
RadA, a Key Gene of the Circadian Rhythm of Escherichia coli

 ,
, 
 , and
, and
Abstract
:1. Introduction
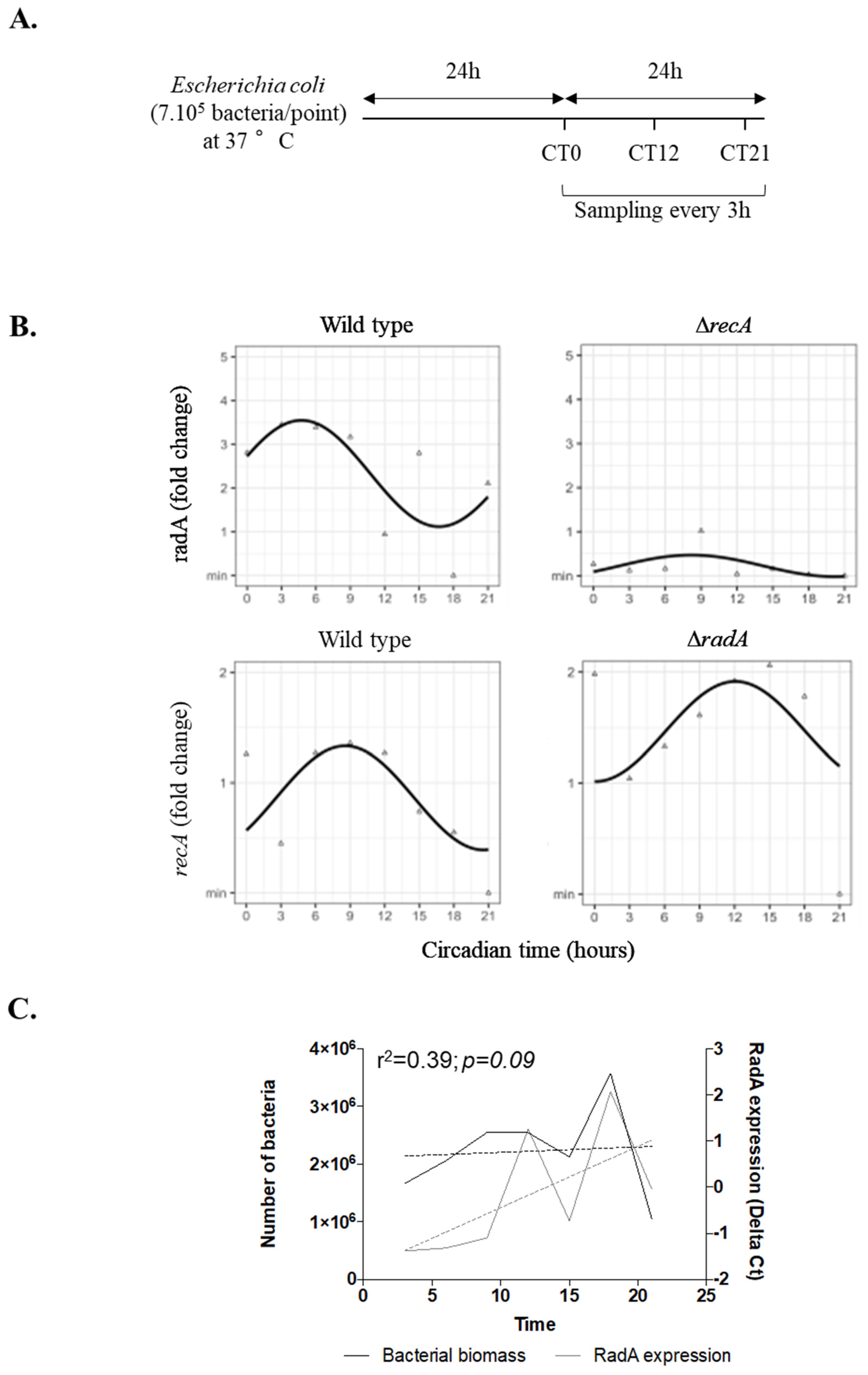
2. Results and Discussion
2.1. RadA Is a KaiC Homologue in Escherichia Coli
2.2. RadA and recA Genes Present a Circadian Rhythm in Escherichia Coli
3. Materials and Methods
3.1. Protein Sequence Analysis
3.2. Bacterial Culture
3.3. Bacterial RNA Extraction and Real-Time Quantitative PCR (q-RTPCR)
3.4. Bacterial DNA Extraction and qPCR
3.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, J.C.; Rosbash, M.; Young, M.W. Nobelprize.org 2017 The Nobel Prize in Physiology or Medicine 2017. Ukr. Biochem. J. 2017, 89, 5. [Google Scholar]
- Diallo, A.B.; Coiffard, B.; Leone, M.; Mezouar, S.; Mege, J.L. For Whom the Clock Ticks: Clinical Chronobiology for Infectious Diseases. Front. Immunol. 2020, 11, 1457. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.H.; Takahashi, J.S. Molecular components of the mammalian circadian clock. Hum. Mol. Genet. 2006, 15, R271–R277. [Google Scholar] [CrossRef] [PubMed]
- Mallet De Lima, C.D.; Göndör, A. Circadian organization of the genome. Science 2018, 359, 1212–1213. [Google Scholar] [CrossRef]
- Swan, J.A.; Golden, S.S.; Li, W.A.; Partch, C.L. Structure, function, and mechanism of the core circadian clock in cyanobacteria. J. Biol. Chem. 2018, 293, 5026–5034. [Google Scholar] [CrossRef] [Green Version]
- Ishiura, M.; Kutsuna, S.; Aoki, S.; Iwasaki, H.; Andersson, C.R.; Tanabe, A.; Golden, S.S.; Johnson, C.H.; Kondo, T. Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science 1998, 281, 1519–1523. [Google Scholar] [CrossRef]
- Cohen, S.E.; Golden, S.S. Circadian Rhythms in Cyanobacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Brettschneider, C.; Rose, R.J.; Hertel, S.; Axmann, I.M.; Heck, A.J.R.; Kollmann, M. A sequestration feedback determines dynamics and temperature entrainment of the KaiABC circadian clock. Mol. Syst. Biol. 2010, 6, 389. [Google Scholar] [CrossRef]
- Schmelling, N.M.; Lehmann, R.; Chaudhury, P.; Beck, C.; Albers, S.V.; Axmann, I.M.; Wiegard, A. Minimal tool set for a prokaryotic circadian clock. BMC Evol. Biol. 2017, 17, 169. [Google Scholar] [CrossRef] [Green Version]
- Paulose, J.K.; Cassone, C.V.; Graniczkowska, K.B.; Cassone, V.M. Entrainment of the Circadian Clock of the Enteric Bacterium Klebsiella aerogenes by Temperature Cycles. iScience 2019, 19, 1202–1213. [Google Scholar] [CrossRef]
- Eelderink-Chen, Z.; Bosman, J.; Sartor, F.; Dodd, A.N.; Kovács, Á.T.; Merrow, M. A circadian clock in a nonphotosynthetic prokaryote. Sci. Adv. 2021, 7, eabe2086. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.H.; Lubkowicz, D.; Yeong, V.; Chang, R.L.; Silver, P.A. Transplantability of a circadian clock to a noncircadian organism. Sci. Adv. 2015, 1, e1500358. [Google Scholar] [CrossRef] [PubMed]
- Coiffard, B.; Diallo, A.B.; Culver, A.; Mezouar, S.; Hammad, E.; Vigne, C.; Nicolino-Brunet, C.; Dignat-George, F.; Baumstarck, K.; Boucekine, M.; et al. Circadian Rhythm Disruption and Sepsis in Severe Trauma Patients. SHOCK 2018, 52, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2017, 27, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.L.; Lovett, S.T. Recombinational branch migration by the RadA/Sms paralog of RecA in Escherichia coli. eLife 2016, 5, e10807. [Google Scholar] [CrossRef]
- Haldenby, S.; White, M.F.; Allers, T. RecA family proteins in archaea: RadA and its cousins. Biochem. Soc. Trans. 2009, 37, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Galperin, M.Y.; Koonin, E.V. Proposed role for KaiC-like ATPases as major signal transduction hubs in archaea. mBio 2017, 8, e01959-17. [Google Scholar] [CrossRef] [Green Version]
- Marie, L.; Rapisarda, C.; Morales, V.; Bergé, M.; Perry, T.; Soulet, A.L.; Gruget, C.; Remaut, H.; Fronzes, R.; Polard, P. Bacterial RadA is a DnaB-Type helicase interacting with RecA to promote bidirectional D-loop extension. Nat. Commun. 2017, 8, 15638. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Zhou, L.; Lim, Q.; Zou, R.; Stephanopoulos, G.; Too, H.P. Novel reference genes for quantifying transcriptional responses of Escherichia coli to protein overexpression by quantitative PCR. BMC Mol. Biol. 2011, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Golden, S.S.; Canales, S.R. Cyanobacterial circadian clocks-timing is everything. Nat. Rev. Microbiol. 2003, 1, 191–199. [Google Scholar] [CrossRef]
- Kovač, U.; Žužek, Z.; Raspor Dall’Olio, L.; Pohar, K.; Ihan, A.; Moškon, M.; Rozman, D.; Starčič Erjavec, M. Escherichia coli Affects Expression of Circadian Clock Genes in Human Hepatoma Cells. Microorganisms 2021, 9, 869. [Google Scholar] [CrossRef] [PubMed]
- Kaluzna, M.; Kuras, A.; Pulawska, J. Validation of reference genes for the normalization of the RT-qPCR gene expression of virulence genes of Erwinia amylovora in apple shoots. Sci. Rep. 2017, 7, 2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Gene | MESOR | CI 95% | p-Value | Amplitude | CI 95% | p-Value | Acrophase | CI 95% | p-Value | Significant | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Wild type | radA | 2.33 | (1.63; 3.02) | <0.001 | 1.21 | (0.23; 2.20) | 0.01 | 4.70 | (1.64; 7.83) | 0.003 | Yes |
| recA | 0.86 | (0.57; 1.15) | <0.001 | 0.47 | (0.06; 0.88) | 0.02 | 8.57 | (5.25; 11.88) | 0.04 | Yes | |
| ihfB | 2.10 | (1.38; 2.83) | <0.001 | 1.09 | (0.07; 2.11) | 0.03 | 2.10 | (0.00; 5.65) | 0.24 | No | |
| cysG | 1.72 | (1.07; 2.37) | <0.001 | 1.49 | (0.57; 2.40) | 0.001 | 4.20 | (1.83; 6.5) | <0.001 | Yes | |
| rrsA | 2.05 | (1.18; 2.91) | <0.001 | 1.59 | (0.37; 2.81) | 0.01 | 21.32 | (18.4; 24.2) | 0.07 | No | |
| ssrA | 3.98 | (2.75; 5.21) | <0.001 | 3.84 | (2.10; 5.58) | <0.001 | 16.85 | (15.1; 18.6) | <0.001 | Yes | |
| ΔradA | radA | - | - | - | - | - | - | - | - | - | |
| recA | 1.36 | (1; 1.68) | <0.001 | 0.45 | (-0.24; 1.14) | 0.005 | 12.1 | (6.21; 17.9) | 0.98 | No | |
| ihfB | 1.12 | (0.56; 2.92) | <0.001 | 0.42 | (-0.37; 1.21) | 0.30 | 19.6 | (12.5; 21.2) | 0.23 | No | |
| cysG | 0.32 | (0.17; 0.45) | <0.001 | 0.20 | (0.00; 0.39) | 0.05 | 19.2 | (15.4; 22.9) | <0.001 | No | |
| rrsA | 0.57 | (0.17; 0.97) | 0.005 | 0.38 | (-0.18; 0.94) | 0.19 | 0.91 | (19.2; 6.60) | 0.75 | No | |
| ssrA | 1.39 | (0.94; 1.84) | <0.001 | 0.83 | (0.20; 1.47) | 0.01 | 0.72 | (21.8; 3.64) | 0.62 | No | |
| ΔrecA | radA | 0.22 | (0.00; 0.45) | 0.05 | 0.24 | (-0.08; 0.57) | 0.14 | 8.18 | (3.17; 13.2) | 0.14 | No |
| recA | - | - | - | - | - | - | - | - | - | ||
| ihfB | 0.29 | (0.09; 0.50) | 0.005 | 0.16 | (-0.12; 0.45) | 0.27 | 10.3 | (3.58; 17.0) | 0.62 | No | |
| cysG | 1.03 | (0.58; 1.48) | <0.001 | 0.32 | (-0.31; 0.96) | 0.32 | 8.14 | (0.67; 15.6) | 0.31 | No | |
| rrsA | 0.96 | (0.69; 1.23) | <0.001 | 0.63 | (0.25; 1.00) | 0.001 | 7.02 | (4.70; 9.33) | <0.001 | Yes | |
| ssrA | 1.12 | (0.80; 1.45) | <0.001 | 0.80 | (0.33; 1.27) | <0.001 | 6.66 | (4.45; 8.86) | <0.001 | Yes |
| Gene Symbol | Forward Primer (5′-3′) | Reverse Primer (3′-5′) |
|---|---|---|
| rpoB | GTTTCACCACCATCCCACATTC | TTCGGCGTTACCTTACCAAC |
| radA | GTGATGGTGGTATGGGAAGG | GCTAAGTCGGCACTGGTTTC |
| recA | ATTCTACGCCTCTGTTCGTCTC | GCATTCGCTTTACCCTGACC |
| rrsA | CTTACGACCAGGGCTACACA | CTTGTTACGACTTCACCCCAGT |
| ssrA | GTTACACATTGGGGCTGATTCT | CTTTTGGGTTTGACCTCTCTTG |
| ihfB | GGTTTCGGCAGTTTCTCTTTG | CCAGTTCTACTTTATCGCCAGTC |
| cysG | AGCGTTTATTCCACAGTTCACC | CGTTACAGAAGATGCGACGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diallo, A.B.; Mezouar, S.; Boumaza, A.; Fiammingo, O.; Coiffard, B.; Pontarotti, P.; Desnues, B.; Mege, J.-L. RadA, a Key Gene of the Circadian Rhythm of Escherichia coli. Int. J. Mol. Sci. 2022, 23, 6136. https://doi.org/10.3390/ijms23116136
Diallo AB, Mezouar S, Boumaza A, Fiammingo O, Coiffard B, Pontarotti P, Desnues B, Mege J-L. RadA, a Key Gene of the Circadian Rhythm of Escherichia coli. International Journal of Molecular Sciences. 2022; 23(11):6136. https://doi.org/10.3390/ijms23116136
Chicago/Turabian StyleDiallo, Aissatou Bailo, Soraya Mezouar, Asma Boumaza, Oksana Fiammingo, Benjamin Coiffard, Pierre Pontarotti, Benoit Desnues, and Jean-Louis Mege. 2022. "RadA, a Key Gene of the Circadian Rhythm of Escherichia coli" International Journal of Molecular Sciences 23, no. 11: 6136. https://doi.org/10.3390/ijms23116136
APA StyleDiallo, A. B., Mezouar, S., Boumaza, A., Fiammingo, O., Coiffard, B., Pontarotti, P., Desnues, B., & Mege, J.-L. (2022). RadA, a Key Gene of the Circadian Rhythm of Escherichia coli. International Journal of Molecular Sciences, 23(11), 6136. https://doi.org/10.3390/ijms23116136