
DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
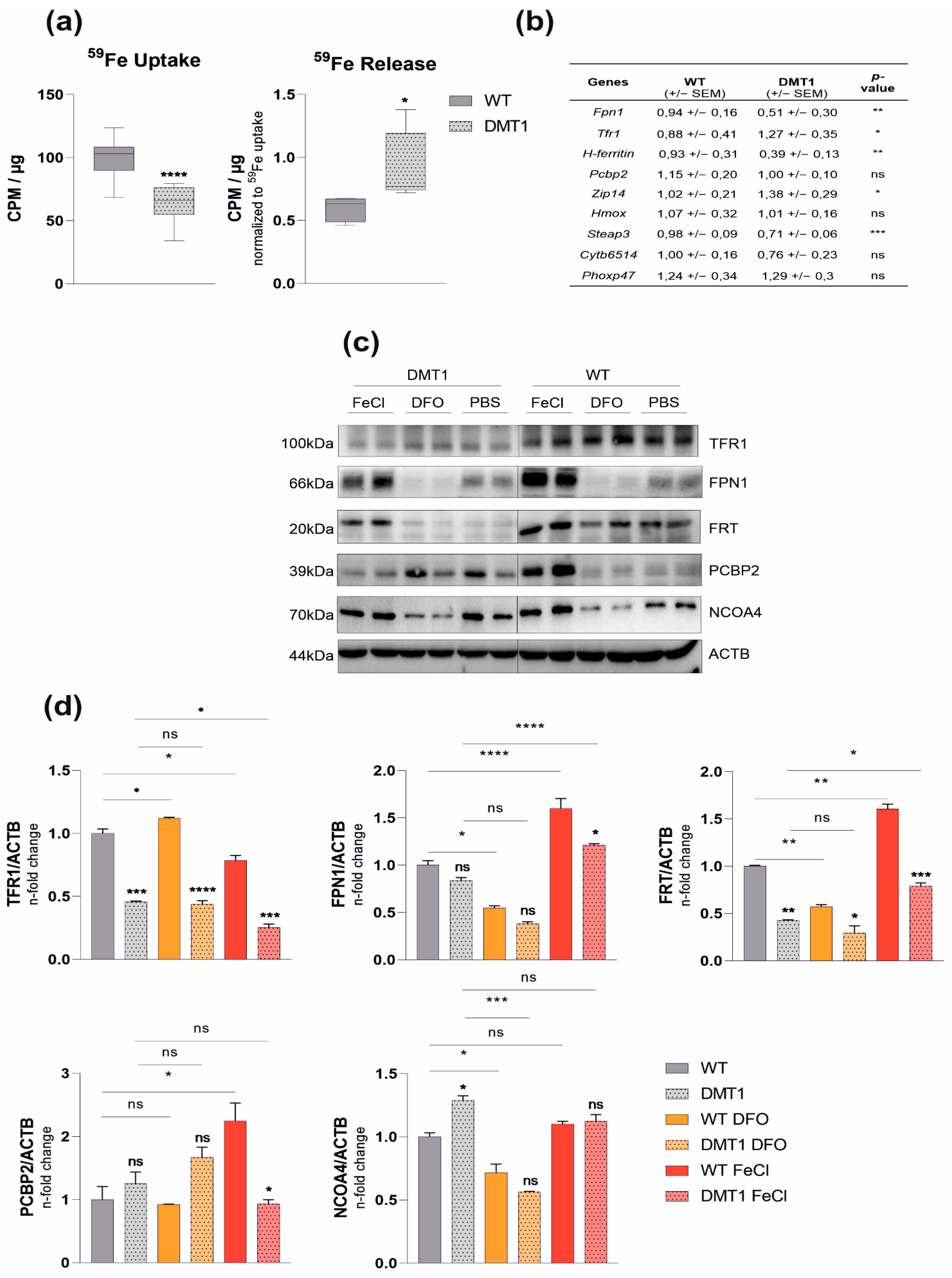
2.1. DMT1 Alters Key Molecules of Iron Homeostasis
2.2. DMT1 Mediates Macrophage Resistance to Salmonella Infection
2.3. DMT1 Modulates Iron Availability in Control and Infected Macrophages
2.4. Macrophage DMT1 Mediates Resistance to Salmonella In Vivo
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Bone Marrow-Derived Macrophages and RAW264.7 Macrophage Culture
4.3. Infection
4.4. CFU
4.5. RNA Analysis
4.6. Protein Analysis
4.7. FACS Analysis
4.8. ELISA
4.9. Determination of Iron Uptake, Release and Content
4.10. Tissue Iron Measurement
4.11. In Vivo Methods
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martins, R.; Carlos, A.R.; Braza, F.; Thompson, J.A.; Bastos-Amador, P.; Ramos, S.; Soares, M.P. Disease Tolerance as an Inherent Component of Immunity. Annu. Rev. Immunol. 2019, 37, 405–437. [Google Scholar] [CrossRef] [Green Version]
- Monteith, A.J.; Skaar, E.P. The Impact of Metal Availability on Immune Function during Infection. Trends Endocrinol. Metab. 2021, 32, 916–928. [Google Scholar] [CrossRef]
- Gerner, R.R.; Nuccio, S.-P.; Raffatellu, M. Iron at the Host-Microbe Interface. Mol. Asp. Med. 2020, 75, 100895. [Google Scholar] [CrossRef]
- Oexle, H.; Gnaiger, E.; Weiss, G. Iron-Dependent Changes in Cellular Energy Metabolism: Influence on Citric Acid Cycle and Oxidative Phosphorylation. Biochim. Biophys. Acta BBA-Bioenerg. 1999, 1413, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Haschka, D.; Hoffmann, A.; Weiss, G. Iron in Immune Cell Function and Host Defense. Semin. Cell Dev. Biol. 2021, 115, 27–36. [Google Scholar] [CrossRef]
- Weiss, G.; Carver, P.L. Role of Divalent Metals in Infectious Disease Susceptibility and Outcome. Clin. Microbiol. Infect. 2018, 24, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Weiss, G.; Goodnough, L.T. Anemia of Chronic Disease. N. Engl. J. Med. 2005, 352, 1011–1023. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T.; Nemeth, E. Iron Homeostasis in Host Defence and Inflammation. Nat. Rev. Immunol. 2015, 15, 500–510. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Theurl, I.; Swirski, F.K.; Weiss, G. “Pumping Iron”—How Macrophages Handle Iron at the Systemic, Microenvironmental, and Cellular Levels. Pflüg. Arch.-Eur. J. Physiol. 2017, 469, 397–418. [Google Scholar] [CrossRef] [Green Version]
- Schaible, U.E.; Kaufmann, S.H.E. Iron and Microbial Infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef]
- Halaas, Ø.; Steigedal, M.; Haug, M.; Awuh, J.A.; Ryan, L.; Brech, A.; Sato, S.; Husebye, H.; Cangelosi, G.A.; Akira, S.; et al. Intracellular Mycobacterium Avium Intersect Transferrin in the Rab11 + Recycling Endocytic Pathway and Avoid Lipocalin 2 Trafficking to the Lysosomal Pathway. J. Infect. Dis. 2010, 201, 783–792. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Metzendorf, C.; Vujic-Spasic, M.; Mitterstiller, A.-M.; Schroll, A.; Haschka, D.; Hoffmann, A.; Von Raffay, L.; Sparla, R.; Huck, C.W.; et al. Cell-Specific Expression of Hfe Determines the Outcome of Salmonella enterica Serovar Typhimurium Infection in Mice. Haematologica 2021, 106, 3149–3161. [Google Scholar] [CrossRef]
- Golonka, R.; Yeoh, B.S.; Vijay-Kumar, M. The Iron Tug-of-War between Bacterial Siderophores and Innate Immunity. J. Innate Immun. 2019, 11, 249–262. [Google Scholar] [CrossRef]
- Dichtl, S.; Haschka, D.; Nairz, M.; Seifert, M.; Volani, C.; Lutz, O.; Weiss, G. Dopamine Promotes Cellular Iron Accumulation and Oxidative Stress Responses in Macrophages. Biochem. Pharmacol. 2018, 148, 193–201. [Google Scholar] [CrossRef]
- Liu, Z.; Reba, S.; Chen, W.-D.; Porwal, S.K.; Boom, W.H.; Petersen, R.B.; Rojas, R.; Viswanathan, R.; Devireddy, L. Regulation of Mammalian Siderophore 2,5-DHBA in the Innate Immune Response to Infection. J. Exp. Med. 2014, 211, 1197–1213. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Ferring-Appel, D.; Casarrubea, D.; Sonnweber, T.; Viatte, L.; Schroll, A.; Haschka, D.; Fang, F.C.; Hentze, M.W.; Weiss, G.; et al. Iron Regulatory Proteins Mediate Host Resistance to Salmonella Infection. Cell Host Microbe 2015, 18, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Siegert, I.; Schödel, J.; Nairz, M.; Schatz, V.; Dettmer, K.; Dick, C.; Kalucka, J.; Franke, K.; Ehrenschwender, M.; Schley, G.; et al. Ferritin-Mediated Iron Sequestration Stabilizes Hypoxia-Inducible Factor-1α upon LPS Activation in the Presence of Ample Oxygen. Cell Rep. 2015, 13, 2048–2055. [Google Scholar] [CrossRef] [Green Version]
- Yanatori, I.; Kishi, F. DMT1 and Iron Transport. Free Radic. Biol. Med. 2019, 133, 55–63. [Google Scholar] [CrossRef]
- Ludwiczek, S.; Theurl, I.; Muckenthaler, M.U.; Jakab, M.; Mair, S.M.; Theurl, M.; Kiss, J.; Paulmichl, M.; Hentze, M.W.; Ritter, M.; et al. Ca2+ Channel Blockers Reverse Iron Overload by a New Mechanism via Divalent Metal Transporter-1. Nat. Med. 2007, 13, 448–454. [Google Scholar] [CrossRef]
- Mair, S.M.; Nairz, M.; Bellmann-Weiler, R.; Muehlbacher, T.; Schroll, A.; Theurl, I.; Moser, P.L.; Talasz, H.; Fang, F.C.; Weiss, G. Nifedipine Affects the Course of Salmonella Enterica Serovar Typhimurium Infection by Modulating Macrophage Iron Homeostasis. J. Infect. Dis. 2011, 204, 685–694. [Google Scholar] [CrossRef]
- Metzger, D.; Chambon, P. Site- and Time-Specific Gene Targeting in the Mouse. Methods 2001, 24, 71–80. [Google Scholar] [CrossRef]
- Fleming, M.D.; Romano, M.A.; Su, M.A.; Garrick, L.M.; Garrick, M.D.; Andrews, N.C. Nramp2 Is Mutated in the Anemic Belgrade (b) Rat: Evidence of a Role for Nramp2 in Endosomal Iron Transport. Proc. Natl. Acad. Sci. USA 1998, 95, 1148–1153. [Google Scholar] [CrossRef] [Green Version]
- Fleming, M.D.; Trenor, C.C.; Su, M.A.; Foernzler, D.; Beier, D.R.; Dietrich, W.F.; Andrews, N.C. Microcytic Anaemia Mice Have a Mutation in Nramp2, a Candidate Iron Transporter Gene. Nat. Genet. 1997, 16, 383–386. [Google Scholar] [CrossRef]
- Ganz, T. Iron in Innate Immunity: Starve the Invaders. Curr. Opin. Immunol. 2009, 21, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Theurl, I.; Ludwiczek, S.; Theurl, M.; Mair, S.M.; Fritsche, G.; Weiss, G. The Co-Ordinated Regulation of Iron Homeostasis in Murine Macrophages Limits the Availability of Iron for Intracellular Salmonella Typhimurium. Cell. Microbiol. 2007, 9, 2126–2140. [Google Scholar] [CrossRef]
- van Santen, S.; de Mast, Q.; Swinkels, D.W.; van der Ven, A.J.A.M. The Iron Link between Malaria and Invasive Non-Typhoid Salmonella Infections. Trends Parasitol. 2013, 29, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Schroll, A.; Haschka, D.; Dichtl, S.; Sonnweber, T.; Theurl, I.; Theurl, M.; Lindner, E.; Demetz, E.; Aßhoff, M.; et al. Lipocalin-2 Ensures Host Defense against Salmonella Typhimurium by Controlling Macrophage Iron Homeostasis and Immune Response. Eur. J. Immunol. 2015, 45, 3073–3086. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Theurl, I.; Schroll, A.; Theurl, M.; Fritsche, G.; Lindner, E.; Seifert, M.; Crouch, M.-L.V.; Hantke, K.; Akira, S.; et al. Absence of Functional Hfe Protects Mice from Invasive Salmonella Enterica Serovar Typhimurium Infection via Induction of Lipocalin-2. Blood 2009, 114, 3642–3651. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.-J.; Feng, D.; Wu, H.; Wang, H.; Chan, Y.; Kolls, J.; Borregaard, N.; Porse, B.; Berger, T.; Mak, T.W.; et al. Liver Is the Major Source of Elevated Serum Lipocalin-2 Levels after Bacterial Infection or Partial Hepatectomy: A Critical Role for IL-6/STAT3. Hepatology 2015, 61, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Soares, M.P.; Weiss, G. The Iron Age of Host–Microbe Interactions. EMBO Rep. 2015, 16, 1482–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drakesmith, H.; Prentice, A.M. Hepcidin and the Iron-Infection Axis. Science 2012, 338, 768–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flo, T.H.; Smith, K.D.; Sato, S.; Rodriguez, D.J.; Holmes, M.A.; Strong, R.K.; Akira, S.; Aderem, A. Lipocalin 2 Mediates an Innate Immune Response to Bacterial Infection by Sequestrating Iron. Nature 2004, 432, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.R.; Gros, P. Divalent-Metal Transport by NRAMP Proteins at the Interface of Host–Pathogen Interactions. Trends Microbiol. 2001, 9, 397–403. [Google Scholar] [CrossRef]
- Jeney, V.; Ramos, S.; Bergman, M.-L.; Bechmann, I.; Tischer, J.; Ferreira, A.; Oliveira-Marques, V.; Janse, C.J.; Rebelo, S.; Cardoso, S.; et al. Control of Disease Tolerance to Malaria by Nitric Oxide and Carbon Monoxide. Cell Rep. 2014, 8, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haschka, D.; Tymoszuk, P.; Petzer, V.; Hilbe, R.; Heeke, S.; Dichtl, S.; Skvortsov, S.; Demetz, E.; Berger, S.; Seifert, M.; et al. Ferritin H Deficiency Deteriorates Cellular Iron Handling and Worsens Salmonella Typhimurium Infection by Triggering Hyperinflammation. JCI Insight 2021, 6, e141760. [Google Scholar] [CrossRef] [PubMed]
- Tuckermann, J.P.; Kleiman, A.; Moriggl, R.; Spanbroek, R.; Neumann, A.; Illing, A.; Clausen, B.E.; Stride, B.; Förster, I.; Habenicht, A.J.R.; et al. Macrophages and Neutrophils Are the Targets for Immune Suppression by Glucocorticoids in Contact Allergy. J. Clin. Investig. 2007, 117, 1381–1390. [Google Scholar] [CrossRef]
- Veuthey, T.; Hoffmann, D.; Vaidya, V.S.; Wessling-Resnick, M. Impaired Renal Function and Development in Belgrade Rats. Am. J. Physiol. Renal Physiol. 2014, 306, F333–F343. [Google Scholar] [CrossRef] [Green Version]
- Haschka, D.; Grander, M.; Eibensteiner, J.; Dichtl, S.; Koppelstätter, S.; Weiss, G. Nifedipine Potentiates Susceptibility of Salmonella Typhimurium to Different Classes of Antibiotics. Antibiotics 2021, 10, 1200. [Google Scholar] [CrossRef]
- Mertens, C.; Mora, J.; Ören, B.; Grein, S.; Winslow, S.; Scholich, K.; Weigert, A.; Malmström, P.; Forsare, C.; Fernö, M.; et al. Macrophage-Derived Lipocalin-2 Transports Iron in the Tumor Microenvironment. OncoImmunology 2018, 7, e1408751. [Google Scholar] [CrossRef] [Green Version]
- Yanatori, I.; Richardson, D.R.; Imada, K.; Kishi, F. Iron Export through the Transporter Ferroportin 1 Is Modulated by the Iron Chaperone PCBP2. J. Biol. Chem. 2016, 291, 17303–17318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.; Kim, K.S.; Jeong, J.-H.; Marques, O.; kim, H.-J.; Song, M.; Lee, T.-H.; Kim, J.I.; Choi, H.-S.; Min, J.-J.; et al. The Hepcidin-Ferroportin Axis Controls the Iron Content of Salmonella-Containing Vacuoles in Macrophages. Nat. Commun. 2018, 9, 2091. [Google Scholar] [CrossRef] [Green Version]
- Chlosta, S.; Fishman, D.S.; Harrington, L.; Johnson, E.E.; Knutson, M.D.; Wessling-Resnick, M.; Cherayil, B.J. The Iron Efflux Protein Ferroportin Regulates the Intracellular Growth of Salmonella enterica. Infect. Immun. 2006, 74, 3065–3067. [Google Scholar] [CrossRef] [Green Version]
- Philpott, C.C.; Ryu, M.-S.; Frey, A.; Patel, S. Cytosolic Iron Chaperones: Proteins Delivering Iron Cofactors in the Cytosol of Mammalian Cells. J. Biol. Chem. 2017, 292, 12764–12771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.J.R.; Richardson, D.R. Chaperone Turns Gatekeeper: PCBP2 and DMT1 Form an Iron-Transport Pipeline. Biochem. J. 2014, 462, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Soe-Lin, S.; Apte, S.S.; Mikhael, M.R.; Kayembe, L.K.; Nie, G.; Ponka, P. Both Nramp1 and DMT1 Are Necessary for Efficient Macrophage Iron Recycling. Exp. Hematol. 2010, 38, 609–617. [Google Scholar] [CrossRef]
- Turcu, A.L.; Versini, A.; Khene, N.; Gaillet, C.; Cañeque, T.; Müller, S.; Rodriguez, R. DMT1 Inhibitors Kill Cancer Stem Cells by Blocking Lysosomal Iron Translocation. Chem.—Eur. J. 2020, 26, 7369–7373. [Google Scholar] [CrossRef]
- Tenopoulou, M.; Kurz, T.; Doulias, P.-T.; Galaris, D.; Brunk, U.T. Does the Calcein-AM Method Assay the Total Cellular ‘Labile Iron Pool’ or Only a Fraction of It? Biochem. J. 2007, 403, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Haschka, D.; Nairz, M.; Demetz, E.; Wienerroither, S.; Decker, T.; Weiss, G. Contrasting Regulation of Macrophage Iron Homeostasis in Response to Infection with Listeria Monocytogenes Depending on Localization of Bacteria. Metallomics 2015, 7, 1036–1045. [Google Scholar] [CrossRef]
- Petzer, V.; Tymoszuk, P.; Asshoff, M.; Carvalho, J.; Papworth, J.; Deantonio, C.; Bayliss, L.; Wake, M.S.; Seifert, M.; Brigo, N.; et al. A Fully Human Anti-BMP6 Antibody Reduces the Need for Erythropoietin in Rodent Models of the Anemia of Chronic Disease. Blood 2020, 136, 1080–1090. [Google Scholar] [CrossRef]
- Hoffmann, A.; Haschka, D.; Valente de Souza, L.; Tymoszuk, P.; Seifert, M.; von Raffay, L.; Hilbe, R.; Petzer, V.; Moser, P.L.; Nairz, M.; et al. Baseline Iron Status and Presence of Anaemia Determine the Course of Systemic Salmonella Infection Following Oral Iron Supplementation in Mice. EBioMedicine 2021, 71, 103568. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grander, M.; Hoffmann, A.; Seifert, M.; Demetz, E.; Grubwieser, P.; Pfeifhofer-Obermair, C.; Haschka, D.; Weiss, G. DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression. Int. J. Mol. Sci. 2022, 23, 6789. https://doi.org/10.3390/ijms23126789
Grander M, Hoffmann A, Seifert M, Demetz E, Grubwieser P, Pfeifhofer-Obermair C, Haschka D, Weiss G. DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression. International Journal of Molecular Sciences. 2022; 23(12):6789. https://doi.org/10.3390/ijms23126789
Chicago/Turabian StyleGrander, Manuel, Alexander Hoffmann, Markus Seifert, Egon Demetz, Philipp Grubwieser, Christa Pfeifhofer-Obermair, David Haschka, and Günter Weiss. 2022. "DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression" International Journal of Molecular Sciences 23, no. 12: 6789. https://doi.org/10.3390/ijms23126789
APA StyleGrander, M., Hoffmann, A., Seifert, M., Demetz, E., Grubwieser, P., Pfeifhofer-Obermair, C., Haschka, D., & Weiss, G. (2022). DMT1 Protects Macrophages from Salmonella Infection by Controlling Cellular Iron Turnover and Lipocalin 2 Expression. International Journal of Molecular Sciences, 23(12), 6789. https://doi.org/10.3390/ijms23126789