
Lysine Deprivation Regulates Npy Expression via GCN2 Signaling Pathway in Mandarin Fish (Siniperca chuatsi)


Abstract
:1. Introduction
2. Results
2.1. Specific Appetite Genes Mainly Contribute to Food Intake Affected by Five Essential Amino Acids
2.2. Lysine Deprivation Might Be Coupled to the GCN2 Pathway to Induce an Increase in Npy Transcription
2.3. Lysine Deprivation Can Activate the GCN2/eIF2α/ATF4 Pathway
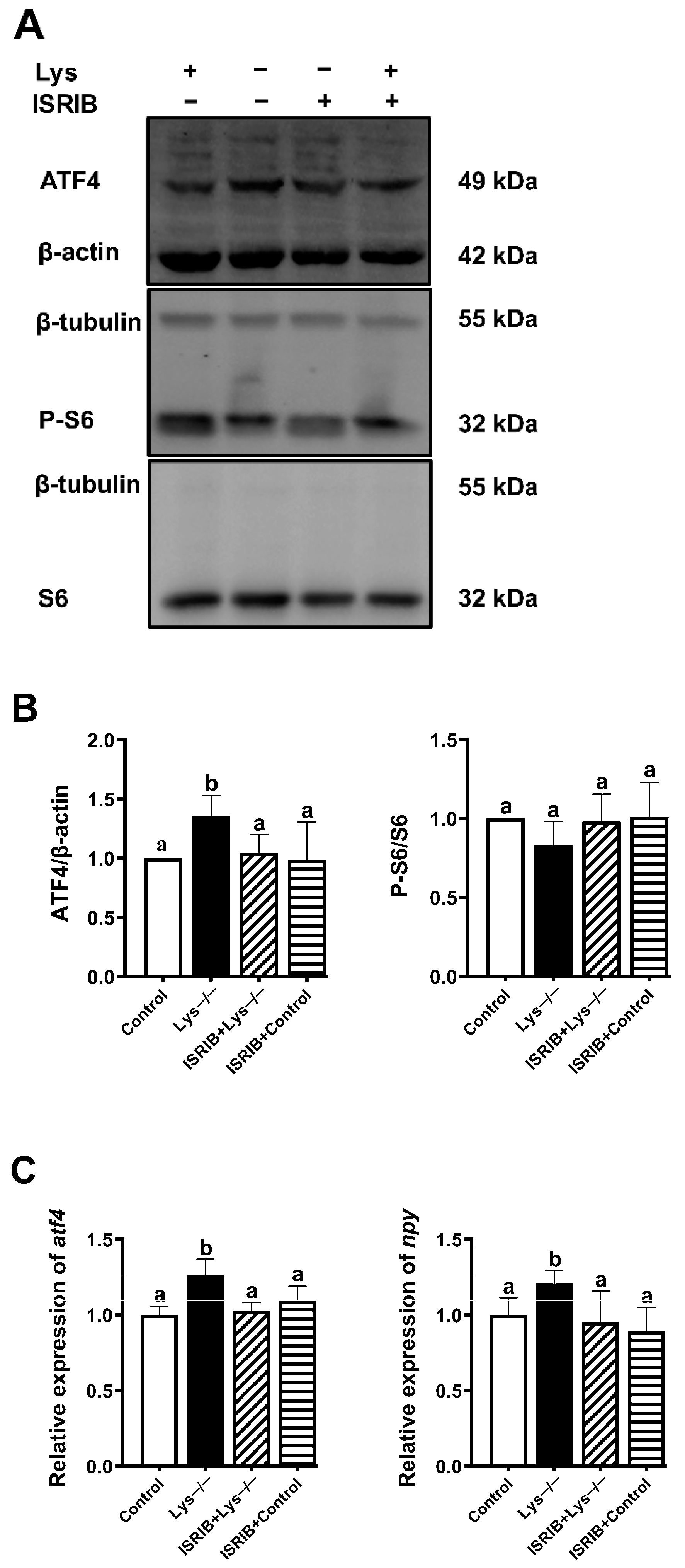
2.4. eIF2α Phosphorylated by GCN2 Kinase Is Essential for the Transcriptional Induction of Npy in Lysine Deprivation
2.5. ATF4 Is Critical for Npy Induction upon Lysine Deprivation
2.6. Npy Immunofluorescence Assay to Validate the Regulatory Function of the GCN2 Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Fish and Reagents
4.2. ICV Administration
4.3. Cell Culture and Treatment
4.4. RNA Extraction and Quantitative PCR
4.5. Western Blotting
4.6. Immunofluorescence Assay
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morton, G.J.; Meek, T.H.; Schwartz, M.W. Neurobiology of food intake in health and disease. Nat. Rev. Neurosci. 2014, 15, 367–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soengas, J.L.; Cerdá-Reverter, J.M.; Delgado, M.J. Central regulation of food intake in fish: An evolutionary perspective. J. Mol. Endocrinol. 2018, 60, R171–R199. [Google Scholar] [CrossRef] [PubMed]
- Blouet, C.; Schwartz, G.J. Hypothalamic nutrient sensing in the control of energy homeostasis. Behav. Brain. Res. 2010, 209, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Conde-Sieira, M.; Soengas, J.L. Nutrient sensing systems in fish: Impact on food intake regulation and energy homeostasis. Front. Neurosci. 2017, 10, 603. [Google Scholar] [CrossRef] [Green Version]
- Schmelzle, T.; Hall, M.N. TOR, a central controller of cell growth. Cell 2000, 103, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Cota, D.; Proulx, K.; Smith, K.A.; Kozma, S.C.; Thomas, G.; Woods, S.C.; Seeley, R.J. Hypothalamic mTOR signaling regulates food intake. Science 2006, 312, 927–930. [Google Scholar] [CrossRef] [Green Version]
- André, C.; Cota, D. Coupling nutrient sensing to metabolic homoeostasis: The role of the mammalian target of rapamycin complex 1 pathway. Proc. Nutr. Soc. 2012, 71, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Pena-Leon, V.; Perez-Lois, R.; Seoane, L.M. mTOR pathway is involved in energy homeostasis regulation as a part of the gut-brain axis. Int. J. Mol. Sci. 2020, 21, 5715. [Google Scholar] [CrossRef]
- Anthony, T.G.; Gietzen, D.W. Detection of amino acid deprivation in the central nervous system. Curr. Opin. Clin. Nutr. Metab. Care. 2013, 16, 96–101. [Google Scholar] [CrossRef]
- Hao, S.; Sharp, J.W.; Ross-Inta, C.M.; McDaniel, B.J.; Anthony, T.G.; Wek, R.C.; Cavener, D.R.; McGrath, B.C.; Rudell, J.B.; Koehnle, T.J.; et al. Uncharged tRNA and sensing of amino acid deficiency in mammalian piriform cortex. Science 2005, 307, 1776–1778. [Google Scholar] [CrossRef]
- Wek, R.C.; Jiang, H.Y.; Anthony, T.G. Coping with stress: eIF2 kinases and translational control. Biochem. Soc. Trans. 2006, 34, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.; et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell. 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Maurin, A.C.; Jousse, C.; Averous, J.; Parry, L.; Bruhat, A.; Cherasse, Y.; Zeng, H.; Zhang, Y.; Harding, H.P.; Ron, D.; et al. The GCN2 kinase biases feeding behavior to maintain amino acid homeostasis in omnivores. Cell. Metab. 2005, 1, 273–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurin, A.C.; Benani, A.; Lorsignol, A.; Brenachot, X.; Parry, L.; Carraro, V.; Guissard, C.; Averous, J.; Jousse, C.; Bruhat, A.; et al. Hypothalamic eIF2α signaling regulates food intake. Cell. Rep. 2014, 6, 438–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Bian, F.; Zhou, H.; Wang, X.; Wang, K.; Mai, K.; He, G. Nutrient sensing and metabolic changes after methionine deprivation in primary muscle cells of turbot (Scophthalmus maximus L.). J. Nutr. Biochem. 2017, 50, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerdá-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the control of food intake in fish. Gen. Comp. Endocrinol. 2005, 142, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K. Recent advances in the regulation of feeding behavior by neuropeptides in fish. Ann. N. Y. Acad. Sci. 2009, 1163, 241–250. [Google Scholar] [CrossRef]
- Otero-Rodiño, C.; Rocha, A.; Sánchez, E.; Álvarez-Otero, R.; Soengas, J.L.; Cerdá-Reverter, J.M. Sensing glucose in the central melanocortin circuits of rainbow trout: A morphological study. Front. Endocrinol. (Lausanne) 2019, 10, 254. [Google Scholar] [CrossRef]
- Comesaña, S.; Velasco, C.; Ceinos, R.M.; López-Patiño, M.A.; Míguez, J.M.; Morais, S.; Soengas, J.L. Evidence for the presence in rainbow trout brain of amino acid-sensing systems involved in the control of food intake. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R201–R215. [Google Scholar] [CrossRef]
- Comesaña, S.; Velasco, C.; Conde-Sieira, M.; Míguez, J.M.; Soengas, J.L.; Morais, S. Feeding stimulation ability and central effects of intraperitoneal treatment of L-leucine, L-valine, and L-proline on amino acid sensing systems in rainbow trout: Implication in food intake control. Front. Physiol. 2018, 9, 1209. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, Z.; Li, J.; Xie, S.; Shi, L.J.; He, Y.H.; Liang, X.F.; Zhu, Q.S.; He, S. Different regulation of branched-chain amino acid on food intake by TOR signaling in Chinese perch (Siniperca chuatsi). Aquaculture 2021, 530, 735792. [Google Scholar] [CrossRef]
- Zhu, Q.S.; He, S.; Liang, X.F.; Chen, K.; Zou, J.M. Effect of histidine and histamine on feeding regulation of Chinese perch (Siniperca chuatsi). J. Huazhong. Agric. Univ. 2020, 39, 180–186. [Google Scholar] [CrossRef]
- Misra, J.; Holmes, M.J.; Mirek, E.T.; Langevin, M.; Kim, H.G.; Carlson, K.R.; Watford, M.; Dong, X.C.; Anthony, T.G.; Wek, R.C. Discordant regulation of eIF2 kinase GCN2 and mTORC1 during nutrient stress. Nucleic. Acids. Res. 2021, 49, 5726–5742. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Nambu, T.; Ebara, S.; Hasegawa, Y.; Toyoshima, K.; Tsuchiya, Y.; Tomita, D.; Fujimoto, J.; Kurasawa, O.; Takahara, C.; et al. Inhibition of GCN2 sensitizes ASNS-low cancer cells to asparaginase by disrupting the amino acid response. Proc. Natl. Acad. Sci. USA 2018, 115, E7776–E7785. [Google Scholar] [CrossRef] [Green Version]
- Sidrauski, C.; Acosta-Alvear, D.; Khoutorsky, A.; Vedantham, P.; Hearn, B.R.; Li, H.; Gamache, K.; Gallagher, C.M.; Ang, K.K.; Wilson, C.; et al. Pharmacological brake-release of mRNA translation enhances cognitive memory. Elife 2013, 2, e00498. [Google Scholar] [CrossRef]
- Heeley, N.; Blouet, C. Central amino acid sensing in the control of feeding behavior. Front. Endocrinol. 2016, 7, 148. [Google Scholar] [CrossRef] [Green Version]
- Laeger, T.; Reed, S.D.; Henagan, T.M.; Fernandez, D.H.; Taghavi, M.; Addington, A.; Münzberg, H.; Martin, R.J.; Hutson, S.M.; Morrison, C.D. Leucine acts in the brain to suppress food intake but does not function as a physiological signal of low dietary protein. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R310–R320. [Google Scholar] [CrossRef] [Green Version]
- Norton, M.; Cheong, W.; Lehmann, A.; Bloom, S.R.; Cox, H.M.; Murphy, K.G. L-arginine promotes gut hormone release and reduces food intake in rodents. Diabetes. Obes. Metab. 2016, 18, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Comesaña, S.; Velasco, C.; Soengas, J.L. Leucine sensing in rainbow trout hypothalamus is direct but separate from mTOR signaling in the regulation of food intake. Aquaculture 2021, 543, 737009. [Google Scholar] [CrossRef]
- Tang, L.; Feng, L.; Sun, C.Y.; Chen, G.F.; Jiang, W.D.; Hu, K.; Liu, Y.; Jiang, J.; Li, S.H.; Kuang, S.Y.; et al. Effect of tryptophan on growth, intestinal enzyme activities and TOR gene expression in juvenile Jian carp (Cyprinus carpio var. Jian): Studies in vivo and in vitro. Aquaculture 2013, 412, 23–33. [Google Scholar] [CrossRef]
- Tan, X.; Lin, H.; Huang, Z.; Zhou, C.; Wang, A.; Qi, C.; Zhao, S. Effects of dietary leucine on growth performance, feed utilization, non-specific immune responses and gut morphology of juvenile golden pompano Trachinotus ovatus. Aquaculture 2016, 465, 100–107. [Google Scholar] [CrossRef]
- Hu, K.; Zhang, J.X.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Jiang, J.; Zhou, X.Q. Effect of dietary glutamine on growth performance, non-specific immunity, expression of cytokine genes, phosphorylation of target of rapamycin (TOR), and anti-oxidative system in spleen and head kidney of Jian carp (Cyprinus carpio var. Jian). Fish. Physiol. Biochem. 2015, 41, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Egerton, S.; Wan, A.; Murphy, K.; Collins, F.; Ahern, G.; Sugrue, I.; Busca, K.; Egan, F.; Muller, N.; Whooley, J.; et al. Replacing fishmeal with plant protein in Atlantic salmon (Salmo salar) diets by supplementation with fish protein hydrolysate. Sci. Rep. 2020, 10, 4194. [Google Scholar] [CrossRef] [PubMed]
- Burr, G.S.; Wolters, W.R.; Barrows, F.T.; Hardy, R.W. Replacing fishmeal with blends of alternative proteins on growth performance of rainbow trout (Oncorhynchus mykiss), and early or late stage juvenile Atlantic salmon (Salmo salar). Aquaculture 2012, 334–337, 110–116. [Google Scholar] [CrossRef]
- Gaylord, T.G.; Barrows, F.T. Multiple amino acid supplementations to reduce dietary protein in plant-based rainbow trout, Oncorhynchus mykiss, feeds. Aquaculture 2009, 287, 180–184. [Google Scholar] [CrossRef]
- Efeyan, A.; Comb, W.C.; Sabatini, D.M. Nutrient-sensing mechanisms and pathways. Nature 2015, 517, 302–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, K.; Sakashita, A.; Yokobori, E.; Azuma, M. Neuroendocrine control of feeding behavior and psychomotor activity by neuropeptide Y in fish. Neuropeptides 2012, 46, 275–283. [Google Scholar] [CrossRef]
- Narnaware, Y.K.; Peyon, P.P.; Lin, X.; Peter, R.E. Regulation of food intake by neuropeptide Y in goldfish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1025–R1034. [Google Scholar] [CrossRef] [Green Version]
- Aldegunde, M.; Mancebo, M. Effects of neuropeptide Y on food intake and brain biogenic amines in the rainbow trout (Oncorhynchus mykiss). Peptides 2006, 27, 719–727. [Google Scholar] [CrossRef]
- Yokobori, E.; Azuma, M.; Nishiguchi, R.; Kang, K.S.; Kamijo, M.; Uchiyama, M.; Matsuda, K. Neuropeptide Y stimulates food intake in the zebrafish, Danio rerio. J. Neuroendocrinol. 2012, 24, 766–773. [Google Scholar] [CrossRef]
- Ye, J.; Palm, W.; Peng, M.; King, B.; Lindsten, T.; Li, M.O.; Koumenis, C.; Thompson, C.B. GCN2 sustains mTORC1 suppression upon amino acid deprivation by inducing Sestrin2. Genes. Dev. 2015, 29, 2331–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; McGrath, B.C.; Reinert, J.; Olsen, D.S.; Lei, L.; Gill, S.; Wek, S.A.; Vattem, K.M.; Wek, R.C.; Kimball, S.R.; et al. The GCN2 eIF2alpha kinase is required for adaptation to amino acid deprivation in mice. Mol. Cell. Biol. 2002, 22, 6681–6688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gietzen, D.W.; Aja, S.M. The brain’s response to an essential amino acid-deficient diet and the circuitous route to a better meal. Mol. Neurobiol. 2012, 6, 332–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanders, D.; Stone, K.P.; Forney, L.A.; Cortez, C.C.; Dille, K.N.; Simon, J.; Xu, M.; Hotard, E.C.; Nikonorova, I.A.; Pettit, A.P.; et al. Role of GCN2-independent signaling through a noncanonical PERK/NRF2 pathway in the physiological responses to dietary methionine restriction. Diabetes 2016, 65, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Tajan, M.; Hennequart, M.; Cheung, E.C.; Zani, F.; Hock, A.K.; Legrave, N.; Maddocks, O.D.K.; Ridgway, R.A.; Athineos, D.; Suárez-Bonnet, A.; et al. Serine synthesis pathway inhibition cooperates with dietary serine and glycine limitation for cancer therapy. Nat. Commun. 2021, 12, 366. [Google Scholar] [CrossRef]
- Morin, G.; Pinel, K.; Dias, K.; Seiliez, I.; Beaumatin, F. RTH-149 cell line, a useful tool to decipher molecular mechanisms related to fish nutrition. Cells 2020, 9, 1754. [Google Scholar] [CrossRef]
- Kilberg, M.S.; Balasubramanian, M.; Fu, L.; Shan, J. The transcription factor network associated with the amino acid response in mammalian cells. Adv. Nutr. 2012, 3, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Bjordal, M.; Arquier, N.; Kniazeff, J.; Pin, J.P.; Léopold, P. Sensing of amino acids in a dopaminergic circuitry promotes rejection of an incomplete diet in Drosophila. Cell 2014, 156, 510–521. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Fortuno, E.S., 3rd; Suh, J.M.; Stenesen, D.; Tang, W.; Parks, E.J.; Adams, C.M.; Townes, T.; Graff, J.M. Atf4 regulates obesity, glucose homeostasis, and energy expenditure. Diabetes 2009, 58, 2565–2573. [Google Scholar] [CrossRef] [Green Version]
- Ohoka, N.; Yoshii, S.; Hattori, T.; Onozaki, K.; Hayashi, H. TRB3, a novel ER stress-inducible gene, is induced via ATF4-CHOP pathway and is involved in cell death. EMBO J. 2005, 24, 1243–1255. [Google Scholar] [CrossRef]
- Zielke, S.; Kardo, S.; Zein, L.; Mari, M.; Covarrubias-Pinto, A.; Kinzler, M.N.; Meyer, N.; Stolz, A.; Fulda, S.; Reggiori, F.; et al. ATF4 links ER stress with reticulophagy in glioblastoma cells. Autophagy 2021, 17, 2432–2448. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhu, L.; Zhu, Y.; Meng, Y.; Li, J.; Song, P.; Yousafzai, N.A.; Feng, L.; Chen, M.; Wang, Y.; et al. Targeting ATF4-dependent pro-survival autophagy to synergize glutaminolysis inhibition. Theranostics 2021, 11, 8464–8479. [Google Scholar] [CrossRef] [PubMed]
- B’chir, W.; Maurin, A.C.; Carraro, V.; Averous, J.; Jousse, C.; Muranishi, Y.; Parry, L.; Stepien, G.; Fafournoux, P.; Bruhat, A. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic. Acids. Res. 2013, 41, 7683–7699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.J.; Liang, X.F.; He, S.; Peng, J. Primary culture and identification of mandarin fish (Siniperca chuatsi) brain neurons. J. Fish. China 2020, 44, 43–48. [Google Scholar] [CrossRef]
- He, S.; Liang, X.F.; Sun, J.; Li, L.; Yu, Y.; Huang, W.; Qu, C.M.; Cao, L.; Bai, X.L.; Tao, Y.X. Insights into food preference in hybrid F1 of Siniperca chuatsi (♀) × Siniperca scherzeri (♂) mandarin fish through transcriptome analysis. BMC. Genom. 2013, 14, 601. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′-3′) | Accession Number | Amplification Size | Anneal Temperature/°C | E-Values /% | R2 |
|---|---|---|---|---|---|---|
| rpl13a | F: TATCCCCCCACCCTATGACA | SC7-LG11-15004 | 100 bp | 59 | 97.7 | 0.995 |
| R: ACGCCCAAGGAGAGCGAACT | ||||||
| npy | F: GGAAGGATACCCGGTGAAA | SC7-LG08-11074 | 201 bp | 53 | 95.4 | 0.993 |
| R: TCTTGACTGTGGAATCGTG | ||||||
| agrp | F: GAGCCAAGCGAAGACCAGA | SC7-LG16-20934 | 151 bp | 60 | 99.4 | 0.978 |
| R: GCAGCACGGCAAATGAGAG | ||||||
| pomc | F: TGTTAGTGGTGGTGATGGC | SC7-LG12-16228 | 268 bp | 58 | 104 | 0.990 |
| R: CTGTCGCTGTGGGCTTTC | ||||||
| cart | F: CTGCTGTCCGTCATTTGTCAC | SC7-LG16-20788 | 171 bp | 60 | 109.8 | 0.998 |
| R: TGGGATGCTTCCTCTTTTCTC | ||||||
| atf4 | F: GGACCAAGATGAAGAAGAAGC | SC7-LG03-03111 | 169 bp | 58 | 94.1 | 0.975 |
| R: CAGCCAGTGGAGCGAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.-M.; Zhu, Q.-S.; Liang, H.; Lu, H.-L.; Liang, X.-F.; He, S. Lysine Deprivation Regulates Npy Expression via GCN2 Signaling Pathway in Mandarin Fish (Siniperca chuatsi). Int. J. Mol. Sci. 2022, 23, 6727. https://doi.org/10.3390/ijms23126727
Zou J-M, Zhu Q-S, Liang H, Lu H-L, Liang X-F, He S. Lysine Deprivation Regulates Npy Expression via GCN2 Signaling Pathway in Mandarin Fish (Siniperca chuatsi). International Journal of Molecular Sciences. 2022; 23(12):6727. https://doi.org/10.3390/ijms23126727
Chicago/Turabian StyleZou, Jia-Ming, Qiang-Sheng Zhu, Hui Liang, Hai-Lin Lu, Xu-Fang Liang, and Shan He. 2022. "Lysine Deprivation Regulates Npy Expression via GCN2 Signaling Pathway in Mandarin Fish (Siniperca chuatsi)" International Journal of Molecular Sciences 23, no. 12: 6727. https://doi.org/10.3390/ijms23126727
APA StyleZou, J. -M., Zhu, Q. -S., Liang, H., Lu, H. -L., Liang, X. -F., & He, S. (2022). Lysine Deprivation Regulates Npy Expression via GCN2 Signaling Pathway in Mandarin Fish (Siniperca chuatsi). International Journal of Molecular Sciences, 23(12), 6727. https://doi.org/10.3390/ijms23126727