
Differential Roles of Three α-Crystallin Domain-Containing sHsps of Beauveria bassiana in Asexual Development, Multiple Stress Tolerance and Virulence




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bioinformatic Features of sHsps in B. bassiana
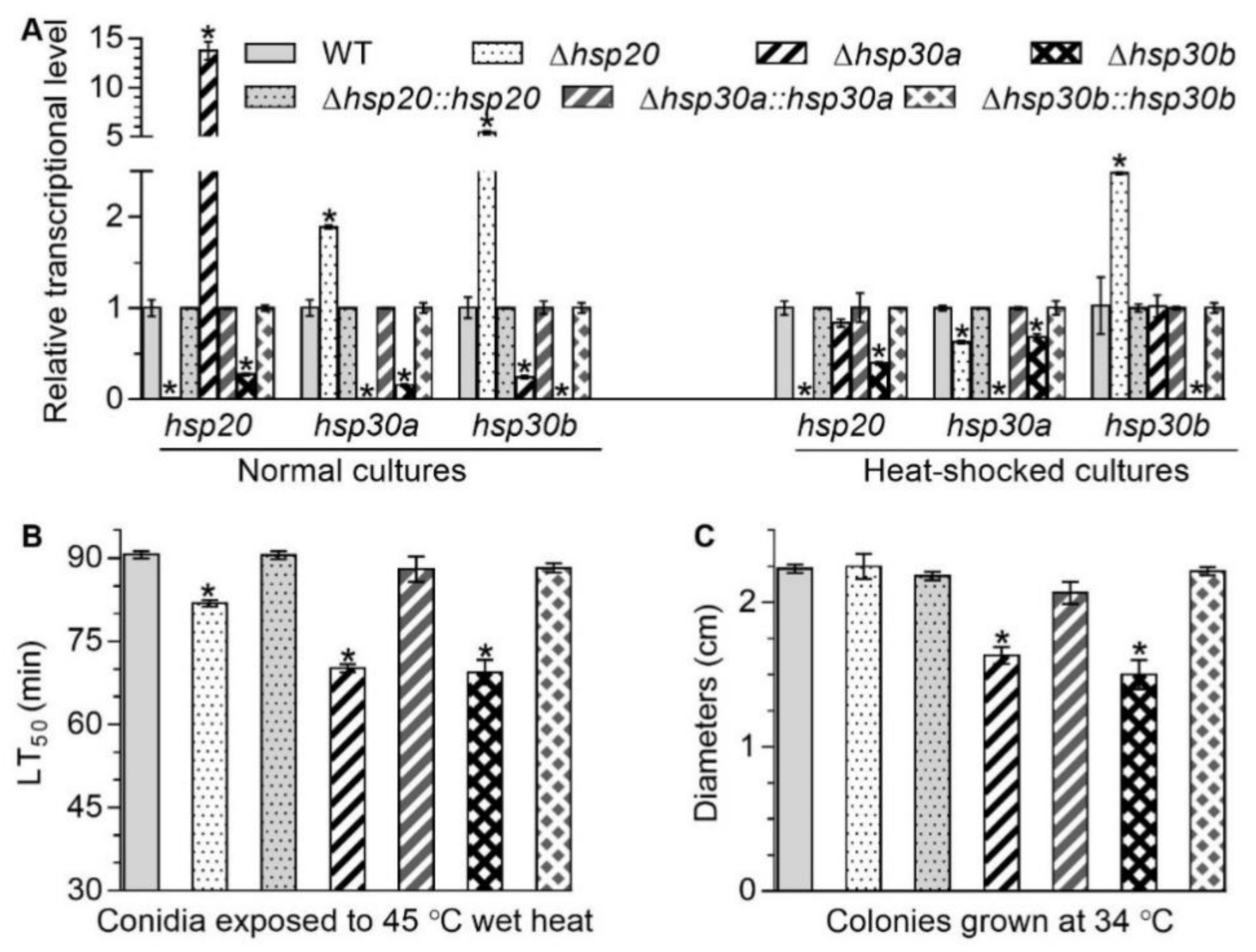
2.2. Interrelationships of hsp20, hsp30a, and hsp30b and Their Roles to Heat Tolerance
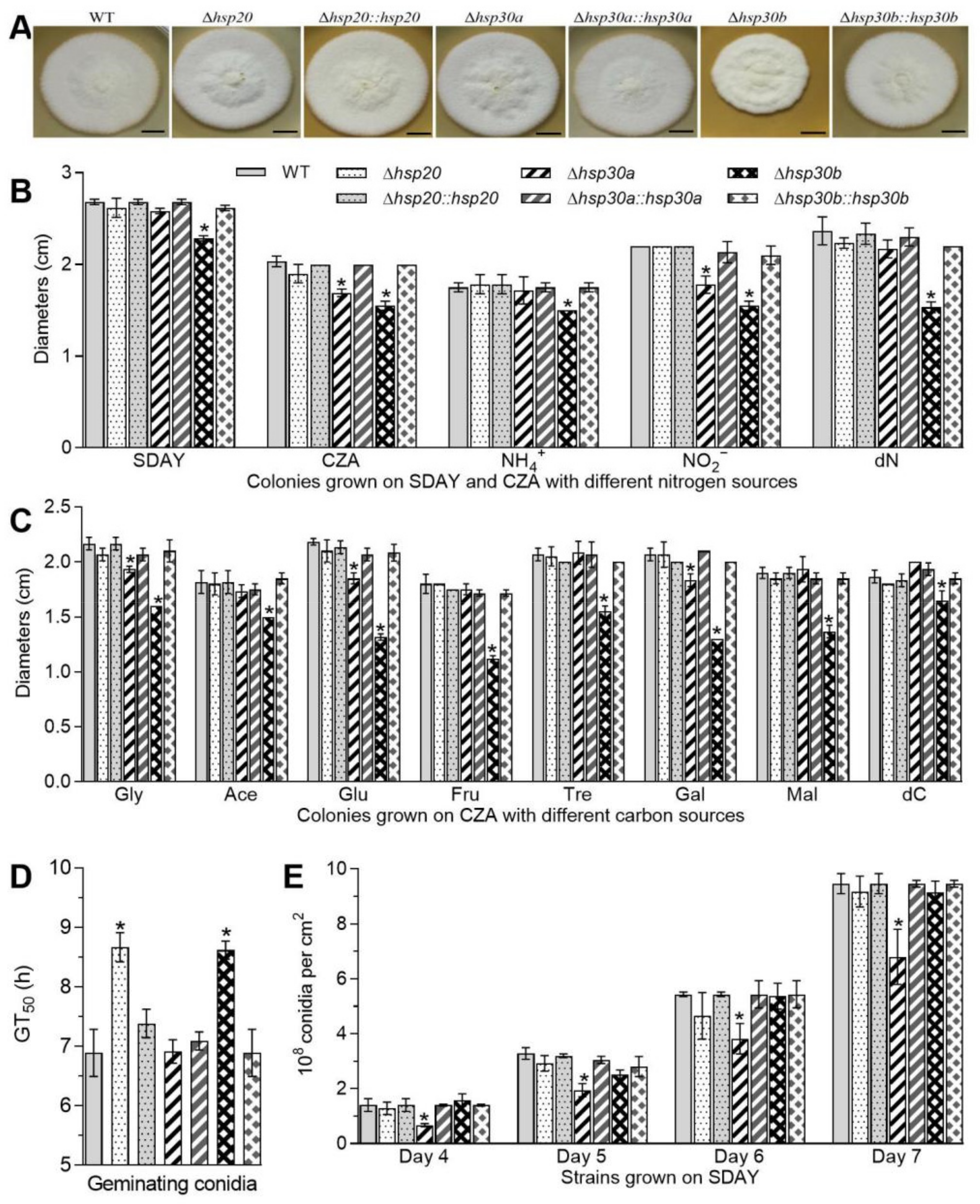
2.3. Differential Roles of hsp20, hsp30a, and hsp30b in Radial Growth and Conidiation
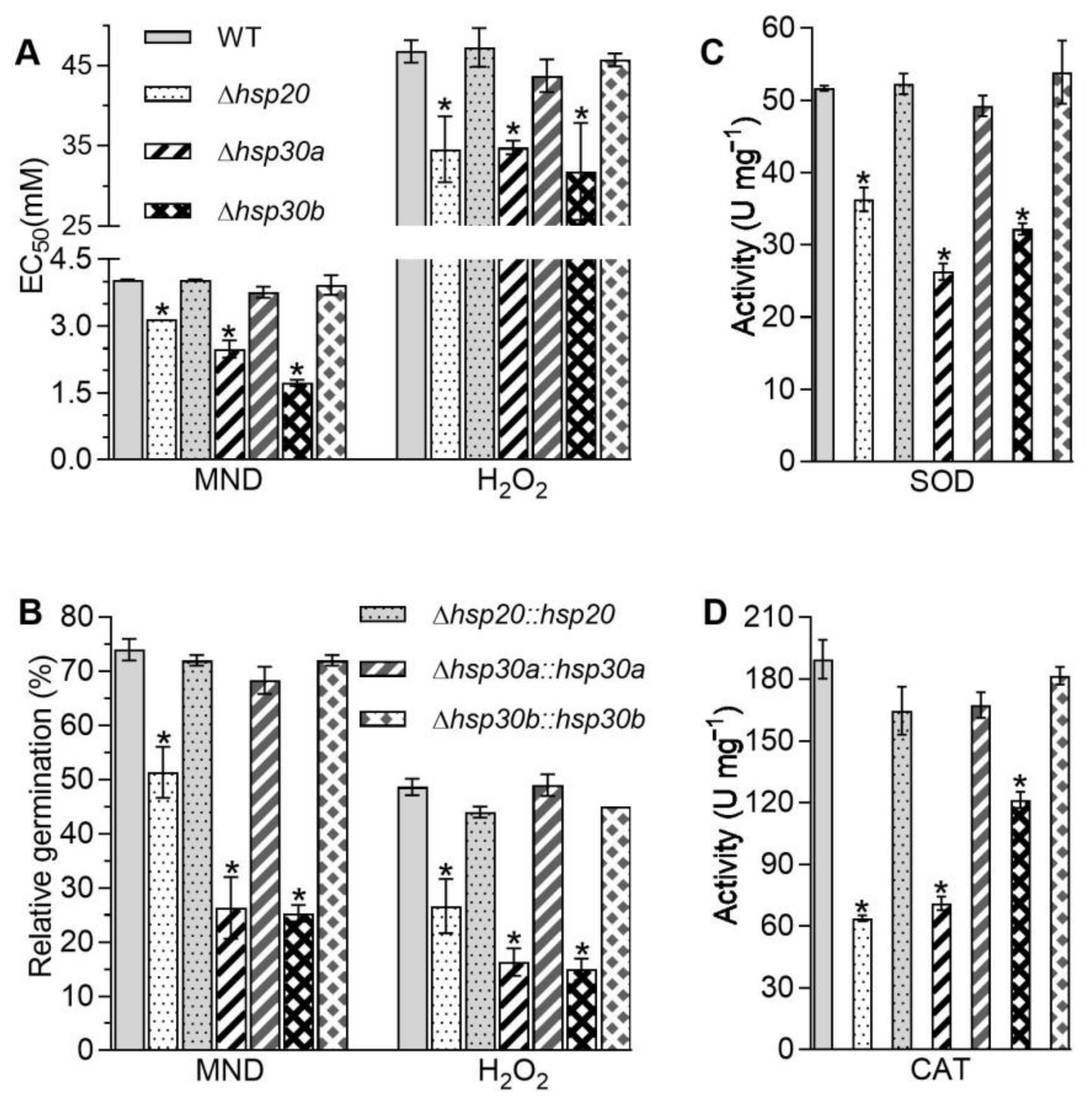
2.4. Essential Roles of hsp20, hsp30a, and hsp30b in Cell Wall Integrity
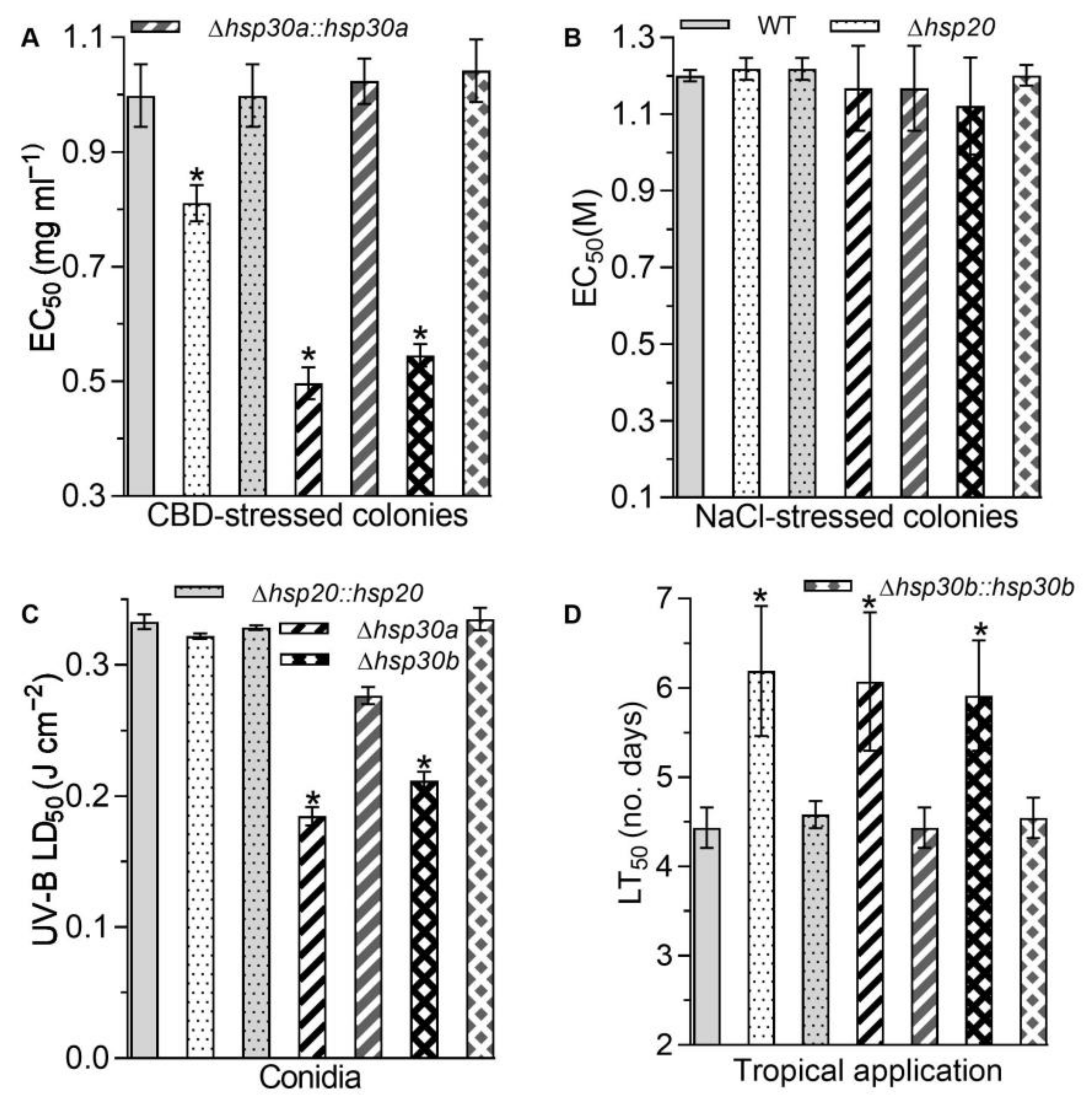
2.5. Contributions of hsp20, hsp30a, and hsp30b to Stress Tolerance and Virulence
3. Discussion
4. Materials and Methods
4.1. Microbial Strains, Culture Conditions, and Chemicals
4.2. Cloning and Analysis of Three sHSPs Containing Conserved α-Crystallin Domain in B. bassiana
4.3. Construction of hsp20, hsp30a, and hsp30b Mutants
4.4. Transcriptional Profiling of hsp20, hsp30a, and hsp30b
4.5. Measurement of Growth and Conidiation
4.6. Assays of Hyphal Responses to Chemical Stresses
4.7. Assays for Conidial Virulence and Tolerance to Various Stresses
4.8. Assessment of Cell Wall Integrity and Antioxidant Enzymes Activities
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| sHsps | Small Heat Shock Proteins |
| WT | wild type |
| CZA | Czapek-Dox agar |
| SDAY | Sabouraud dextrose agar |
| GM | germination medium |
| LB | Luria-Bertani |
| qRT-PCR | quantitative real-time PCR |
| NaAc | acetate |
| LT50 | median lethal time |
| LD50 | median lethal dose |
| GNL | Galanthus nivalis lectin |
| WGA | wheat germ agglutinin |
| SODs | superoxide dismutases |
| CATs | catalases |
| SD | standard deviation |
| ANOVA | analysis of variance |
| HSD | Tukey’s honestly significant difference |
| CBD | carbendazim |
References
- Hideaki, I.; Yohtalou, T. The stress (heat shock) proteins. Int. J. Biochem. 1991, 23, 1185–1191. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.; Ismail, M.; Malek, M.; Latif, M.A.; Oladosu, Y. Heat shock proteins: Functions and response against heat stress in plants. Int. J. Sci. Technol. Res. 2014, 3, 204–218. [Google Scholar]
- Mogk, A.; Ruger-Herreros, C.; Bukau, B. Cellular functions and mechanisms of action of small heat shock proteins. Annu. Rev. Microbiol. 2019, 73, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Franzmann, T.; Weinfurtner, D.; Buchner, J. Some like it hot: The structure and function of small heat-shock proteins. Nat. Struct. Mol. Biol. 2005, 12, 842–846. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell. Mol. Life Sci. 2005, 62, 2460–2476. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Braun, N.; Stromer, T.; Richter, B.; Model, N.; Weinkauf, S.; Buchner, J. Hsp42 is the general small heat shock protein in the cytosol of Saccharomyces cerevisiae. EMBO J. 2004, 23, 638–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petko, L.; Lindquist, S. Hsp26 is not required for growth at high temperatures, nor for thermotolerance, spore development, or germination. Cell 1986, 45, 885–894. [Google Scholar] [CrossRef]
- Specht, S.; Miller, S.B.M.; Mogk, A.; Bukau, B. Hsp42 is required for sequestration of protein aggregates into deposition sites in Saccharomyces cerevisiae. J. Cell Biol. 2011, 195, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Jacobsen, I.D.; Miramón, P.; Slesiona, S.; Bohovych, I.M.; Brown, A.J.; Hube, B. Small but crucial: The novel small heat shock protein Hsp21 mediates stress adaptation and virulence in Candida albicans. PLoS ONE 2012, 7, e38584. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A. Small heat shock proteins (HSP12, HSP20 and HSP30) play a role in Ustilago maydis pathogenesis. FEMS Microbiol. Lett. 2014, 361, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Plesofsky, N.; Brambl, R. Glucose metabolism in Neurospora is altered by heat shock and by disruption of HSP30. Biochim. Biophys. Acta 1999, 1449, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Plesofsky-Vig, N.; Brambl, R. Disruption of the gene for hsp30, an alpha-crystallin-related heat shock protein of Neurospora crassa, causes defects in thermotolerance. Proc. Natl. Acad. Sci. USA 1995, 92, 5032–5036. [Google Scholar] [CrossRef] [Green Version]
- Xiao, G.; Ying, S.-H.; Zheng, P.; Wang, Z.-L.; Zhang, S.; Xie, X.-Q.; Shang, Y.; St Leger, R.J.; Zhao, G.-P.; Wang, C. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Poprawski, T.; Khachatourians, G.G. Production, formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control: Current status. Biocontrol Sci. Technol. 1994, 4, 3–34. [Google Scholar] [CrossRef]
- Wang, J.; Ying, S.H.; Hu, Y.; Feng, M.G. Mas5, a homologue of bacterial DnaJ, is indispensable for the host infection and environmental adaptation of a filamentous fungal insect pathogen. Environ. Microbiol. 2016, 18, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ying, S.-H.; Hu, Y.; Feng, M.-G. Vital role for the J-domain protein Mdj1 in asexual development, multiple stress tolerance, and virulence of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2017, 101, 185–195. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Hu, Y.; Ying, S.-H.; Feng, M.-G. Roles of six Hsp70 genes in virulence, cell wall integrity, antioxidant activity and multiple stress tolerance of Beauveria bassiana. Fungal Genet. Biol. 2020, 144, 103437. [Google Scholar] [CrossRef]
- Wu, J.; Wang, M.; Zhou, L.; Yu, D. Small heat shock proteins, phylogeny in filamentous fungi and expression analyses in Aspergillus nidulans. Gene 2016, 575, 675–679. [Google Scholar] [CrossRef]
- Baek, J.-H.; Park, J.-A.; Kim, J.-M.; Oh, J.-M.; Park, S.-M.; Kim, D.-H. Functional analysis of a tannic-acid-inducible and hypoviral-regulated small heat-shock protein Hsp24 from the chestnut blight fungus Cryphonectria parasitica. Mol. Plant-Microbe Interact. 2014, 27, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.-Q.; Li, F.; Ying, S.-H.; Feng, M.-G. Additive contributions of two manganese-cored superoxide dismutases (MnSODs) to antioxidation, UV tolerance and virulence of Beauveria bassiana. PLoS ONE 2012, 7, e30298. [Google Scholar] [CrossRef]
- Wang, Z.L.; Zhang, L.B.; Ying, S.H.; Feng, M.G. Catalases play differentiated roles in the adaptation of a fungal entomopathogen to environmental stresses. Environ. Microbiol. 2013, 15, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Shi, H.-Q.; Ying, S.-H.; Feng, M.-G. Distinct contributions of one Fe-and two Cu/Zn-cofactored superoxide dismutases to antioxidation, UV tolerance and virulence of Beauveria bassiana. Fungal Genet. Biol. 2015, 81, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Lu, H.-L.; Fang, W.; St Leger, R.J. Overexpression of a Metarhizium robertsii HSP25 gene increases thermotolerance and survival in soil. Appl. Microbiol. Biotechnol. 2014, 98, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Montero-Barrientos, M.; Cardoza, R.E.; Gutiérrez, S.; Monte, E.; Hermosa, R. The heterologous overexpression of hsp23, a small heat-shock protein gene from Trichoderma virens, confers thermotolerance to T. harzianum. Curr. Genet. 2007, 52, 45–53. [Google Scholar] [CrossRef]
- Holder, D.J.; Keyhani, N.O. Adhesion of the entomopathogenic fungus Beauveria (Cordyceps) bassiana to substrata. Appl. Environ. Microbiol. 2005, 71, 5260–5266. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Zhang, Y.; Yang, X.; Zheng, X.; Duan, H.; Li, Y.; Pei, Y. Agrobacterium tumefaciens-mediated transformation of Beauveria bassiana using an herbicide resistance gene as a selection marker. J. Invertebr. Pathol. 2004, 85, 18–24. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zeng, F.; Gong, X.; Hamid, M.I.; Fu, Y.; Jiatao, X.; Cheng, J.; Li, G.; Jiang, D. A fungal cell wall integrity-associated MAP kinase cascade in Coniothyrium minitans is required for conidiation and mycoparasitism. Fungal Genet. Biol. 2012, 49, 347–357. [Google Scholar] [CrossRef]
- Wanchoo, A.; Lewis, M.W.; Keyhani, N.O. Lectin mapping reveals stage-specific display of surface carbohydrates in in vitro and haemolymph-derived cells of the entomopathogenic fungus Beauveria bassiana. Microbiology 2009, 155, 3121–3133. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, J.; Xie, X.-Q.; Keyhani, N.O.; Feng, M.-G.; Ying, S.-H. The autophagy gene BbATG5, involved in the formation of the autophagosome, contributes to cell differentiation and growth but is dispensable for pathogenesis in the entomopathogenic fungus Beauveria bassiana. Microbiology 2013, 159, 243–252. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, G.; Ying, S.-H.; Feng, M.-G.; Wang, J. Differential Roles of Three α-Crystallin Domain-Containing sHsps of Beauveria bassiana in Asexual Development, Multiple Stress Tolerance and Virulence. Int. J. Mol. Sci. 2022, 23, 6717. https://doi.org/10.3390/ijms23126717
Zhou G, Ying S-H, Feng M-G, Wang J. Differential Roles of Three α-Crystallin Domain-Containing sHsps of Beauveria bassiana in Asexual Development, Multiple Stress Tolerance and Virulence. International Journal of Molecular Sciences. 2022; 23(12):6717. https://doi.org/10.3390/ijms23126717
Chicago/Turabian StyleZhou, Gang, Sheng-Hua Ying, Ming-Guang Feng, and Jie Wang. 2022. "Differential Roles of Three α-Crystallin Domain-Containing sHsps of Beauveria bassiana in Asexual Development, Multiple Stress Tolerance and Virulence" International Journal of Molecular Sciences 23, no. 12: 6717. https://doi.org/10.3390/ijms23126717
APA StyleZhou, G., Ying, S. -H., Feng, M. -G., & Wang, J. (2022). Differential Roles of Three α-Crystallin Domain-Containing sHsps of Beauveria bassiana in Asexual Development, Multiple Stress Tolerance and Virulence. International Journal of Molecular Sciences, 23(12), 6717. https://doi.org/10.3390/ijms23126717