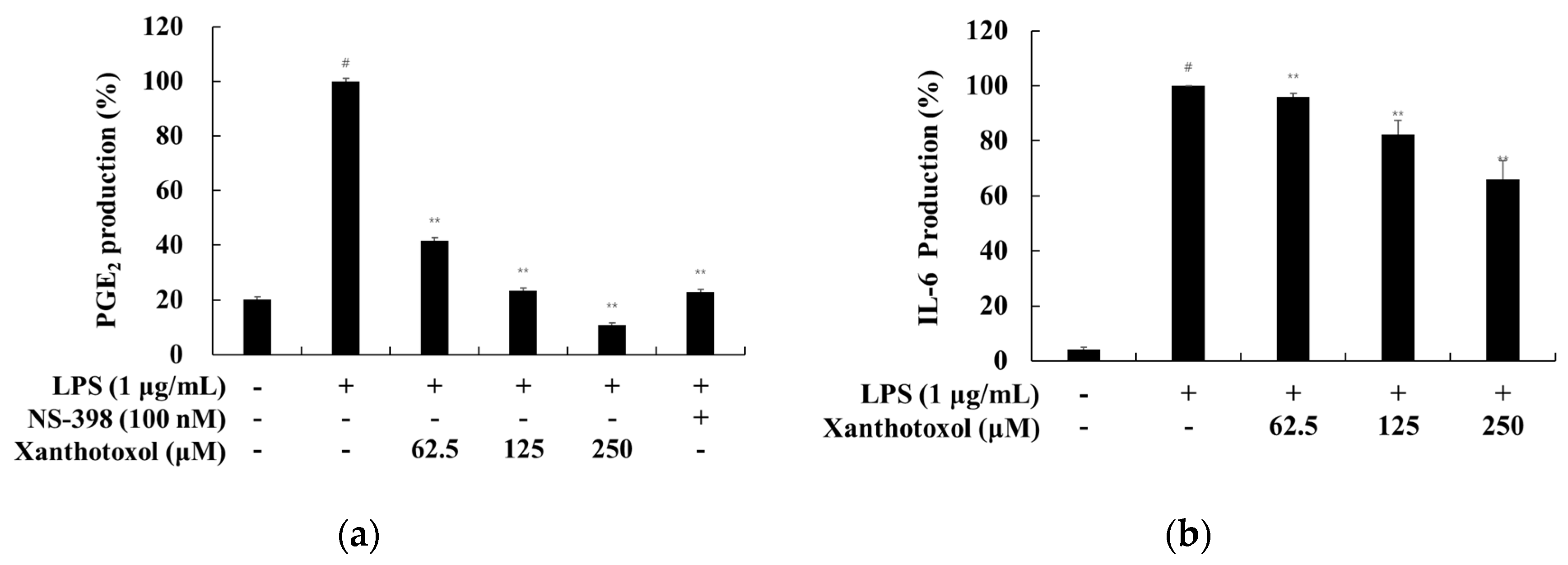
2.3. Effect of Xanthotoxol on PGE2 and Inflammatory Cytokines
2.4. Effect of Xanthotoxol on INOS and COX-2 Production
2.5. Effect of Xanthotoxol on the MAPK Signaling Pathway
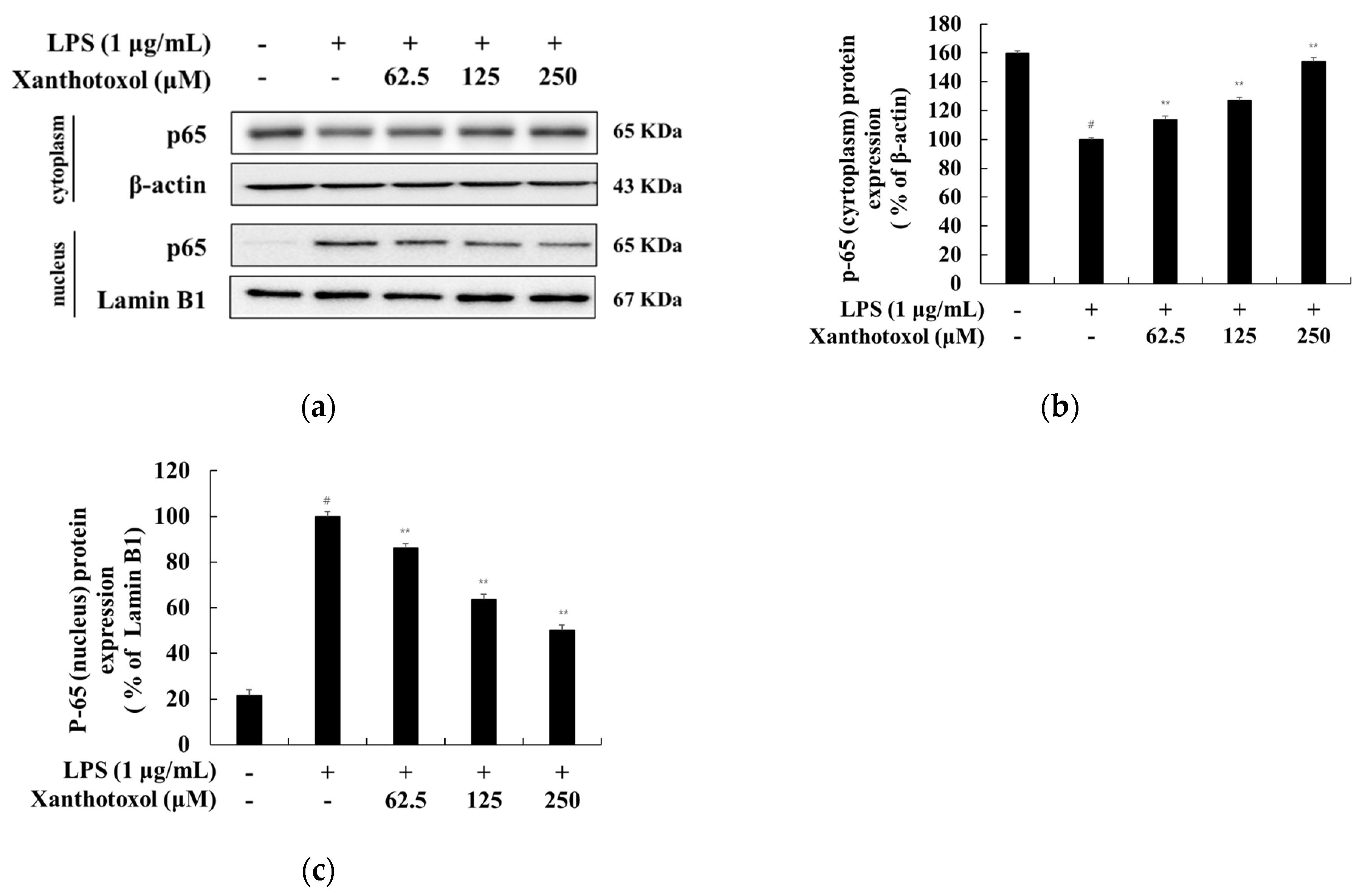
2.6. Effect of Xanthotoxol on the NF-κB Signaling Pathway
2.7. Skin Primary Irritation Test
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability
4.4. Nitric Oxide
4.5. PGE2 and Cytokines
4.6. Preparation of Nuclear and Cytoplasmic Extraction
4.7. Western Blotting
4.8. Human Skin Patch Test
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, B.; Zhang, Y. Teaching an old dog new tricks: Drug discovery by repositioning natural products and their derivatives. Drug Discov. Today 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41. [Google Scholar] [CrossRef] [PubMed]
- Barreca, M.; Spanò, V.; Raimondi, M.V.; Bivacqua, R.; Giuffrida, S.; Montalbano, A.; Cavalli, A.; Bertoni, F.; Barraja, P. GPCR Inhibition in Treating Lymphoma. ACS Med. Chem. Let. 2022, 13, 358–364. [Google Scholar] [CrossRef]
- Spanò, V.; Barreca, M.; Cilibrasi, V.; Genovese, M.; Renda, M.; Montalbano, A.; Galietta, L.J.V.; Barraja, P. Evaluation of Fused Pyrrolothiazole Systems as Correctors of Mutant CFTR Protein. Molecules 2021, 26, 1275. [Google Scholar] [CrossRef]
- Barreca, M.; Ingarra, A.M.; Raimondi, M.V.; Spanò, V.; De Franco, M.; Menilli, L.; Gandin, V.; Miolo, G.; Barraja, P.; Montalbano, A. Insight on pyrimido[5,4-g]indolizine and pyrimido[4,5-c]pyrrolo[1,2-a]azepine systems as promising photosensitizers on malignant cells. Eur. J. Med. Chem. 2022, 237, 114399. [Google Scholar] [CrossRef]
- Cilibrasi, V.; Spanò, V.; Bortolozzi, R.; Barreca, M.; Raimondi, M.V.; Rocca, R.; Maruca, A.; Montalbano, A.; Alcaro, S.; Ronca, R.; et al. Synthesis of 2H-Imidazo[2′,1′:2,3] [1,3]thiazolo[4,5-e]isoindol-8-yl-phenylureas with promising therapeutic features for the treatment of acute myeloid leukemia (AML) with FLT3/ITD mutations. Eur. J. Med. Chem. 2022, 235, 114292. [Google Scholar] [CrossRef]
- Ullah, S.; Chung, Y.C.; Hyun, C.G. Induction of Melanogenesis by Fosfomycin in B16F10 Cells Through the Upregulation of P-JNK and P-p38 Signaling Pathways. Antibiotics 2020, 9, 172. [Google Scholar] [CrossRef]
- Moon, S.H.; Chung, Y.C.; Hyun, C.G. Tobramycin Promotes Melanogenesis by Upregulating p38 MAPK Protein Phosphorylation in B16F10 Melanoma Cells. Antibiotics 2019, 8, 140. [Google Scholar] [CrossRef] [Green Version]
- Hyun, S.B.; Chung, Y.C.; Hyun, C.G. Nojirimycin suppresses inflammation via regulation of NF-κ B signaling pathways. Pharmazie 2020, 75, 637. [Google Scholar]
- Kang, H.K.; Hyun, C.G. Anti-inflammatory Effect of d -(+)-Cycloserine Through Inhibition of NF-κB and MAPK Signaling Pathways in LPS-Induced RAW 264.7 Macrophages. Nat. Prod. Commun. 2020, 15, 1934578X20920481. [Google Scholar] [CrossRef]
- Kang, J.K.; Kang, H.K.; Hyun, C.G. Anti-inflammatory effects of spiramycin in LPS-activated RAW 264.7 macrophages. Molecules 2022, 27, 3202. [Google Scholar] [CrossRef]
- Rastelli, G.; Pellati, F.; Pinzi, L.; Gamberini, M.C. Repositioning Natural Products in Drug Discovery. Molecules 2020, 25, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T.; International Natural Product Sciences Taskforce. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200. [Google Scholar] [CrossRef] [PubMed]
- Ribaudo, G.; Memo, M.; Gianoncelli, A. A Perspective on Natural and Nature-Inspired Small Molecules Targeting Phosphodiesterase 9 (PDE9): Chances and Challenges against Neurodegeneration. Pharmaceuticals 2021, 14, 58. [Google Scholar] [CrossRef] [PubMed]
- Mazzini, S.; Musso, L.; Dallavalle, S.; Artali, R. Putative SARS-CoV-2 Mpro Inhibitors from an In-House Library of Natural and Nature-Inspired Products: A Virtual Screening and Molecular Docking Study. Molecules 2020, 25, 3745. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Hyun, C.G. Inhibitory Effects of Pinostilbene on Adipogenesis in 3T3-L1 Adipocytes: A Study of Possible Mechanisms. Int. J. Mol. Sci. 2021, 22, 13446. [Google Scholar] [CrossRef]
- Chung, Y.C.; Hyun, C.G. Inhibitory Effects of Pinostilbene Hydrate on Melanogenesis in B16F10 Melanoma Cells via ERK and p38 Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 4732. [Google Scholar] [CrossRef]
- Chung, Y.C.; Kim, S.; Kim, J.H.; Lee, G.S.; Lee, J.N.; Lee, N.H.; Hyun, C.G. Pratol, an O-Methylated Flavone, Induces Melanogenesis in B16F10 Melanoma Cells via p-p38 and p-JNK Upregulation. Molecules 2017, 22, 1704. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, S.S.; Park, K.J.; An, H.J.; Choi, Y.H.; Lee, N.H.; Hyun, C.G. Methyl jasmonate inhibits lipopolysaccharide-induced inflammatory cytokine production via mitogen-activated protein kinase and nuclear factor-κB pathways in RAW 264.7 cells. Pharmazie 2016, 71, 540. [Google Scholar]
- Yoon, W.J.; Ham, Y.M.; Yoon, H.S.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Acanthoic acid inhibits melanogenesis through tyrosinase downregulation and melanogenic gene expression in B16 melanoma cells. Nat. Prod. Commun. 2013, 8, 1359. [Google Scholar]
- Kim, Y.J.; Lee, G.H.; Kwong, B.Y.; Martires, K.J. Evidence-based, Skin-directed Treatments for Cutaneous Chronic Graft-versus-host Disease. Cureus 2019, 11, e6462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marka, A.; Carter, J.B. Phototherapy for Cutaneous T-Cell Lymphoma. Dermatol. Clin. 2020, 38, 127. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Montalbano, A.; Spanò, V.; Musante, I.; Galietta, L.J.V.; Barraja, P. Furocoumarins as multi-target agents in the treatment of cystic fibrosis. Eur. J. Med. Chem. 2019, 180, 283. [Google Scholar] [CrossRef] [PubMed]
- Buhimschi, A.D.; Gooden, D.M.; Jing, H.; Fels, D.R.; Hansen, K.S.; Beyer, W.F., Jr.; Dewhirst, M.W.; Walder, H.; Gasparro, F.P. Psoralen Derivatives with Enhanced Potency. Photochem Photobiol. 2020, 96, 1014. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.; Rodrigues, L.R. Biosynthesis and heterologous production of furanocoumarins: Perspectives and current challenges. Nat. Prod. Rep. 2021, 38, 869. [Google Scholar] [CrossRef]
- Bonte, F.; Dumas, M.; Chaudagne, C.; Meybeck, A. Influence of asiatic acid, madecassic acid, and asiaticoside on human collagen I synthesis. Planta Med. 1994, 60, 133. [Google Scholar] [CrossRef]
- Maquart, F.X.; Bellon, G.; Gillery, P.; Wegrowski, Y.; Borel, J.P. Stimulation of collagen synthesis in fibroblast cultures by a triterpene extracted from Centella Asiatica. Connect. Tissue Res. 1990, 24, 107. [Google Scholar] [CrossRef]
- Lee, J.; Jung, E.; Park, J.; Jung, K.; Lee, S.; Hong, S.; Park, J.; Park, E.; Kim, J.; Park, S.; et al. Anti-inflammatory effects of magnolol and honokiol are mediated through inhibition of the downstream pathway of MEKK-1 in NF-kappaB activation signaling. Planta Med. 2005, 71, 338. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.; Jung, E.; Park, Y.; Kim, K.; Park, B.; Jung, K.; Park, E.; Kim, J.; Park, D. In vitro antibacterial and anti-inflammatory effects of honokiol and magnolol against Propionibacterium sp. Eur. J. Pharmacol. 2004, 496, 189. [Google Scholar] [CrossRef]
- Zhang, H.L.; Wu, X.Y.; Mi, J.; Peng, Y.J.; Wang, Z.G.; Liu, Y.; Wu, X.L.; Gao, Y. A New Anti-Inflammatory Alkaloid from Roots of Heracleum dissectum. Chem. Biodivers. 2017, 14, e1700184. [Google Scholar] [CrossRef]
- Kang, J.K.; Chung, Y.C.; Hyun, C.G. Anti-Inflammatory Effects of 6-Methylcoumarin in LPS-Stimulated RAW 264.7 Macrophages via Regulation of MAPK and NF-κB Signaling Pathways. Molecules 2021, 26, 5351. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, X.; Ruan, C.C.; Cheang, W.S. Two methoxy derivatives of resveratrol, 3,3′,4,5′-tetramethoxy-trans-stilbene and 3,4′,5-trimethoxy-trans-stilbene, suppress lipopolysaccharide-induced inflammation through inactivation of MAPK and NF-κB pathways in RAW 264.7 cells. Chin. Med. 2021, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Cheong, K.A.; Lee, A.Y. Increased Skin Irritation by Hydroquinone and Rsetinoic Acid Used in Combination. Ann. Dermatol. 2017, 29, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, S.B.; Bae, S.; Hyun, C.G. Antioxidant Activities of Jeju Wax Apple (Syzygium samarangense) and Safety of Human Keratinocytes and Primary Skin Irritation Test. Cosmetics 2020, 7, 39. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Test Samples | No. of Responder | 24 h | 48 h | Reaction Grade | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +1 | +2 | +3 | +4 | +1 | +2 | +3 | +4 | 24 h | 48 h | Mean | |||
| 1 | Xanthotoxol (100 μM) | 0 | - | - | - | - | - | - | - | - | 0 | 0 | 0 |
| 2 | Control (Squalene) | 0 | - | - | - | - | - | - | - | - | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Hyun, C.-G. Anti-Inflammatory Effects of Psoralen Derivatives on RAW264.7 Cells via Regulation of the NF-κB and MAPK Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 5813. https://doi.org/10.3390/ijms23105813
Lee Y, Hyun C-G. Anti-Inflammatory Effects of Psoralen Derivatives on RAW264.7 Cells via Regulation of the NF-κB and MAPK Signaling Pathways. International Journal of Molecular Sciences. 2022; 23(10):5813. https://doi.org/10.3390/ijms23105813
Chicago/Turabian StyleLee, Yeji, and Chang-Gu Hyun. 2022. "Anti-Inflammatory Effects of Psoralen Derivatives on RAW264.7 Cells via Regulation of the NF-κB and MAPK Signaling Pathways" International Journal of Molecular Sciences 23, no. 10: 5813. https://doi.org/10.3390/ijms23105813
APA StyleLee, Y., & Hyun, C. -G. (2022). Anti-Inflammatory Effects of Psoralen Derivatives on RAW264.7 Cells via Regulation of the NF-κB and MAPK Signaling Pathways. International Journal of Molecular Sciences, 23(10), 5813. https://doi.org/10.3390/ijms23105813