
Epigenomic Analysis Reveals the KCNK9 Potassium Channel as a Potential Therapeutic Target for Adenomyosis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
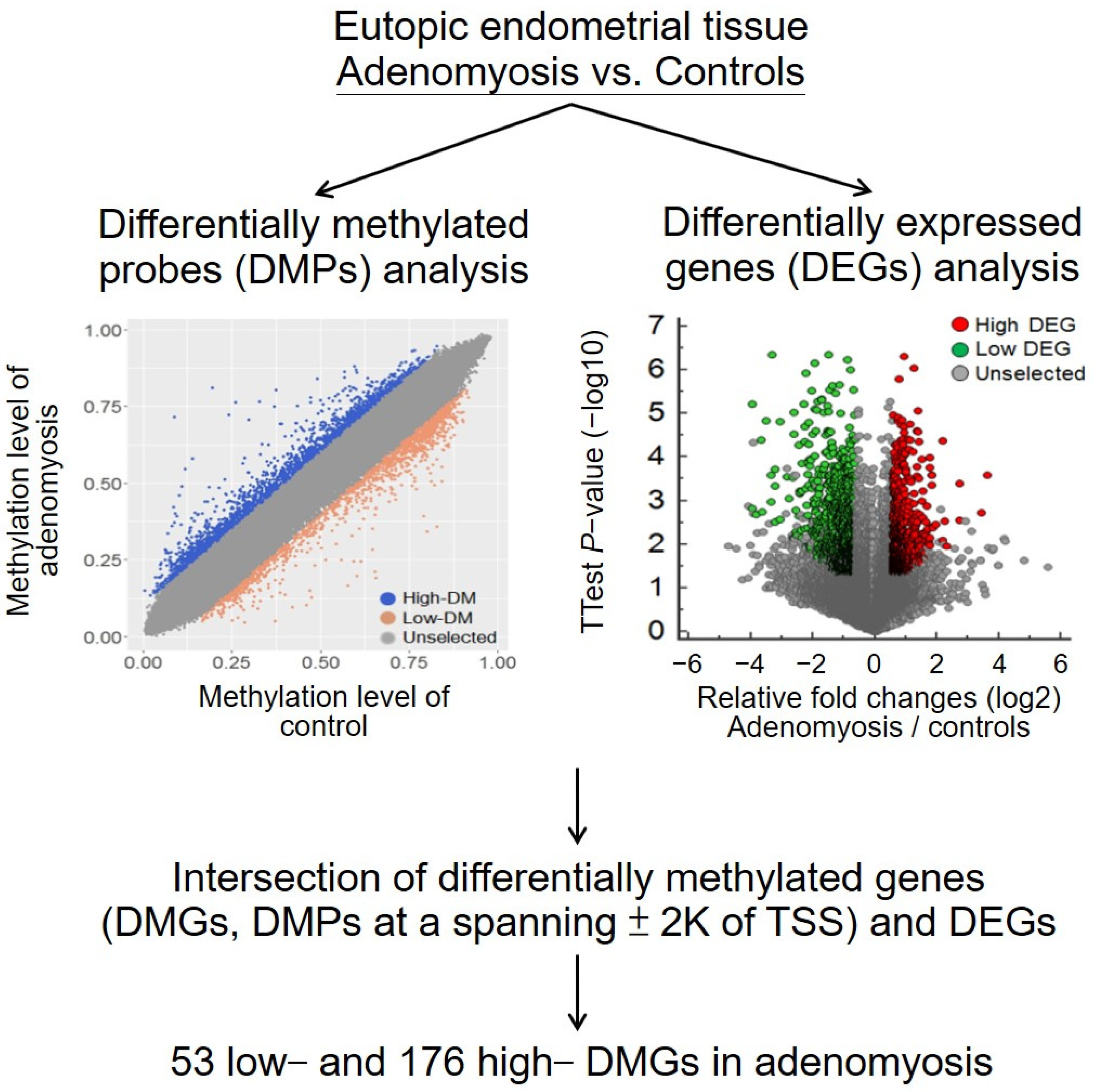
2.1. Differential Methylomic and Transcriptomic Profiles of Eutopic Endometrial Tissues in Adenomyosis and Control Groups
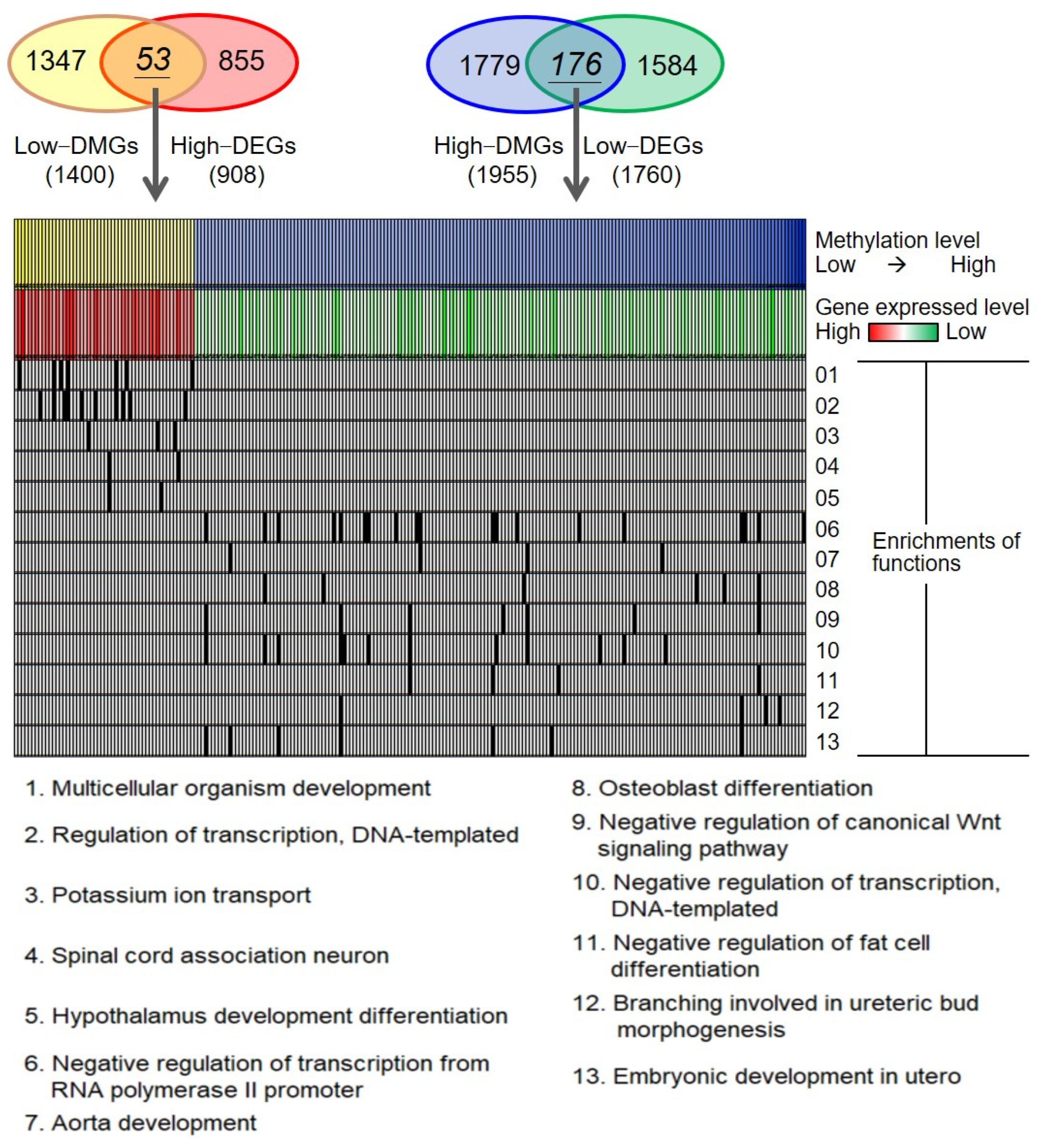
2.2. Biological Functions of Adenomyosis-Associated Epigenetic Changes
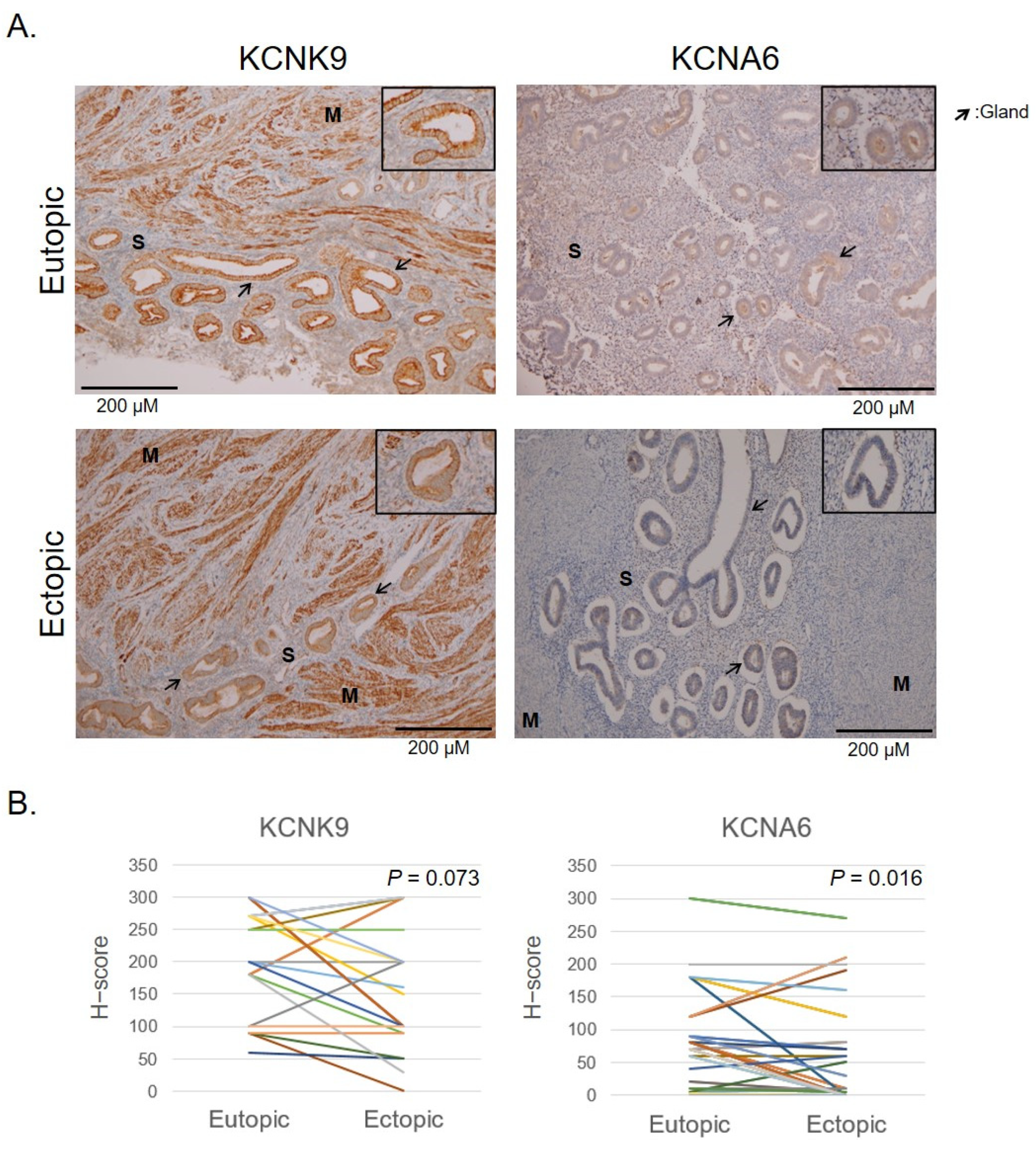
2.3. Verification of Hypomethylated Genes
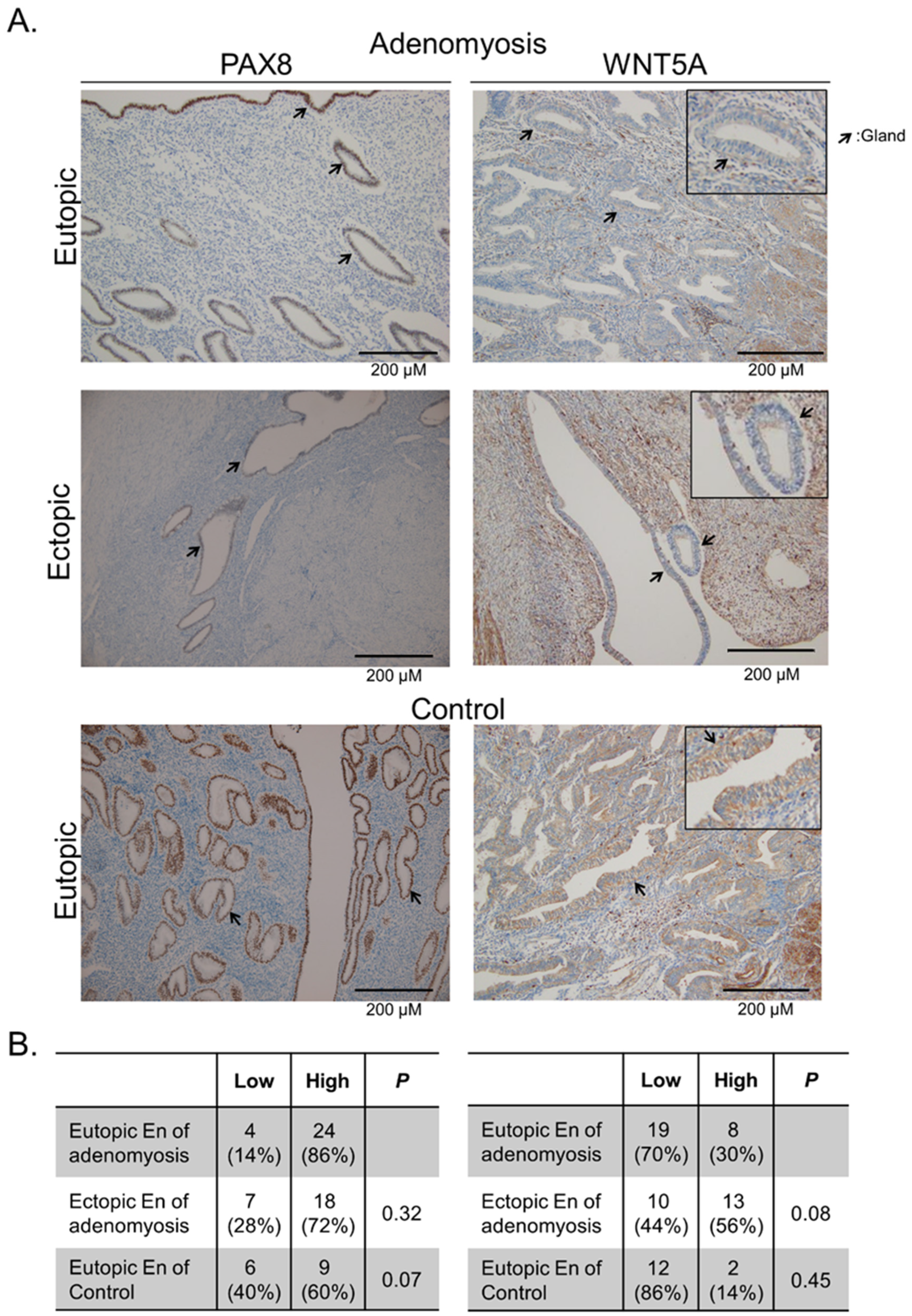
2.4. Verification of Hypermethylated Genes
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. DNA Extraction and Bisulfite Conversion
4.3. Methylomic and Transcriptomic Data Analysis
4.4. Functional Enrichment Analysis
4.5. Immunohistochemistry Staining
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GnRH | gonadotropin-releasing hormone |
| UTR | untranslated region |
| DEGs | differentially expressed genes |
| K2P | two-pore domain potassium |
| VGKCs | voltage-gated potassium channels |
| TSS | transcriptional start site |
| IHC | immunohistochemistry |
| FFPE | formalin-fixed paraffin-embedded |
References
- Benagiano, G.; Brosens, I. History of adenomyosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Benagiano, G.; Brosens, I.; Habiba, M. Structural and molecular features of the endomyometrium in endometriosis and adenomyosis. Hum. Reprod. Update 2014, 20, 386–402. [Google Scholar] [CrossRef]
- Harada, T.; Khine, Y.M.; Kaponis, A.; Nikellis, T.; Decavalas, G.; Taniguchi, F. The Impact of Adenomyosis on Women’s Fertility. Obstet. Gynecol. Surv. 2016, 71, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Vannuccini, S.; Clifton, V.L.; Fraser, I.S.; Taylor, H.S.; Critchley, H.; Giudice, L.C.; Petraglia, F. Infertility and reproductive disorders: Impact of hormonal and inflammatory mechanisms on pregnancy outcome. Hum. Reprod. Update 2016, 22, 104–115. [Google Scholar] [CrossRef]
- Kitawaki, J. Adenomyosis: The pathophysiology of an oestrogen-dependent disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 493–502. [Google Scholar] [CrossRef]
- Streuli, I.; Dubuisson, J.; Santulli, P.; de Ziegler, D.; Batteux, F.; Chapron, C. An update on the pharmacological management of adenomyosis. Expert Opin. Pharmacother. 2014, 15, 2347–2360. [Google Scholar] [CrossRef]
- Vannuccini, S.; Tosti, C.; Carmona, F.; Huang, S.J.; Chapron, C.; Guo, S.W.; Petraglia, F. Pathogenesis of adenomyosis: An update on molecular mechanisms. Reprod. BioMed. Online 2017, 35, 592–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrarelli, P.; Yen, C.F.; Funghi, L.; Arcuri, F.; Tosti, C.; Bifulco, G.; Luddi, A.; Lee, C.L.; Petraglia, F. Expression of Inflammatory and Neurogenic Mediators in Adenomyosis. Reprod. Sci. 2017, 24, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Chang, J.A.; Kim, Y.A.; Chang, S.H.; Chun, K.C.; Koh, J.W. Innervation in women with uterine myoma and adenomyosis. Obstet. Gynecol. Sci. 2015, 58, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.W.; Mao, X.; Ma, Q.; Liu, X. Dysmenorrhea and its severity are associated with increased uterine contractility and overexpression of oxytocin receptor (OTR) in women with symptomatic adenomyosis. Fertil. Steril. 2013, 99, 231–240. [Google Scholar] [CrossRef]
- Campo, S.; Campo, V.; Benagiano, G. Infertility and adenomyosis. Obstet. Gynecol. Int. 2012, 2012, 786132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, C.P.; Kayisili, U.; Taylor, H.S. HOXA10 expression is decreased in endometrium of women with adenomyosis. Fertil. Steril. 2011, 95, 1133–1136. [Google Scholar] [CrossRef] [PubMed]
- Kokcu, A. A current view of the role of epigenetic changes in the aetiopathogenesis of endometriosis. J. Obstet. Gynaecol. J. Inst. Obstet. Gynaecol. 2016, 36, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Balch, C.; Fang, F.; Matei, D.E.; Huang, T.H.; Nephew, K.P. Minireview: Epigenetic changes in ovarian cancer. Endocrinology 2009, 150, 4003–4011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, M.J.; Shin, Y.K. Epigenetic regulation of cancer-associated genes in ovarian cancer. Int. J. Mol. Sci. 2011, 12, 983–1008. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Jiang, W.; Jiang, Y.; Wang, S.; Fang, J.; Zhu, L.; Zhu, Y.; Yan, G.; Sun, H.; Chen, L.; et al. Preliminary functional inquiry of lncRNA ENST00000433673 in embryo implantation using bioinformatics analysis. Syst. Biol. Reprod. Med. 2019, 65, 164–173. [Google Scholar] [CrossRef]
- Breitling, L.P.; Yang, R.; Korn, B.; Burwinkel, B.; Brenner, H. Tobacco-smoking-related differential DNA methylation: 27K discovery and replication. Am. J. Hum. Genet. 2011, 88, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Sapienza, C.; Issa, J.P. Diet, Nutrition, and Cancer Epigenetics. Annu. Rev. Nutr. 2016, 36, 665–681. [Google Scholar] [CrossRef]
- Dumitrescu, R.G. Early Epigenetic Markers for Precision Medicine. Methods Mol. Biol 2018, 1856, 3–17. [Google Scholar]
- Esfandiari, F.; Favaedi, R.; Heidari-Khoei, H.; Chitsazian, F.; Yari, S.; Piryaei, A.; Ghafari, F.; Baharvand, H.; Shahhoseini, M. Insight into epigenetics of human endometriosis organoids: DNA methylation analysis of HOX genes and their cofactors. Fertil. Steril. 2021, 115, 125–137. [Google Scholar] [CrossRef]
- Kim, T.H.; Yoo, J.Y.; Choi, K.C.; Shin, J.H.; Leach, R.E.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Yoon, H.G.; Jeong, J.W. Loss of HDAC3 results in nonreceptive endometrium and female infertility. Sci. Transl. Med. 2019, 11, eaaf7533. [Google Scholar] [CrossRef] [PubMed]
- Langford, D.J.; West, C.; Elboim, C.; Cooper, B.A.; Abrams, G.; Paul, S.M.; Schmidt, B.L.; Levine, J.D.; Merriman, J.D.; Dhruva, A.; et al. Variations in potassium channel genes are associated with breast pain in women prior to breast cancer surgery. J. Neurogenet. 2014, 28, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Silver, K.; Littlejohn, A.; Thomas, L.; Marsh, E.; Lillich, J.D. Inhibition of Kv channel expression by NSAIDs depolarizes membrane potential and inhibits cell migration by disrupting calpain signaling. Biochem. Pharmacol. 2015, 98, 614–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumawat, K.; Gosens, R. WNT-5A: Signaling and functions in health and disease. Cell Mol. Life Sci. 2016, 73, 567–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozcan, A.; Liles, N.; Coffey, D.; Shen, S.S.; Truong, L.D. PAX2 and PAX8 expression in primary and metastatic mullerian epithelial tumors: A comprehensive comparison. Am. J. Surg. Pathol. 2011, 35, 1837–1847. [Google Scholar] [CrossRef]
- Yu, W.Z.; Chen, X.M.; Niu, W.B.; Wang, F.; Sun, B.; Sun, Y.P. Role of Wnt5a in the differentiation of human embryonic stem cells into endometrium-like cells. Int. J. Clin. Exp. Pathol. 2015, 8, 5478–5484. [Google Scholar] [PubMed]
- Hever, A.; Roth, R.B.; Hevezi, P.A.; Lee, J.; Willhite, D.; White, E.C.; Marin, E.M.; Herrera, R.; Acosta, H.M.; Acosta, A.J.; et al. Molecular characterization of human adenomyosis. Mol. Hum. Reprod. 2006, 12, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Liu, Y.; Tan, X.; Wang, Z.; Li, Y.; Gao, M.; Li, Y.; Fang, Z.; Sun, Y.; Zhang, L.; Wang, X.; et al. Down-regulation of tumor suppressor PDCD4 expression in endometrium of adenomyosis patients. Curr. Res. Transl. Med. 2016, 64, 123–128. [Google Scholar] [CrossRef]
- Kolioulis, I.; Zafrakas, M.; Grimbizis, G.; Miliaras, D.; Timologou, A.; Bontis, J.N.; Tarlatzis, B.C. Immunohistochemical expression pattern of metastasis suppressor KISS-1 protein in adenomyosis lesions and normal endometrium. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 210, 64–68. [Google Scholar] [CrossRef]
- Patel, S.K.; Jackson, L.; Warren, A.Y.; Arya, P.; Shaw, R.W.; Khan, R.N. A role for two-pore potassium (K2P) channels in endometrial epithelial function. J. Cell Mol. Med. 2013, 17, 134–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Luo, L.; Lal, B.; Ma, X.; Chen, L.; Hann, C.L.; Fulton, A.M.; Leahy, D.J.; Laterra, J.; Li, M. A monoclonal antibody against KCNK9 K(+) channel extracellular domain inhibits tumour growth and metastasis. Nat. Commun. 2016, 7, 10339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herndon, C.N.; Aghajanova, L.; Balayan, S.; Erikson, D.; Barragan, F.; Goldfien, G.; Vo, K.C.; Hawkins, S.; Giudice, L.C. Global Transcriptome Abnormalities of the Eutopic Endometrium from Women with Adenomyosis. Reprod. Sci. 2016, 23, 1289–1303. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, L.-H.; Liao, C.-C.; Liew, P.-L.; Chen, C.-W.; Su, P.-H.; Wen, K.-C.; Lai, H.-C.; Huang, R.-L.; Chen, L.-Y. Epigenomic Analysis Reveals the KCNK9 Potassium Channel as a Potential Therapeutic Target for Adenomyosis. Int. J. Mol. Sci. 2022, 23, 5973. https://doi.org/10.3390/ijms23115973
Chu L-H, Liao C-C, Liew P-L, Chen C-W, Su P-H, Wen K-C, Lai H-C, Huang R-L, Chen L-Y. Epigenomic Analysis Reveals the KCNK9 Potassium Channel as a Potential Therapeutic Target for Adenomyosis. International Journal of Molecular Sciences. 2022; 23(11):5973. https://doi.org/10.3390/ijms23115973
Chicago/Turabian StyleChu, Ling-Hui, Chi-Chun Liao, Phui-Ly Liew, Chien-Wen Chen, Po-Hsuan Su, Kuo-Chang Wen, Hung-Cheng Lai, Rui-Lan Huang, and Lin-Yu Chen. 2022. "Epigenomic Analysis Reveals the KCNK9 Potassium Channel as a Potential Therapeutic Target for Adenomyosis" International Journal of Molecular Sciences 23, no. 11: 5973. https://doi.org/10.3390/ijms23115973
APA StyleChu, L. -H., Liao, C. -C., Liew, P. -L., Chen, C. -W., Su, P. -H., Wen, K. -C., Lai, H. -C., Huang, R. -L., & Chen, L. -Y. (2022). Epigenomic Analysis Reveals the KCNK9 Potassium Channel as a Potential Therapeutic Target for Adenomyosis. International Journal of Molecular Sciences, 23(11), 5973. https://doi.org/10.3390/ijms23115973