
Transcription Factor SrsR (YgfI) Is a Novel Regulator for the Stress-Response Genes in Stationary Phase in Escherichia coli K-12


 and
and
Abstract
:1. Introduction
2. Results
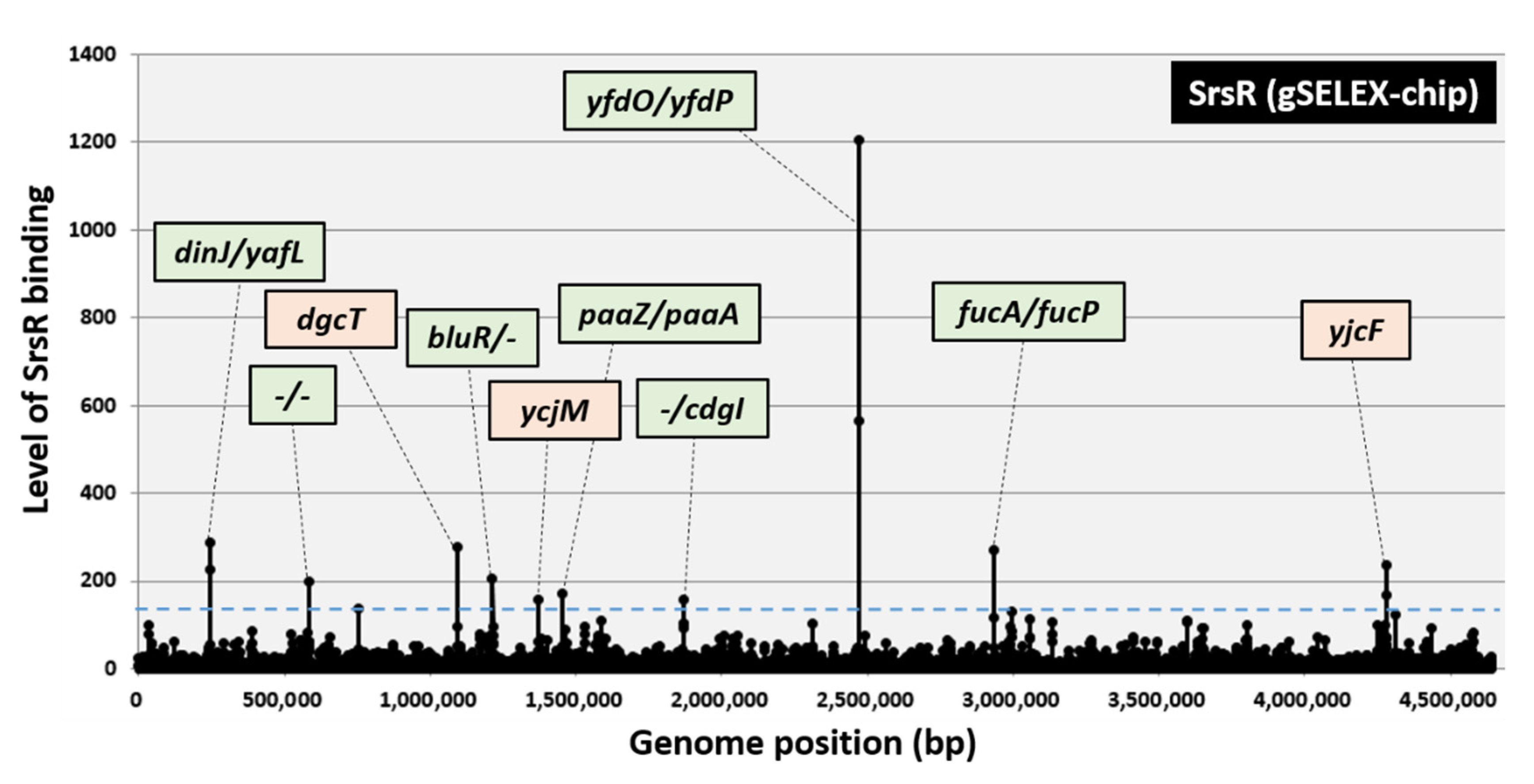
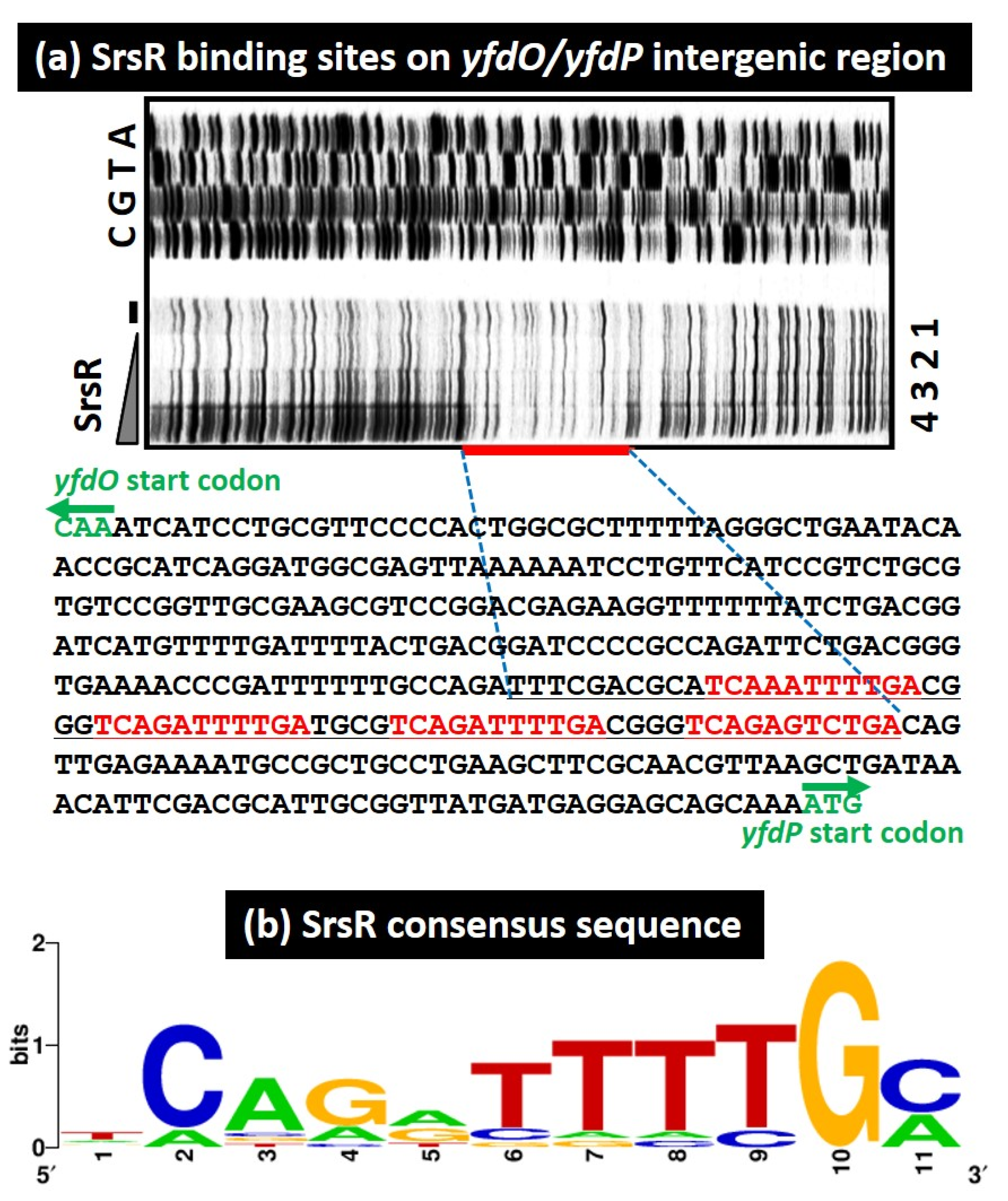
2.1. Regulation Targets of SrsR per gSELEX-Chip Screening
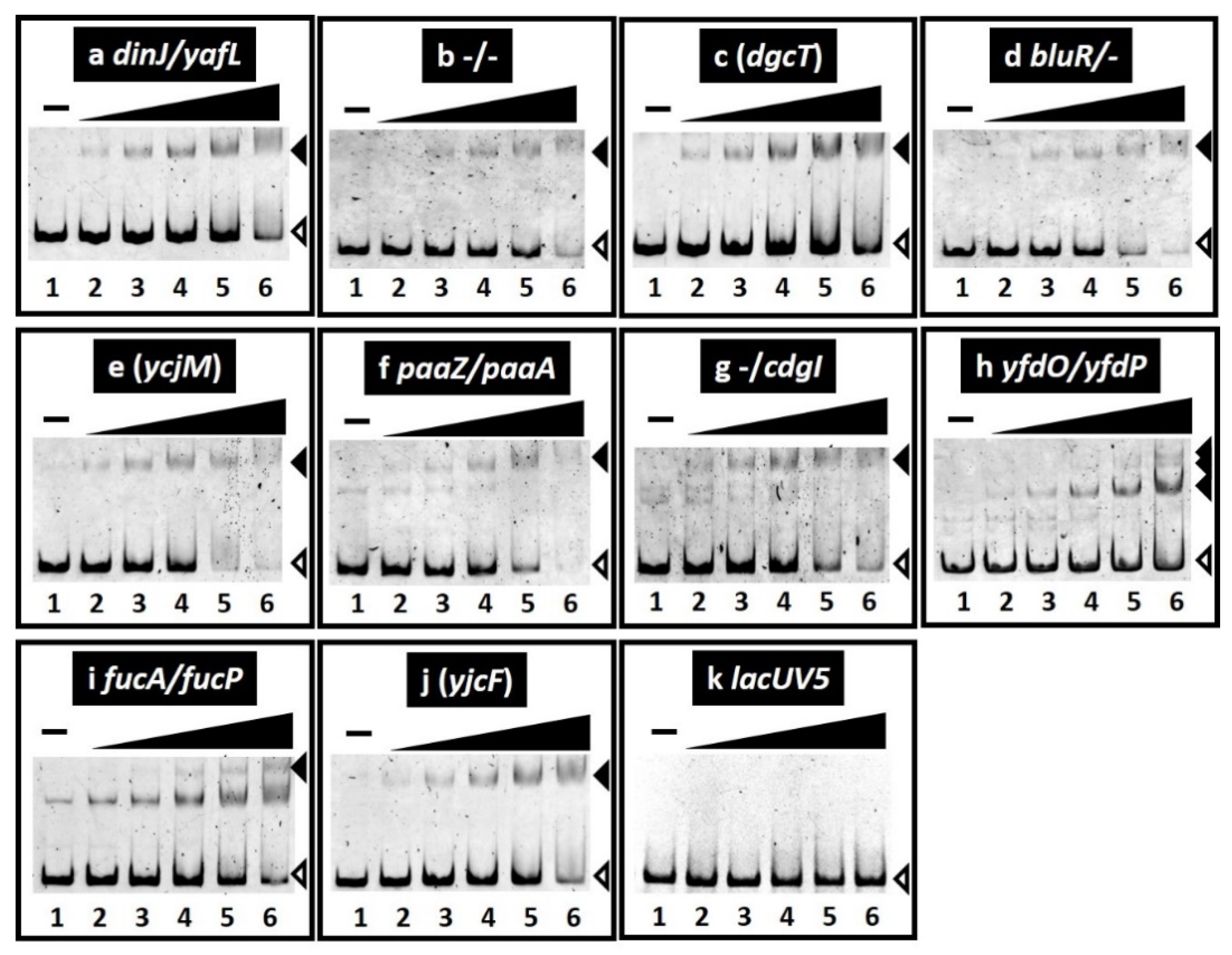
2.2. Binding In Vitro of SrsR to the Target Sequences
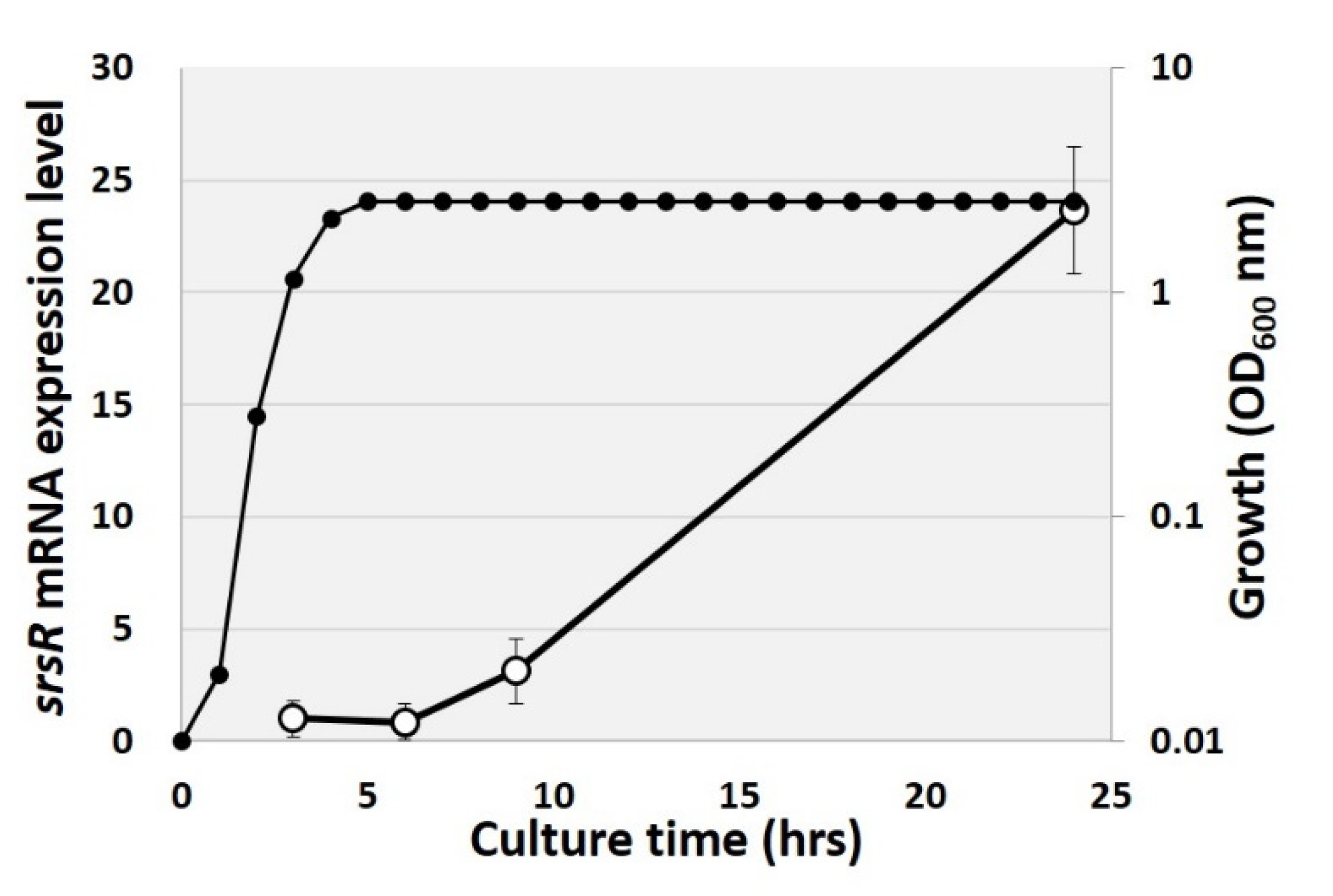
2.3. Growth-Dependent Expression Level of srsR Gene
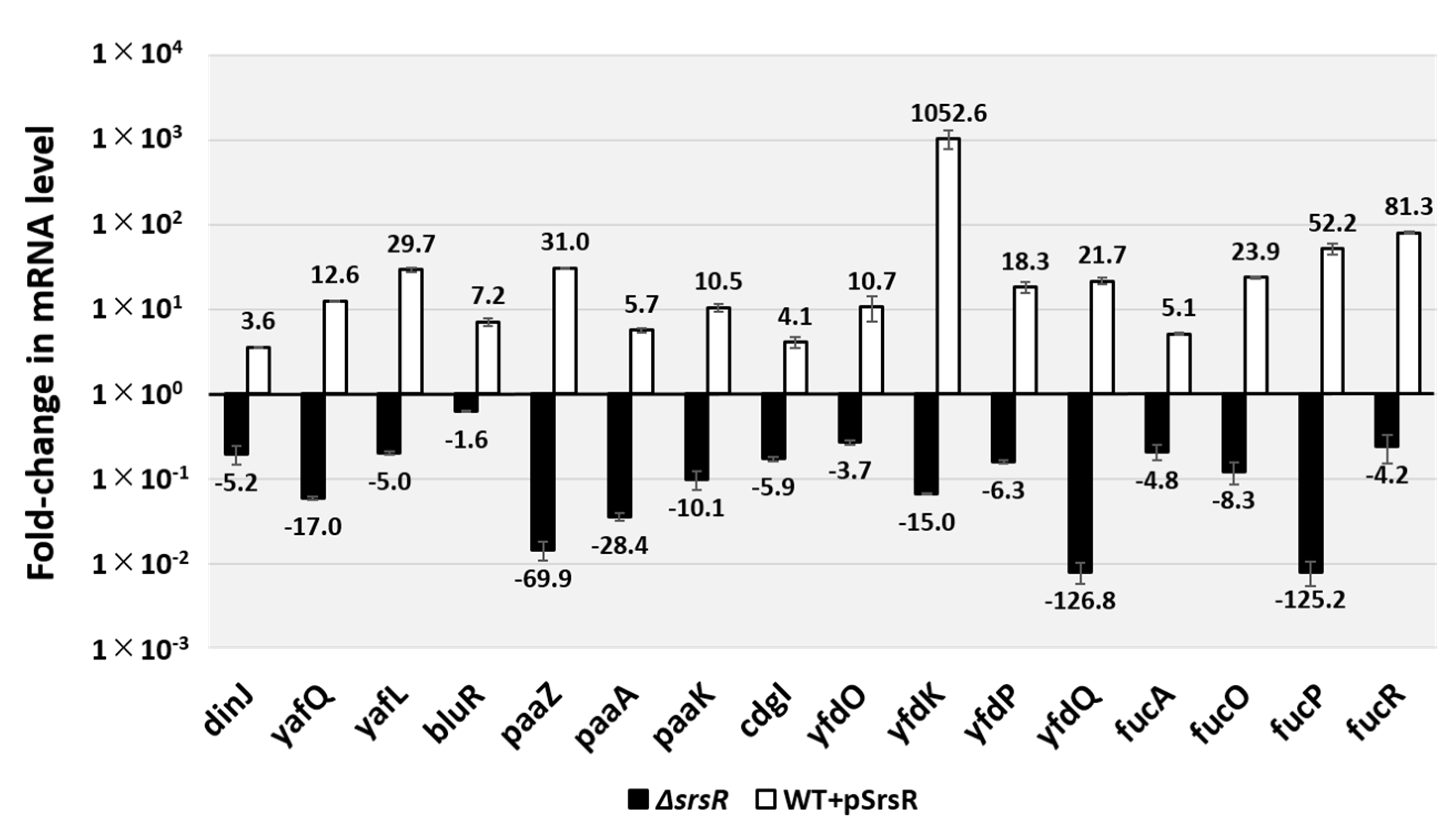
2.4. Regulatory Role of Srsr in Expression of the Target Genes In Vivo
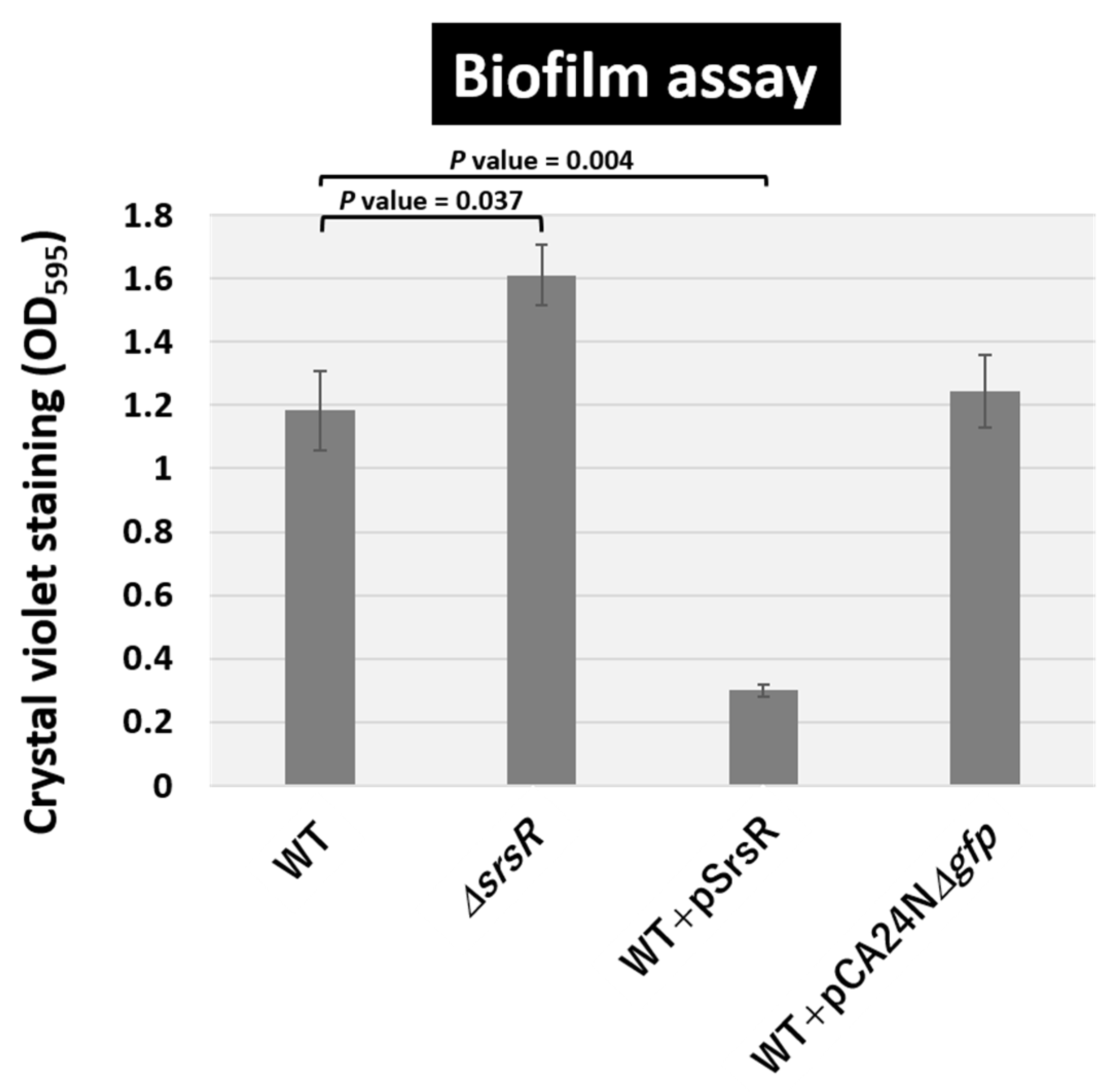
2.5. Regulatory Role of SrsR in Biofilm Formation
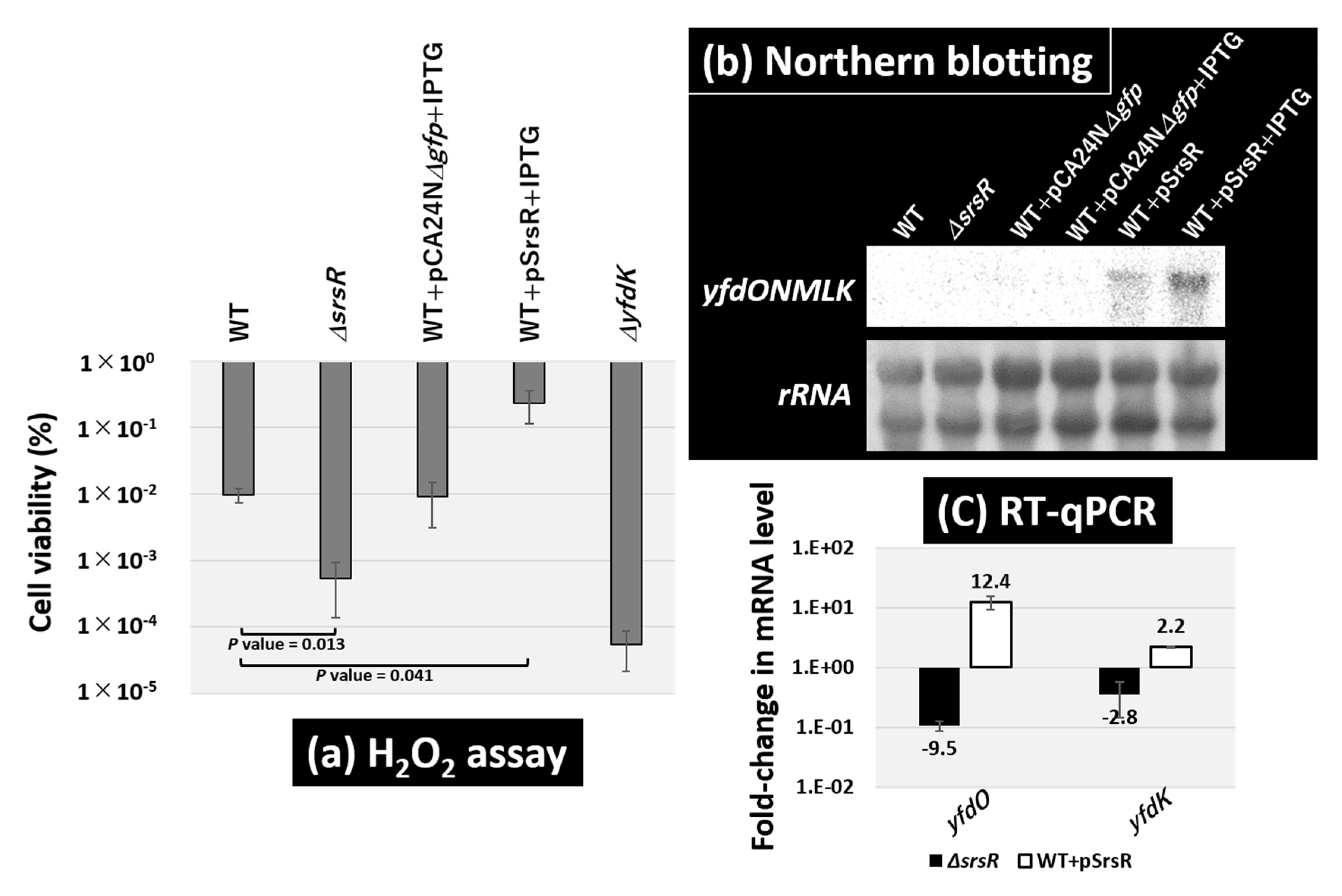
2.6. Regulatory Role of the SrsR in Hydrogen Peroxide Sensitivity
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.2. Purification of SrsR Protein
4.3. Genomic SELEX (gSELEX) Screening of SrsR-Binding Sequences
4.4. Gel Shift Assay
4.5. DNase-I Footprinting Assay
4.6. Consensus Sequence Analysis
4.7. RT-qPCR Analysis
4.8. Biofilm Assay
4.9. Hydrogen Peroxide Test
4.10. Northern Blot Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ishihama, A. Adaptation of gene expression in stationary phase bacteria. Curr. Opin. Genet. Dev. 1997, 7, 582–588. [Google Scholar] [CrossRef]
- Ishihama, A. Modulation of the nucleoid, the transcription apparatus, and the translation machinery in bacteria for stationary phase survival. Genes Cells 1999, 4, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Hengge, R. Stationary-phage gene regulation in Escherichia coli. EcoSal Plus 2011, 4, ecosalplus5.6.3. [Google Scholar] [CrossRef] [PubMed]
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The complete genome sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Morooka, N.; Yamamoto, Y.; Fujita, K.; Isono, K.; Choi, S.; Ohtsubo, E.; Baba, T.; Wanner, B.L.; Mori, H.; et al. Highly accurate genome sequences of Escherichia coli K-12 strains MG1655 and W3110. Mol. Syst. Biol. 2006, 2, 2006.0007. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kim, Y.; Ma, Q.; Hong, S.H.; Pokusaeva, K.; Sturino, J.M.; Wood, T.K. Cryptic prophages help bacteria cope with adverse environments. Nat. Commun. 2010, 1, 147. [Google Scholar] [CrossRef] [Green Version]
- Soo, V.W.C.; Hanson-Manful, P.; Patrick, W.M. Artificial gene amplification reveals an abundance of promiscuous resistance determinants in Escherichia coli. Proc. Natl. Acad. Sci. USA 2011, 108, 1484–1489. [Google Scholar] [CrossRef] [Green Version]
- Ishihama, A.; Kori, A.; Koshio, E.; Yamada, K.; Maeda, H.; Shimada, T.; Makinoshima, H.; Iwata, A.; Fujita, N. Intracellular concentrations of 65 species of transcription factors with known regulatory functions in Escherichia coli. J. Bacteriol. 2014, 196, 2718–2727. [Google Scholar] [CrossRef] [Green Version]
- Ishihama, A.; Shimada, T.; Yamazaki, Y. Transcription profile of Escherichia coli: Genomic SELEX search for regulatory targets of transcription factors. Nucleic Acids Res. 2016, 44, 2058–2074. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Fujita, N.; Maeda, M.; Ishihama, A. Systematic search for the Cra-binding promoters using genomic SELEX system. Genes Cells 2005, 10, 907–918. [Google Scholar] [CrossRef]
- Shimada, T.; Ogasawara, H.; Ishihama, A. Genomic SELEX screening of regulatory targets of Escherichia coli transcription factors. Methods Mol. Biol. 2018, 1837, 49–69. [Google Scholar] [PubMed]
- Ishihama, A. Prokaryotic genome regulation: A revolutionary paradigm. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 485–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, T.; Ogasawara, H.; Ishihama, A. Single-target regulators form a minor group of transcription factors in Escherichia coli K-12. Nucleic Acids Res. 2018, 46, 3921–3936. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Yamamoto, K.; Nakano, M.; Watanabe, H.; Schleheck, D.; Ishihama, A. Regulatory role of CsqR (YihW) in transcription of the genes for catabolism of the anionic sugar sulfoquinovose (SQ) in Escherichia coli K-12. Microbiology 2019, 165, 78–89. [Google Scholar] [CrossRef]
- Shimada, T.; Yokoyama, Y.; Anzai, T.; Yamamoto, K.; Ishihama, A. Regulatory role of PlaR (YiaJ) for plant utilization in Escherichia coli K-12. Sci. Rep. 2019, 9, 20415. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Ogasawara, H.; Kobayashi, I.; Kobayashi, N.; Ishihama, A. Single-target regulators constitute the minority group of transcription factors in Escherichia coli K-12. Front. Microbiol. 2021, 12, 697803. [Google Scholar] [CrossRef]
- Shimada, T.; Murayama, R.; Mashima, T.; Kawano, N.; Ishihama, A. Regulatory role of CsuR (YiaU) in determination of cell surface properties of Escherichia coli K-12. Microbiology 2022, in press. [CrossRef]
- Ogasawara, H.; Yamada, K.; Kori, A.; Yamamoto, K.; Ishihama, A. Regulation of the Escherichia coli csgD promoter: Interplay between five transcription factors. Microbiology 2010, 156, 2470–2483. [Google Scholar] [CrossRef] [Green Version]
- Teramoto, J.; Yoshimura, S.H.; Takeyasu, K.; Ishihama, A. A novel nucleoid protein of Escherichia coli induced under anaerobiotic growth conditions. Nucleic Acids Res. 2010, 38, 3605–3618. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Bridier, A.; Briandet, R.; Ishihama, A. Novel roles of LeuO in transcription regulation of E. coli genome: Antagonistic interplay with the universal silencer H-NS. Mol. Microbiol. 2011, 82, 378–397. [Google Scholar] [CrossRef]
- Ishihama, A. Prokaryotic genome regulation: Multifactor promoters, multitarget regulators and hierarchic networks. FEMS Microbiol. Rev. 2010, 34, 628–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.J.; Wade, W.D.; Akierman, S.; Vacchi-Suzzi, C.; Stremick, C.A.; Turner, R.J.; Ceri, H. The chromosomal toxin gene yafQ is a determinant of multidrug tolerance for Escherichia coli growing in a biofilm. Antimicrob. Agents Chemother. 2009, 53, 2253–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschowri, N.; Busse, S.; Hengge, R. The BLUF-EAL protein YcgF acts as a direct anti-repressor in a blue-light response of Escherichia coli. Genes Dev. 2009, 23, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Sommerfeldt, N.; Possling, A.; Becker, G.; Pesavento, C.; Tschowri, N.; Hengge, R. Gene expression patterns and differential input into curli fimbriae regulation of all GGDEF/EAL domain proteins in Escherichia coli. Microbiology 2009, 155, 1318–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Ruf, C.; Taddei, F.; Matic, I. Stress and survival of aging Escherichia coli rpoS colonies. Genetics 2004, 168, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lee, J.; Page, R.; Garcia-Contreras, R.; Palermino, J.M.; Zhang, X.S.; Doshi, O.; Wood, T.K.; Peti, W. Structure and function of the E. coli protein YmgB: A protein critical for biofilm formation and acid-resistance. J. Mol. Biol. 2007, 373, 11–26. [Google Scholar] [CrossRef]
- Domka, J.; Lee, J.; Bansal, T.; Wood, T.K. Temporal gene-expression in Escherichia coli K-12 biofilms. Environ. Microbiol. 2007, 9, 332–346. [Google Scholar] [CrossRef]
- Sanchez-Torres, V.; Hu, H.; Wood, T.K. GGDEF proteins YeaI, YedQ, and YfiN reduce early biofilm formation and swimming motility in Escherichia coli. Appl. Microbiol. Biotechnol. 2011, 90, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Autieri, S.M.; Lins, J.J.; Leatham, M.P.; Laux, D.C.; Conway, T.; Cohen, P.S. L-fucose stimulates utilization of D-ribose by Escherichia coli MG1655 ΔfucAO and E. coli Nissle 1917 ΔfucAO mutants in the mouse intestine and in M9 minimal medium. Infect. Immun. 2007, 75, 5465–5475. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.E.; Smalley, D.J.; Tucker, D.L.; Leatham, M.P.; Norris, W.E.; Stevenson, S.J.; Anderson, A.B.; Grissom, J.E.; Laux, D.C.; Cohen, P.S.; et al. Carbon nutrition of Escherichia coli in the mouse intestine. Proc. Natl. Acad. Sci. USA 2004, 101, 7427–7432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, T.; Katayama, Y.; Kawakita, S.; Ogasawara, H.; Nakano, M.; Yamamoto, K.; Ishihama, A. A novel regulator RcdA of the csgD gene encoding the master regulator of biofilm formation in Escherichia coli. MicrobiologyOpen 2012, 1, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Karatan, E.; Watnick, P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, T.K. Insights on Escherichia coli biofilm formation and inhibition from whole-transcriptome profiling. Environ. Microbiol. 2009, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Sharma, S.; Sharma, P.; Chandola, D.; Dang, S.; Gupta, S.; Gabrani, R. Escherichia coli biofilm: Development and therapeutic strategies. J. Appl. Microbiol. 2016, 121, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, H.; Ishizuka, T.; Hotta, S.; Aoki, M.; Shimada, T.; Ishihama, A. Novel regulators of the csgD gene encoding the master regulator of biofilm formation in Escherichia coli K-12. Microbiology 2020, 166, 880–890. [Google Scholar] [CrossRef]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef]
- Barraud, N.; Letoffe, S.; Beloin, C.; Vinh, J.; Chiappetta, G.; Ghigo, J.M. Lifestyle-specific S-nitrosylation of protein cysteine thiols regulates Escherichia coli biofilm formation and resistance to oxidative stress. NPJ Biofilms Microbiomes 2021, 7, 34. [Google Scholar] [CrossRef]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef] [Green Version]
- Aquino, P.; Honda, B.; Jaini, S.; Lyubetskaya, A.; Hosur, K.; Chiu, J.G.; Ekladious, I.; Hu, D.; Jin, L.; Sayeg, M.K.; et al. Coordinated regulation of acid resistance in Escherichia coli. BMC Syst. Biol. 2017, 11, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jishage, M.; Ishihama, A. Variation in RNA polymerase sigma subunit composition within different stocks of Escherichia coli W3110. J. Bacteriol. 1997, 179, 959–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Ishihama, A.; Busby, S.J.W.; Grainger, D.C. The Escherichia coli RutR transcription factor binds at targets within genes as well as intergenic regions. Nucleic Acids Res. 2008, 36, 3950–3955. [Google Scholar] [CrossRef]
- Shimada, T.; Tanaka, K.; Ishihama, A. Transcription factor DecR (YbaO) controls detoxification of L-cysteine in Escherichia coli. Microbiology 2016, 162, 1698–1707. [Google Scholar] [CrossRef]
- Shimada, T.; Hirao, K.; Kori, A.; Yamamoto, K.; Ishihama, A. RutR is the uracil/thymine-sensing master regulator of a set of genes for synthesis and degradation of pyrimidines. Mol. Micribiol. 2007, 66, 744–757. [Google Scholar] [CrossRef]
- Anzai, T.; Imamura, S.; Ishihama, A.; Shimada, T. Expanded roles of pyruvate-sensing PdhR in transcription regulation of the Escherichia coli K-12 genome: Fatty acid catabolism and cell motility. Microb. Genom. 2020, 6, mgen000442. [Google Scholar] [CrossRef]
- Shimada, T.; Saito, N.; Maeda, M.; Tanaka, K.; Ishihama, A. Expanded roles of leucine-responsive regulatory protein in transcription regulation of the Escherichia coli genome: Genomic SELEX screening of the regulation targets. Microb. Genom. 2015, 1, e000001. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Position (bp) | Intensity | Function | Operon | Gene | D | SrsR Site | D | Gene | Operon | Function | SrsR Box TCAGATTTTGC | Conservation | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 246,530 | 288 | Antitoxin/DNA-binding transcriptional repressor | dinJ-yafQ | dinJ | < | > | yafL | yafL | predicted lipoprotein and C40 family peptidase | aaAGgcaTTGC | 6/11 | |
| 2 | 583,766 | 200 | appY | > | < | ompT | TCcatTTTTGC | 8/11 | |||||
| 3 | 1,092,430 | 278 | pgaA | < | dgcT | < | insEF | cCAGgcgTTGa | 6/11 | ||||
| 4 | 1,213,450 | 207 | Transcriptional repressor | bluR | bluR | < | < | ycgF | gCAGgTTTTGC | 9/11 | |||
| 5 | 1,368,672 | 158 | pspE | > | ycjM | > | ycjN | TCActTTTTGC | 9/11 | ||||
| 6 | 1,451,756 | 170 | Oxepin-CoA hydrolase/3-oxo-5,6-dehydrosuberyl-CoA semialdehyde dehydrogenase | paaZ | paaZ | < | > | paaA | paaABCDEFGHIJK | predicted ring 1,2-phenylacetyl-CoA epoxidase subunit | aCgGATTTcGC | 8/11 | |
| 7 | 1,868,366 | 158 | yeaH | > | > | cdgI | cdgI | c-di-GMP binding protein | aaAGgTTaTGC | 7/11 | |||
| 8 | 2,471,464 | 1206 | CPS-53 (KpLE1) prophage; predicted defective phage replication protein O | yfdONMLK | yfdO | < | > | yfdP | yfdPQ | CPS-53 (KpLE1) prophage; predicted protein | TCAaATTTTGa, TCAGATTTTGa, TCAGATTTTGa, TCAGAgTcTGa | 9/11, 10/11, 10/11, 8/11 | |
| 9 | 2,931,960 | 270 | L-fuculose-phosphate aldolase | fucAO | fucA | < | > | fucP | fucPIKUR | FucP fucose MFS transporter | gCtGgTTTTGC | 8/11 | |
| 10 | 4,280,360 | 235 | yjcE | > | yjcF | < | actP | TCAaATTTcGC | 9/11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, I.; Mochizuki, K.; Teramoto, J.; Imamura, S.; Takaya, K.; Ishihama, A.; Shimada, T. Transcription Factor SrsR (YgfI) Is a Novel Regulator for the Stress-Response Genes in Stationary Phase in Escherichia coli K-12. Int. J. Mol. Sci. 2022, 23, 6055. https://doi.org/10.3390/ijms23116055
Kobayashi I, Mochizuki K, Teramoto J, Imamura S, Takaya K, Ishihama A, Shimada T. Transcription Factor SrsR (YgfI) Is a Novel Regulator for the Stress-Response Genes in Stationary Phase in Escherichia coli K-12. International Journal of Molecular Sciences. 2022; 23(11):6055. https://doi.org/10.3390/ijms23116055
Chicago/Turabian StyleKobayashi, Ikki, Kenji Mochizuki, Jun Teramoto, Sousuke Imamura, Kazuhiro Takaya, Akira Ishihama, and Tomohiro Shimada. 2022. "Transcription Factor SrsR (YgfI) Is a Novel Regulator for the Stress-Response Genes in Stationary Phase in Escherichia coli K-12" International Journal of Molecular Sciences 23, no. 11: 6055. https://doi.org/10.3390/ijms23116055
APA StyleKobayashi, I., Mochizuki, K., Teramoto, J., Imamura, S., Takaya, K., Ishihama, A., & Shimada, T. (2022). Transcription Factor SrsR (YgfI) Is a Novel Regulator for the Stress-Response Genes in Stationary Phase in Escherichia coli K-12. International Journal of Molecular Sciences, 23(11), 6055. https://doi.org/10.3390/ijms23116055