
HDACs Gene Family Analysis of Eight Rosaceae Genomes Reveals the Genomic Marker of Cold Stress in Prunus mume


Abstract
:1. Introduction
2. Results
2.1. Identification of Rosaceae HDACs Gene Families
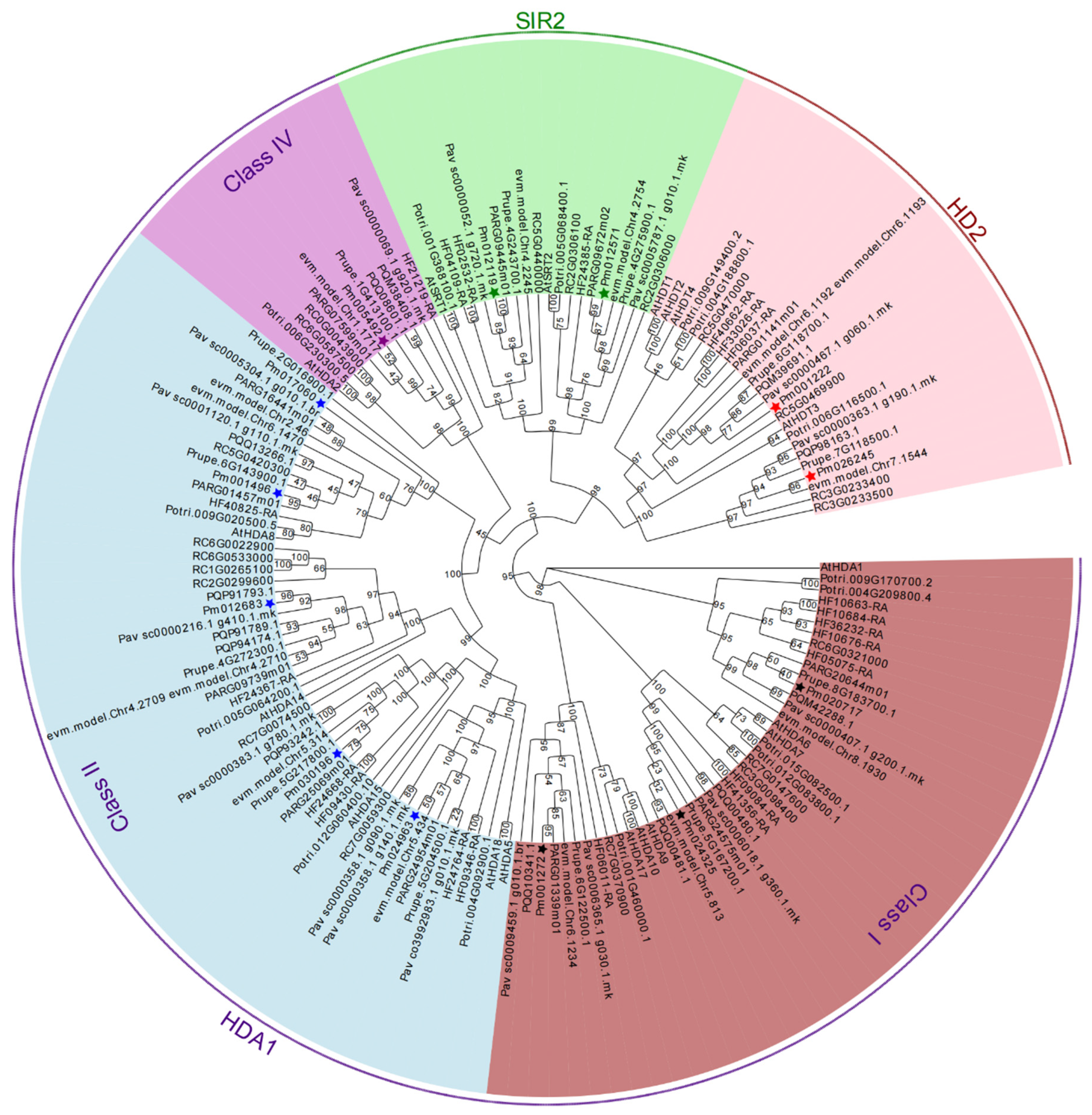
2.2. Multiple Sequences Alignment, Phylogenetic and Classification Analysis of RoHDACs
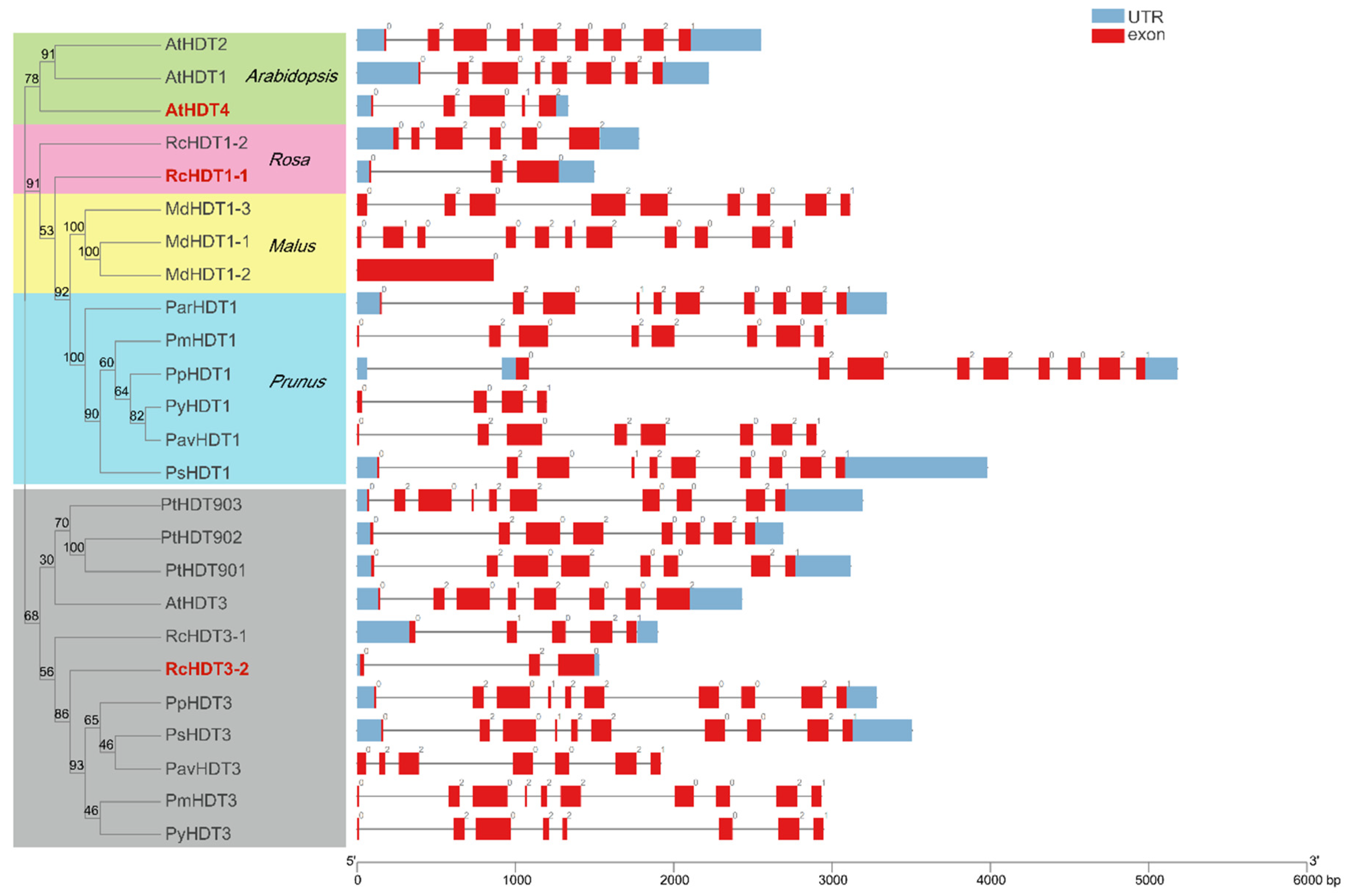
2.3. Gene Structure and Conversed Domains Analysis
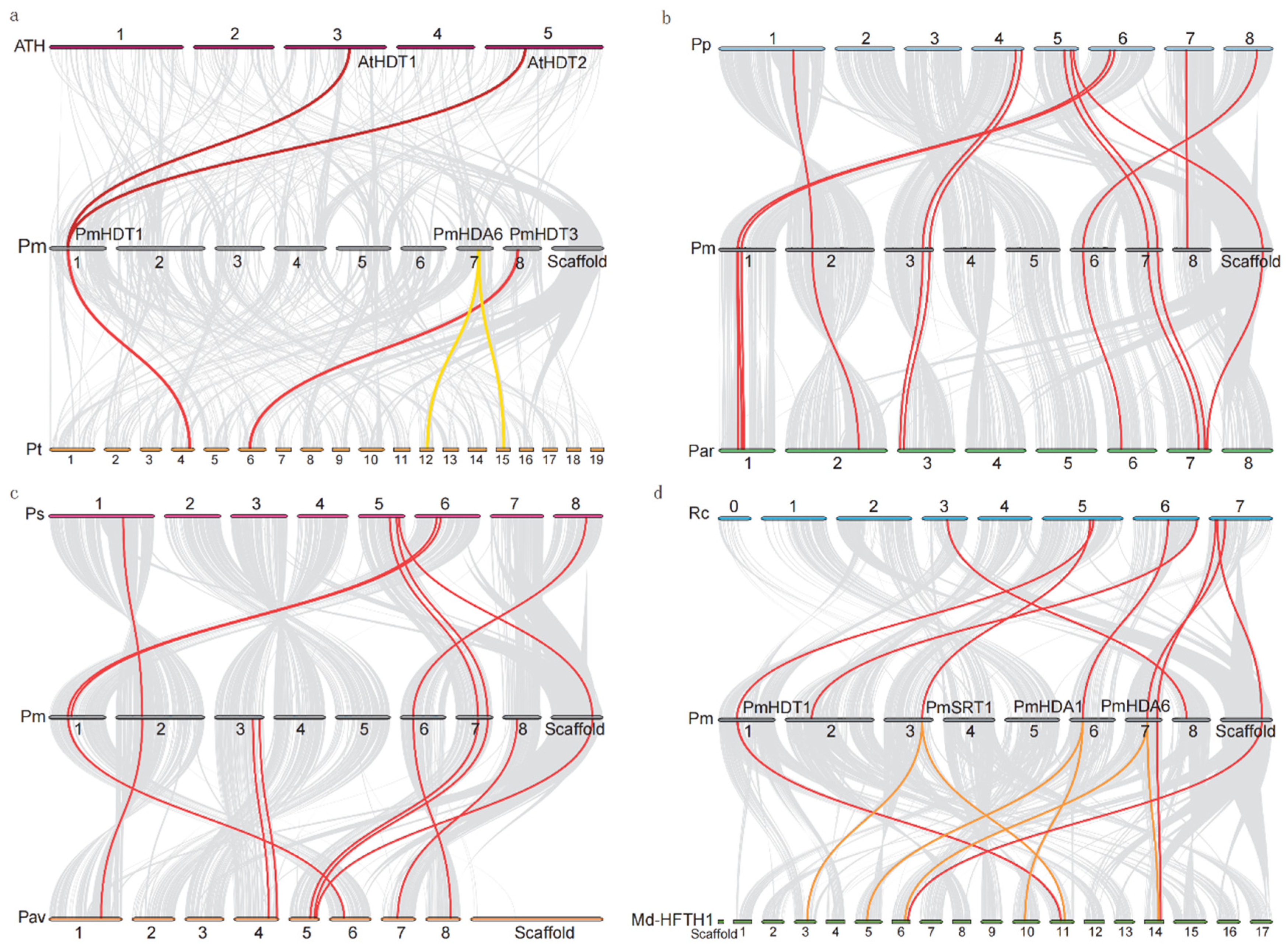
2.4. RoHDACs Gene Locations on Chromosome, Segmental Duplications and Synteny Analysis
2.5. Cis-Element of PmHDACs
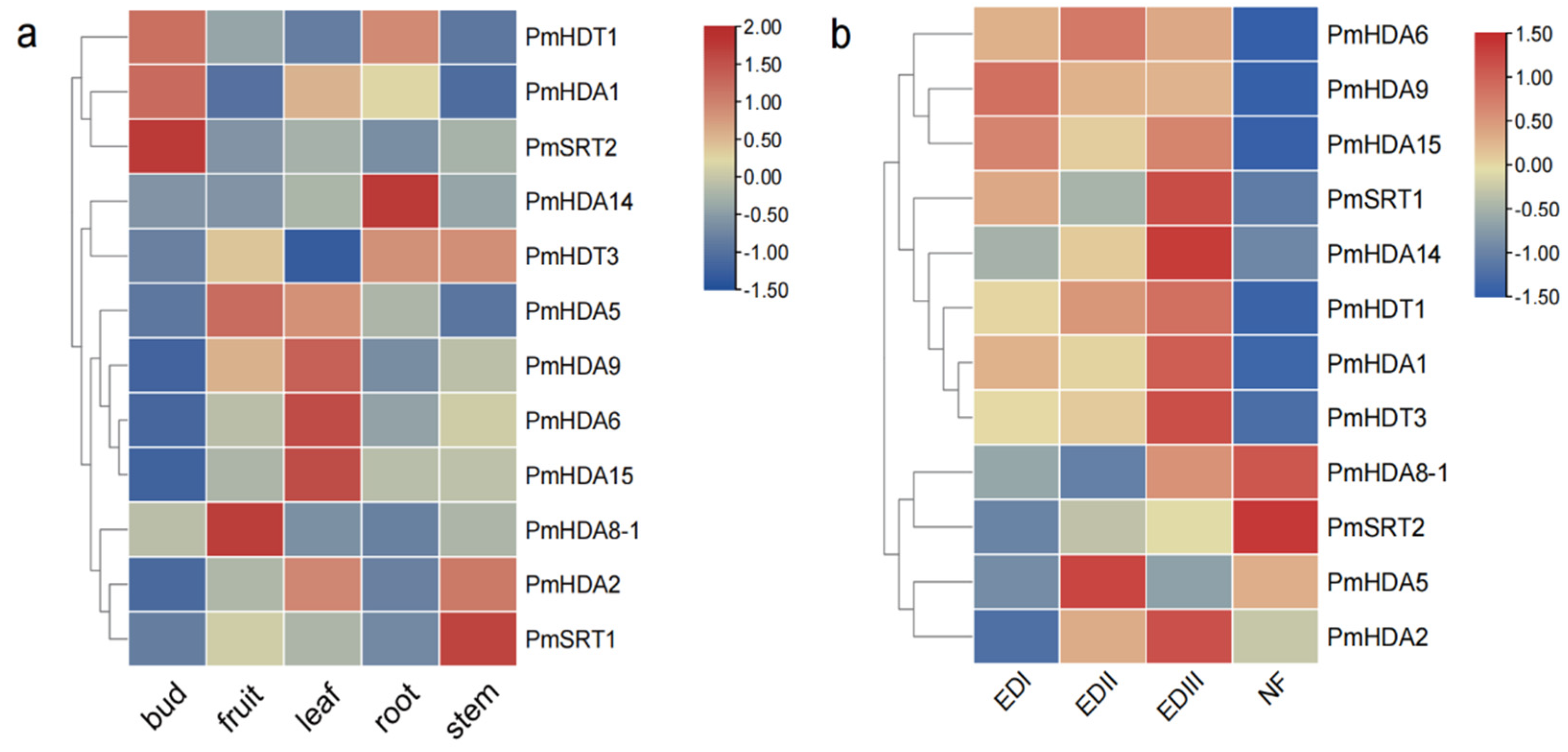
2.6. PmHDACs Expression Profiles in Tissues and Treatments
2.7. qRT-PCR of PmHDACs under Cold Stress and Gene Functional Annotation
3. Discussion
4. Materials and Methods
4.1. Plants Genome Resources
4.2. Identification of HDACs Gene Family
4.3. Phylogenetic Analysis and Classification of HDACs Genes
4.4. Gene Structure and Protein Conserved Motif Analysis
4.5. Duplications, Synteny and Genes Chromosome Location Analysis
4.6. PmHDACs Cis-Acting Element Analysis
4.7. Expression Profiles of PmHDACs
4.8. Plant Materials, Cold Stress Treatments and qRT-PCR
4.9. GO Annotation and Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Campbell, R.R.; Wood, M.A. How the epigenome integrates information and reshapes the synapse. Nat. Rev. Neurosci. 2019, 20, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Loidl, P. A plant dialect of the histone language. Trends Plant Sci. 2004, 9, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Pazin, M.J.; Kadonaga, J.T. What’s Up and Down with Histone Deacetylation and Transcription. Cell 1997, 89, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Kiermaier, A.; Eilers, M. Transcriptional control: Calling in histone deacetylase. Curr. Biol. 1997, 7, R505–R507. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.H.; Allis, C.D. Roles of histone acetyltransferases and deacetylases in gene regulation. BioEssays News Rev. Mol. Cell. Dev. Biol. 1998, 20, 615–626. [Google Scholar] [CrossRef]
- Shen, Y.; Wei, W.; Zhou, D. Histone Acetylation Enzymes Coordinate Metabolism and Gene Expression. Trends Plant Sci. 2015, 20, 614–621. [Google Scholar] [CrossRef]
- Sharon, Y.; Roth, J.M.D.A. Histone acetyltransferases. Annu. Rev. Biochem. 2001, 70, 81–120. [Google Scholar] [CrossRef]
- Gerald Brosch, M.G.P.L. Purification of histone deacetylase HD 1-A of germinating maize embryos. FEBS Lett. 1996, 393, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Rossi, V.; Hartings, H.; Motto, M. Identification and characterisation of an RPD3 homologue from maize (Zea mays L.) that is able to complement an rpd3 null mutant of Saccharomyces cerevisiae. Mol. Gen. Genet. MGG 1998, 258, 288. [Google Scholar] [CrossRef]
- Lusser, A.; Brosch, G.; Loidl, A.; Haas, H.; Loidl, P. Identification of maize histone deacetylase HD2 as an acidic nucleolar phosphoprotein. Science 1997, 277, 88–91. [Google Scholar] [CrossRef]
- Wu, K.; Tian, L.; Malik, K.; Brown, D.; Miki, B. Functional analysis of HD2 histone deacetylase homologues in Arabidopsis thaliana. Plant J. 2000, 22, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Alinsug, M.V.; Yu, C.; Wu, K. Phylogenetic analysis, subcellular localization, and expression patterns of RPD3/HDA1 family histone deacetylases in plants. BMC Plant Biol. 2009, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey Ritu, E.A. Analysis of histone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatin modification among multicellular eukaryotes. Nucleic Acids Res. 2002, 30, 5033–5036. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Wu, K.; Duan, J. Sequence and expression analysis of histone deacetylases in rice. Biochem. Biophys. Res. Commun. 2007, 356, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Lv, S.; Zhang, C.; Yang, C. Histone deacetylases and their functions in plants. Plant Cell Rep. 2013, 32, 465–478. [Google Scholar] [CrossRef]
- Aquea, F.; Timmermann, T.; Arce-Johnson, P. Analysis of histone acetyltransferase and deacetylase families of Vitis vinifera. Plant Physiol. Biochem. 2010, 48, 194–199. [Google Scholar] [CrossRef]
- Hou, J.; Ren, R.; Xiao, H.; Chen, Z.; Yu, J.; Zhang, H.; Shi, Q.; Hou, H.; He, S.; Li, L. Characteristic and evolution of HAT and HDAC genes in Gramineae genomes and their expression analysis under diverse stress in Oryza sativa. Planta 2021, 253, 72. [Google Scholar] [CrossRef]
- Yang, C.; Shen, W.; Chen, H.; Chu, L.; Xu, Y.; Zhou, X.; Liu, C.; Chen, C.; Zeng, J.; Liu, J.; et al. Characterization and subcellular localization of histone deacetylases and their roles in response to abiotic stresses in soybean. BMC Plant Biol. 2018, 18, 226. [Google Scholar] [CrossRef]
- Eom, S.H.; Hyun, T.K. Comprehensive Analysis of the Histone Deacetylase Gene Family in Chinese Cabbage (Brassica rapa): From Evolution and Expression Pattern to Functional Analysis of BraHDA3. Agriculture 2021, 11, 244. [Google Scholar] [CrossRef]
- Jin, P.; Gao, S.; He, L.; Xu, M.; Zhang, T.; Zhang, F.; Jiang, Y.; Liu, T.; Yang, J.; Yang, J.; et al. Genome-Wide Identification and Expression Analysis of the Histone Deacetylase Gene Family in Wheat (Triticum aestivum L.). Plants 2021, 10, 19. [Google Scholar] [CrossRef]
- Yuan, L.; Dai, H.; Zheng, S.; Huang, R.; Tong, H. Genome-wide identification of the HDAC family proteins and functional characterization of CsHD2C, a HD2-type histone deacetylase gene in tea plant (Camellia sinensis L. O. Kuntze). Plant Physiol. Bioch. 2020, 155, 898–913. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Shafiq, S.; Naeem, M.K.; Widemann, E.; Munir, M.Z.; Jensen, K.B.; Wang, R.R.C. Histone Deacetylase (HDAC) Gene Family in Allotetraploid Cotton and Its Diploid Progenitors: In Silico Identification, Molecular Characterization, and Gene Expression Analysis under Multiple Abiotic Stresses, DNA Damage and Phytohormone Treatments. Int. J. Mol. Sci. 2020, 21, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Lu, J.; Zhang, J.; Wu, P.; Yang, S.; Wu, K. Identification and characterization of histone deacetylases in tomato (Solanum lycopersicum). Front. Plant Sci. 2015, 5, 760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogan, N.T.; Hogan, K.; Long, J.A. APETALA2 negatively regulates multiple floral organ identity genes in Arabidopsis by recruiting the co-repressor TOPLESS and the histone deacetylase HDA19. Development 2012, 139, 4180–4190. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Chang, K.; Wu, K. Genome-Wide Analysis of Gene Regulatory Networks of the FVE-HDA6-FLD Complex in Arabidopsis. Front. Plant Sci. 2016, 7, 555. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.; Bowen, A.J.; Carroll, T.S.; Conlan, R.S. The Transcription Corepressor LEUNIG Interacts with the Histone Deacetylase HDA19 and Mediator Components MED14 (SWP) and CDK8 (HEN3) To Repress Transcription. Mol. Cell. Biol. 2007, 27, 5306–5315. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Latrasse, D.; Servet, C.; Zhou, D. Arabidopsis histone deacetylase HDA9 regulates flowering time through repression of AGL19. Biochem. Bioph. Res. Commun. 2013, 432, 394–398. [Google Scholar] [CrossRef]
- de Rooij, P.G.H.; Perrella, G.; Kaiserli, E.; van Zanten, M. The diverse and unanticipated roles of histone deacetylase 9 in coordinating plant development and environmental acclimation. J. Exp. Bot. 2020, 71, 6211–6225. [Google Scholar] [CrossRef]
- Jiang, D.; Wang, Y.; Wang, Y.; He, Y. Repression of FLOWERING LOCUS C and FLOWERING LOCUS T by the Arabidopsis Polycomb repressive complex 2 components. PLoS ONE 2008, 3, e3404. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J. Abiotic stress responses in plants. Nat. Reviews. Genet. 2021, 23, 104–119. [Google Scholar] [CrossRef]
- Zhu, J. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Luo, M.; Wang, Y.; Wu, K. Involvement of Arabidopsis histone deacetylase HDA6 in ABA and salt stress response. J. Exp. Bot. 2010, 61, 3345–3353. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wu, K. Role of histone deacetylases HDA6 and HDA19 in ABA and abiotic stress response. Plant Signal. Behav. 2010, 5, 1318–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Ge, J.; Bao, C.; Chang, W.; Liu, J.; Shao, J.; Liu, X.; Su, L.; Pan, L.; Zhou, D.X. Histone deacetylase HDA9 and transcription factor WRKY53 are mutual antagonists in regulation of plant stress response. Mol. Plant 2019, 13, 598–611. [Google Scholar] [CrossRef]
- Jung, J.H.; Park, J.H.; Lee, S.; To, T.K.; Kim, J.M.; Seki, M.; Park, C.M. The cold signaling attenuator HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE1 activates FLOWERING LOCUS C transcription via chromatin remodeling under short-term cold stress in Arabidopsis. Plant Cell 2013, 25, 4378–4390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, T.K.; Nakaminami, K.; Kim, J.; Morosawa, T.; Ishida, J.; Tanaka, M.; Yokoyama, S.; Shinozaki, K.; Seki, M. Arabidopsis HDA6 is required for freezing tolerance. Biochem. Bioph. Res. Commun. 2011, 406, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; To, T.K.; Seki, M. An epigenetic integrator: New insights into genome regulation, environmental stress responses and developmental controls by histone deacetylase 6. Plant Cell Physiol. 2012, 53, 794–800. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Wang, Y.; Liu, X.; Yang, S.; Lu, Q.; Cui, Y.; Wu, K. HD2C interacts with HDA6 and is involved in ABA and salt stress response in Arabidopsis. J. Exp. Bot. 2012, 63, 3297–3306. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Cheng, K.; Xu, Y.; Yang, S.; Wu, K. Plant Responses to Abiotic Stress Regulated by Histone Deacetylases. Front. Plant Sci. 2017, 8, 2147. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Wang, Y.Y.; Liu, X.; Yang, S.; Wu, K. HD2 proteins interact with RPD3-type histone deacetylases. Plant Signal. Behav. 2012, 7, 608–610. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Yu, H.; Zhao, Z.; Hunter, D.; Luo, X.; Duan, J.; Tian, L. AtHD2D Gene Plays a Role in Plant Growth, Development, and Response to Abiotic Stresses in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Hu, J.; Han, X.; Li, J.; Gao, Y.; Richards, C.M.; Zhang, C.; Tian, Y.; Liu, G.; Gul, H.; et al. A high-quality apple genome assembly reveals the association of a retrotransposon and red fruit colour. Nat. Commun. 2019, 10, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Shen, F.; Chen, Y.; Cao, K.; Wang, L. Chromosome-scale genome assembly and population genomics provide insights into the adaptation, domestication, and flavonoid metabolism of Chinese plum. Plant J. 2021, 108, 1174–1192. [Google Scholar] [CrossRef] [PubMed]
- Groppi, A.; Liu, S.; Cornille, A.; Decroocq, S.; Bui, Q.T.; Tricon, D.; Cruaud, C.; Arribat, S.; Belser, C.; Marande, W.; et al. Population genomics of apricots unravels domestication history and adaptive events. Nat. Commun. 2021, 12, 3956. [Google Scholar] [CrossRef] [PubMed]
- Bourque, S.; Jeandroz, S.; Grandperret, V.; Lehotai, N.; Aimé, S.; Soltis, D.E.; Miles, N.W.; Melkonian, M.; Deyholos, M.K.; Leebens-Mack, J.H.; et al. The Evolution of HD2 Proteins in Green Plants. Trends Plant Sci. 2016, 21, 1008–1016. [Google Scholar] [CrossRef]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Liu, X.; Yang, S.; Zhao, M.; Luo, M.; Yu, C.; Chen, C.; Tai, R.; Wu, K. Transcriptional Repression by Histone Deacetylases in Plants. Mol. Plant 2014, 7, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Hollender, C.; Liu, Z. Histone Deacetylase Genes in Arabidopsis Development. J. Integr. Plant Biol. 2008, 50, 875–885. [Google Scholar] [CrossRef]
- Zhou, C.; Labbe, H.; Sridha, S.; Wang, L.; Tian, L.; Latoszek-Green, M.; Yang, Z.; Brown, D.; Miki, B.; Wu, K. Expression and function of HD2-type histone deacetylases in Arabidopsis development. Plant J. 2004, 38, 715–724. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Jin, H.; Noh, Y.; Noh, B. Repression of flowering under a noninductive photoperiod by the HDA9-AGL19-FT module in Arabidopsis. New Phytol. 2015, 206, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Wani, S.H.; Roychoudhury, A. Epigenetic Control of Plant Cold Responses. Front. Plant Sci. 2017, 8, 1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Yang, Y.; Yang, T.; Ba, L.; Zhang, H.; Han, Y.; Xiao, Y.; Shan, W.; Kuang, J.; Chen, J.; et al. MaMYB4 Recruits Histone Deacetylase MaHDA2 and Modulates the Expression of ω-3 Fatty Acid Desaturase Genes during Cold Stress Response in Banana Fruit. Plant Cell Physiol. 2019, 60, 2410–2422. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Zhang, L.; Zhou, C.; Yu, C.W.; Chaikam, V. HDA6 is required for jasmonate response, senescence and flowering in Arabidopsis. J. Exp. Bot. 2008, 59, 225–234. [Google Scholar] [CrossRef]
- Baek, D.; Shin, G.; Kim, M.C.; Shen, M.; Lee, S.Y.; Yun, D. Histone Deacetylase HDA9 With ABI4 Contributes to Abscisic Acid Homeostasis in Drought Stress Response. Front. Plant Sci. 2020, 11, 143. [Google Scholar] [CrossRef]
- Yang, L.; Chen, X.; Wang, Z.; Sun, Q.; Hong, A.; Zhang, A.; Zhong, X.; Hua, J. HOS15 and HDA9 negatively regulate immunity through histone deacetylation of intracellular immune receptor NLR genes in Arabidopsis. New Phytol. 2020, 226, 507–522. [Google Scholar] [CrossRef]
- Park, H.J.; Baek, D.; Cha, J.; Liao, X.; Kang, S.; McClung, C.R.; Lee, S.Y.; Yun, D.; Kim, W. HOS15 Interacts with the Histone Deacetylase HDA9 and the Evening Complex to Epigenetically Regulate the Floral Activator GIGANTEA. Plant Cell 2019, 31, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Jeong, J.C.; Zhu, Y.; Sokolchik, I.; Miyazaki, S.; Zhu, J.K.; Hasegawa, P.M.; Bohnert, H.J.; Shi, H.; Yun, D.J.; et al. Involvement of Arabidopsis HOS15 in histone deacetylation and cold tolerance. Proc. Natl. Acad. Sci. USA 2008, 105, 4945–4950. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Lim, C.J.; Shen, M.; Park, H.J.; Cha, J.; Iniesto, E.; Rubio, V.; Mengiste, T.; Zhu, J.; Bressan, R.A.; et al. Epigenetic switch from repressive to permissive chromatin in response to cold stress. Proc. Natl. Acad. Sci. USA 2018, 115, E5400–E5409. [Google Scholar] [CrossRef] [Green Version]
- Tong, B.; Xia, D.; Lv, S.; Ma, X. Cloning and expression analysis of PtHDT903, a HD2-type histone deacetylase gene in Populus trichocarpa. Biotechnol. Biotechnol. Equip. 2018, 32, 1098–1104. [Google Scholar] [CrossRef] [Green Version]
- Tahir, M.S.; Tian, L. HD2-type histone deacetylases: Unique regulators of plant development and stress responses. Plant Cell Rep. 2021, 40, 1603–1615. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Dong, Y.; Yue, X.; Hu, J.; Jiang, S.; Xu, H.; Wang, Y.; Su, M.; Zhang, J.; Zhang, Z.; et al. The B-box zinc finger protein MdBBX20 integrates anthocyanin accumulation in response to ultraviolet radiation and low temperature. Plant Cell Environ. 2019, 42, 2090–2104. [Google Scholar] [CrossRef] [PubMed]
- Renkova, A.; Valitova, J.; Schaller, H.; Minibayeva, F. The homoeologous genes encoding C24-sterol methyltransferase 1 in Triticum aestivum: Structural characteristics and effects of cold stress. Biol. Plantarum 2019, 63, 59–69. [Google Scholar] [CrossRef]
- Wang, N.; Qu, C.; Jiang, S.; Chen, Z.; Xu, H.; Fang, H.; Su, M.; Zhang, J.; Wang, Y.; Liu, W.; et al. The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples. Plant J. 2018, 96, 39–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.; Lee, T.; Cheng, C.; Buble, K.; Zheng, P.; Yu, J.; Humann, J.; Ficklin, S.P.; Gasic, K.; Scott, K.; et al. 15 years of GDR: New data and functionality in the Genome Database for Rosaceae. Nucleic Acids Res. 2019, 47, D1137–D1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Mistry, J.; Tate, J.; Coggill, P.; Heger, A.; Pollington, J.E.; Gavin, O.L.; Gunasekaran, P.; Ceric, G.; Forslund, K.; et al. The Pfam protein families database. Nucleic Acids Res. 2010, 38, D211–D222. [Google Scholar] [CrossRef]
- Ponting, C.P.; Schultz, J.; Milpetz, F.; Bork, P. SMART: Identification and annotation of domains from signalling and extracellular protein sequences. Nucleic Acids Res. 1999, 27, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifinopoulos, J.; Lam-Tung, N.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Chengjie Chen, H.C.Y.H. TBtools, a Toolkit for Biologists integrating various biological data handling tools with a user-friendly interface. Mol. Plant 2018, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Chao, J.; Li, Z.; Sun, Y.; Aluko, O.O.; Wu, X.; Wang, Q.; Liu, G. MG2C: A user-friendly online tool for drawing genetic maps. Mol. Hortic. 2021, 1, 16. [Google Scholar] [CrossRef]
- Walsh, B. Population-Genetic Models of the Fates of Duplicate Genes. Genetica 2003, 118, 279–294. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W.; Sun, L.; Zhao, F.; Huang, B.; Yang, W.; Tao, Y.; Wang, J.; Yuan, Z.; Fan, G.; et al. The genome of Prunus mume. Nat. Commun. 2012, 3, 1318. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome Profiles Reveal the Crucial Roles of Hormone and Sugar in the Bud Dormancy of Prunus mume. Sci. Rep. 2018, 8, 5090. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | A.thaliana | P.trichocarpa | P.mume | P.armeniaca | P.salicina | P.persica | P.avium | P.yedoensis | M.domenstica | R.chinensis |
|---|---|---|---|---|---|---|---|---|---|---|
| HDA1/Class I | 5 | 5 | 3 | 3 | 3 | 3 | 4 | 4 | 8 | 4 |
| HDA1/Class II | 6 | 4 | 5 | 5 | 5 | 5 | 7 | 5 | 6 | 7 |
| HDA1/Class IV | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 2 |
| HD2 | 4 | 3 | 2 | 1 | 2 | 2 | 2 | 2 | 3 | 4 |
| SIR2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | / | 3 | 3 |
| Total numbers | 18 | 15 | 13 | 12 | 13 | 13 | 16 | 13 | 21 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, J.; Wen, Z.; Li, M.; Cheng, T.; Zhang, Q.; Sun, L. HDACs Gene Family Analysis of Eight Rosaceae Genomes Reveals the Genomic Marker of Cold Stress in Prunus mume. Int. J. Mol. Sci. 2022, 23, 5957. https://doi.org/10.3390/ijms23115957
Meng J, Wen Z, Li M, Cheng T, Zhang Q, Sun L. HDACs Gene Family Analysis of Eight Rosaceae Genomes Reveals the Genomic Marker of Cold Stress in Prunus mume. International Journal of Molecular Sciences. 2022; 23(11):5957. https://doi.org/10.3390/ijms23115957
Chicago/Turabian StyleMeng, Juan, Zhenying Wen, Mingyu Li, Tangren Cheng, Qixiang Zhang, and Lidan Sun. 2022. "HDACs Gene Family Analysis of Eight Rosaceae Genomes Reveals the Genomic Marker of Cold Stress in Prunus mume" International Journal of Molecular Sciences 23, no. 11: 5957. https://doi.org/10.3390/ijms23115957
APA StyleMeng, J., Wen, Z., Li, M., Cheng, T., Zhang, Q., & Sun, L. (2022). HDACs Gene Family Analysis of Eight Rosaceae Genomes Reveals the Genomic Marker of Cold Stress in Prunus mume. International Journal of Molecular Sciences, 23(11), 5957. https://doi.org/10.3390/ijms23115957