
A Small Molecule That Promotes Cellular Senescence Prevents Fibrogenesis and Tumorigenesis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
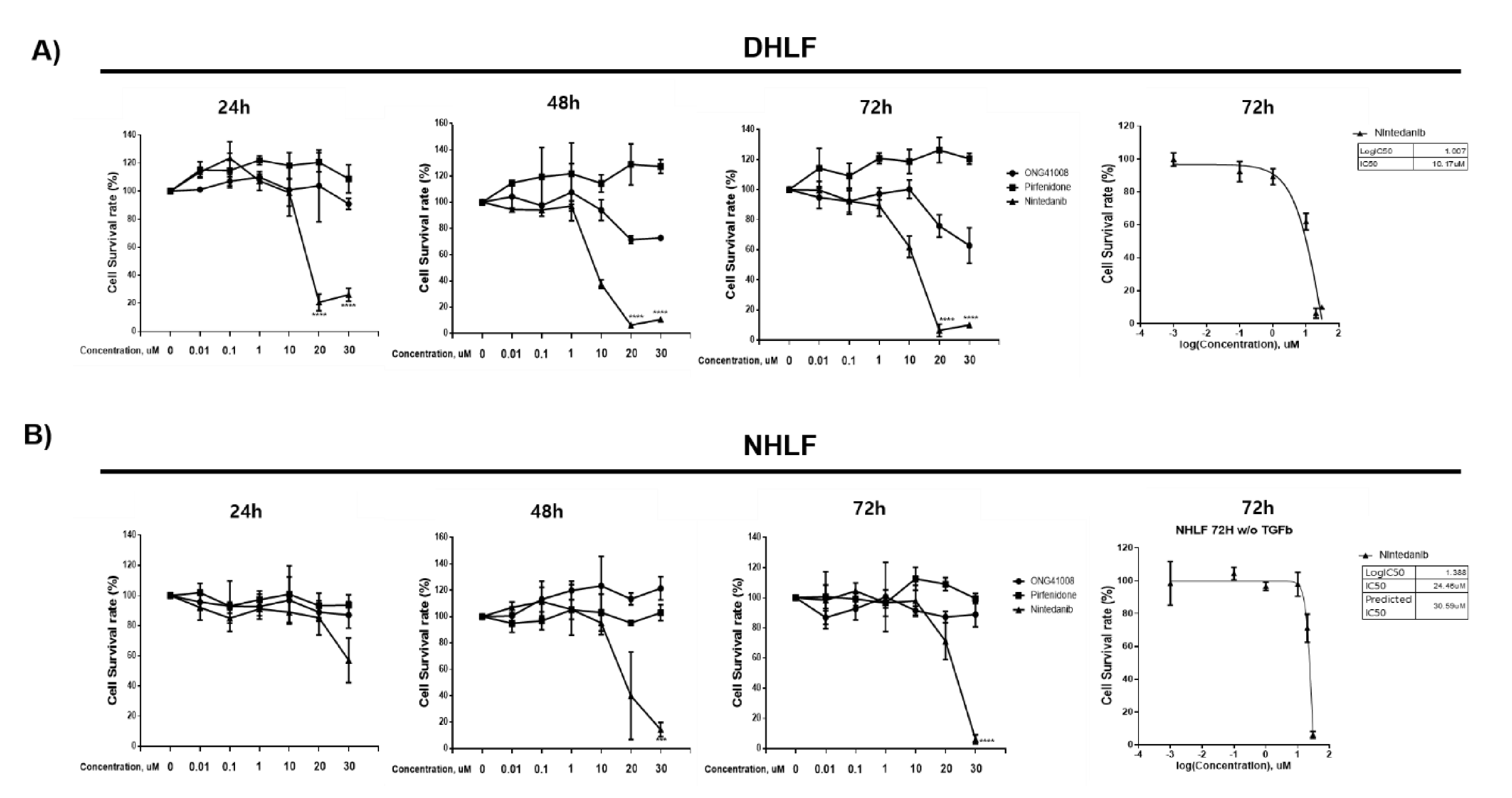
2.1. ONG41008 Induced Cellular Senescence in DHLFs
2.2. ONG41008 Induced Cellular Senescence Similarly to Dasatinib in DHLFs
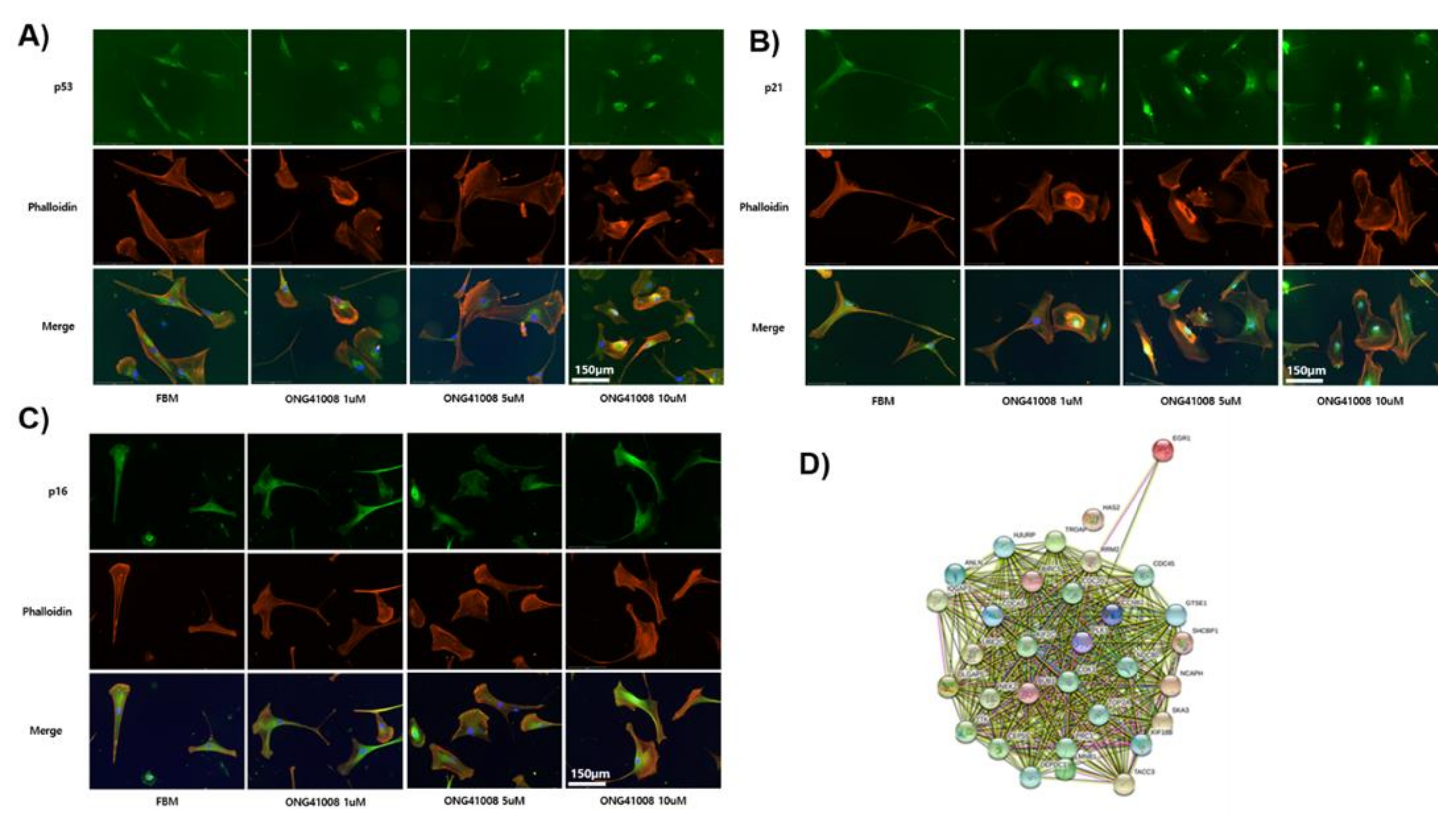
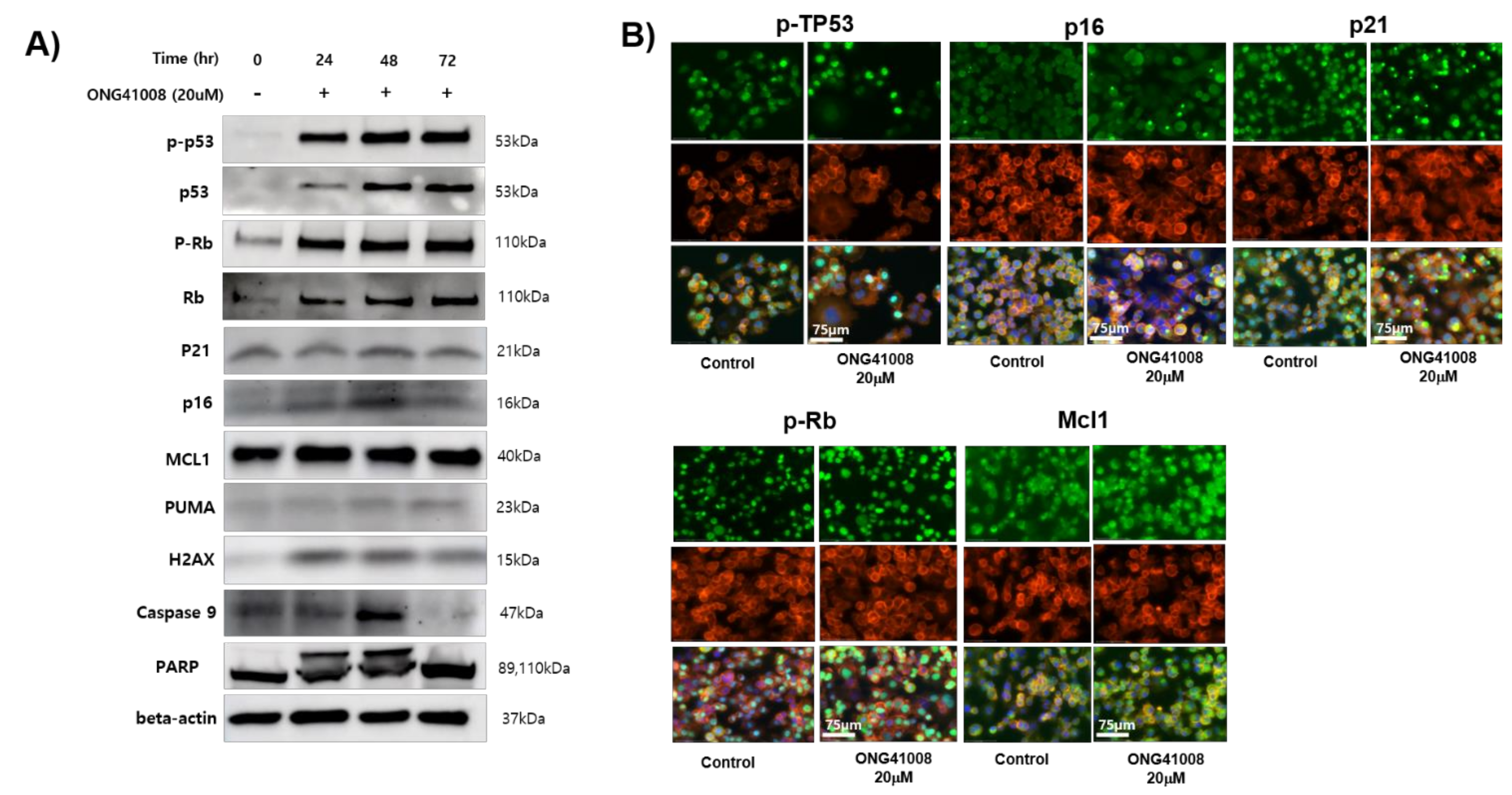
2.3. TP53, p21, and p16 Activities Were Required for ONG41008-Induced Cellular Senescence in DHLFs
2.4. ONG41008 Generated Several Potential Interactomes in DHLFs during Transdifferentiation into RS
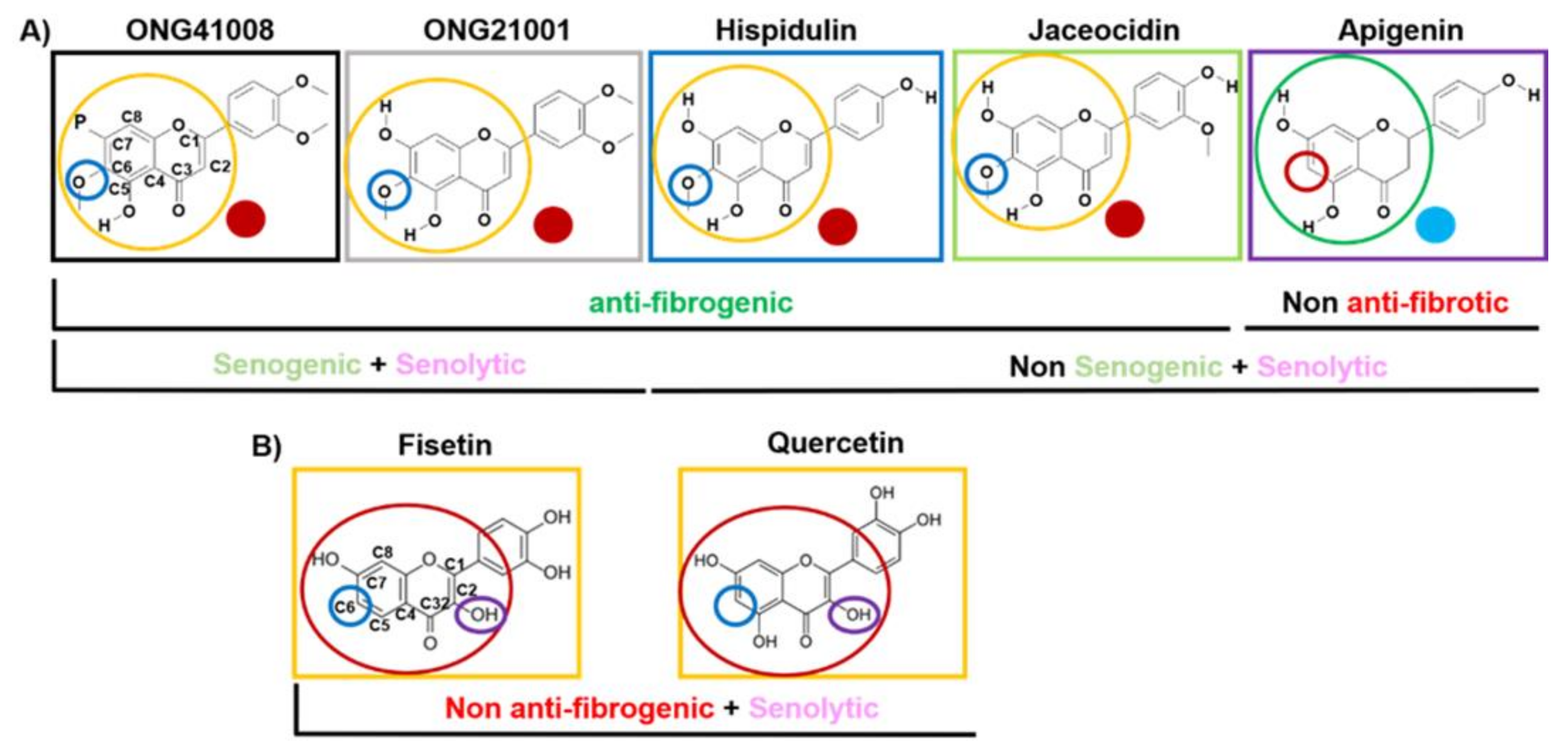
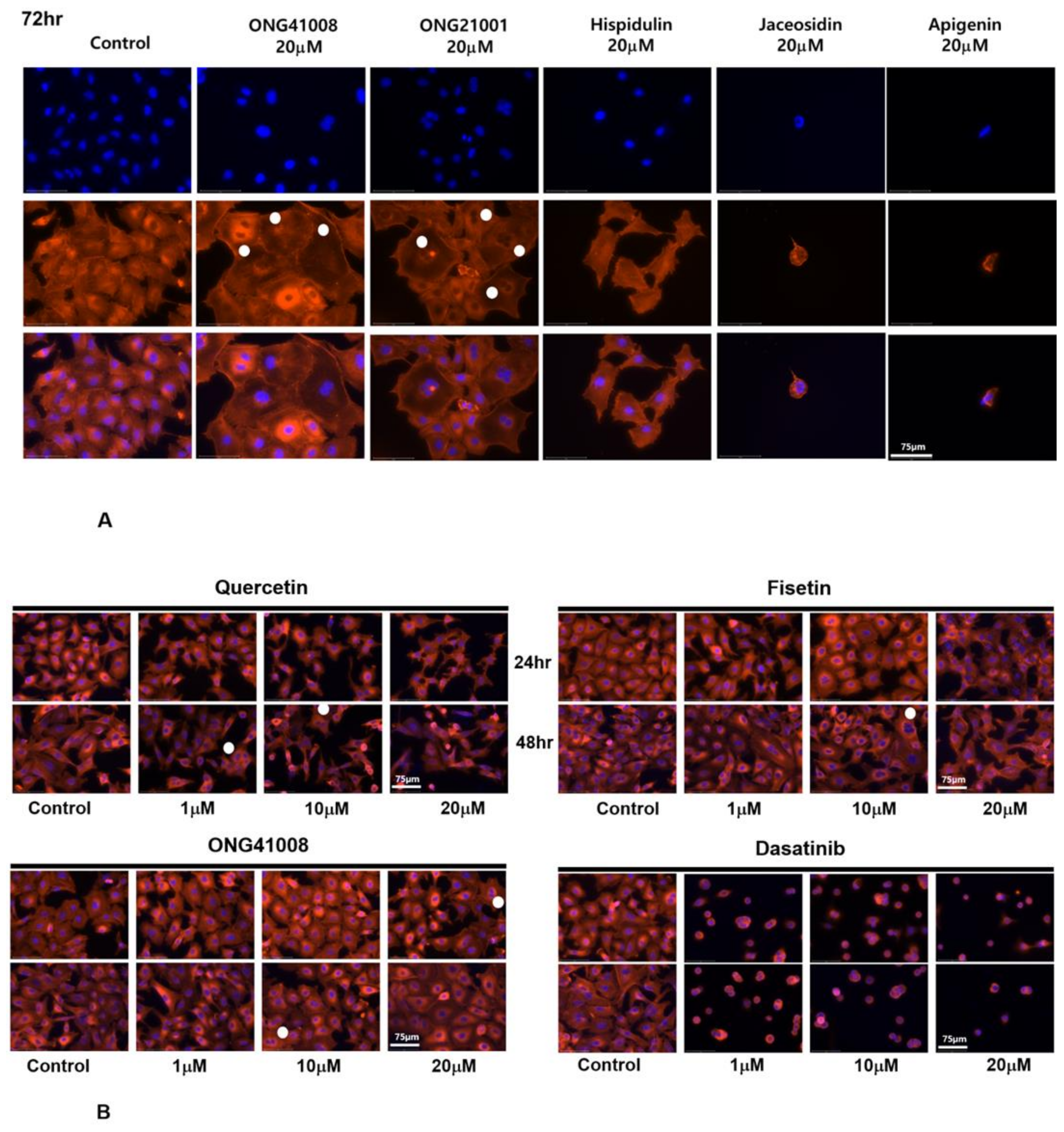
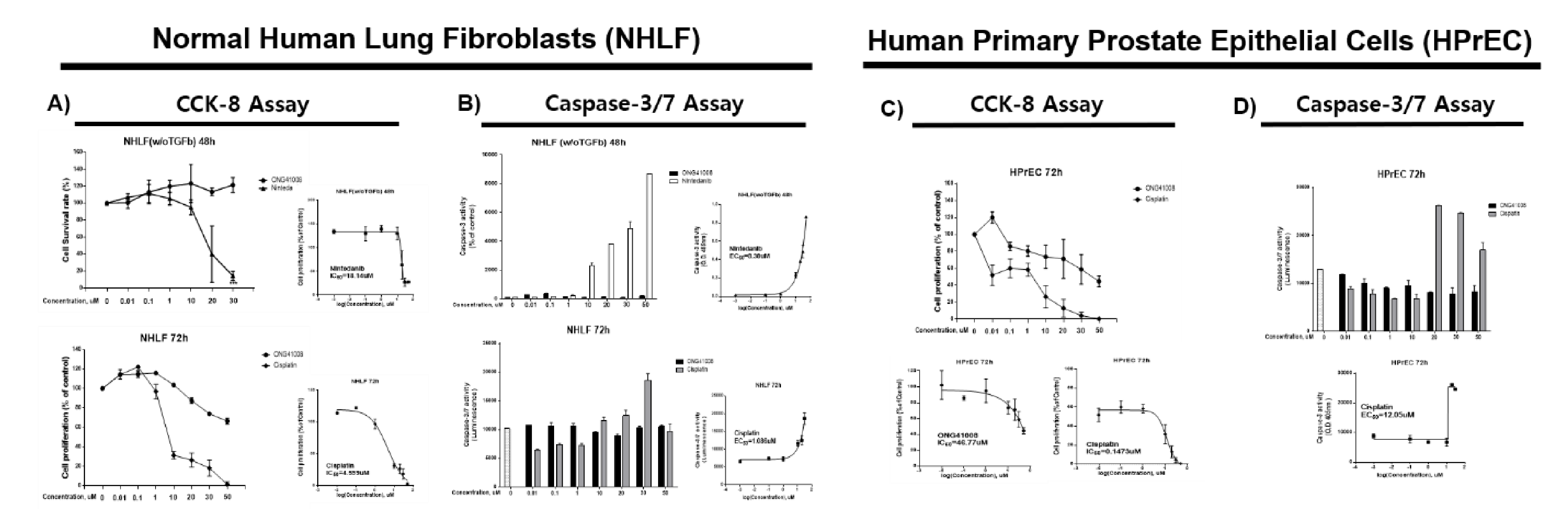
2.5. ONG41008 Harbors Cancer-Cell Senogenic Potential
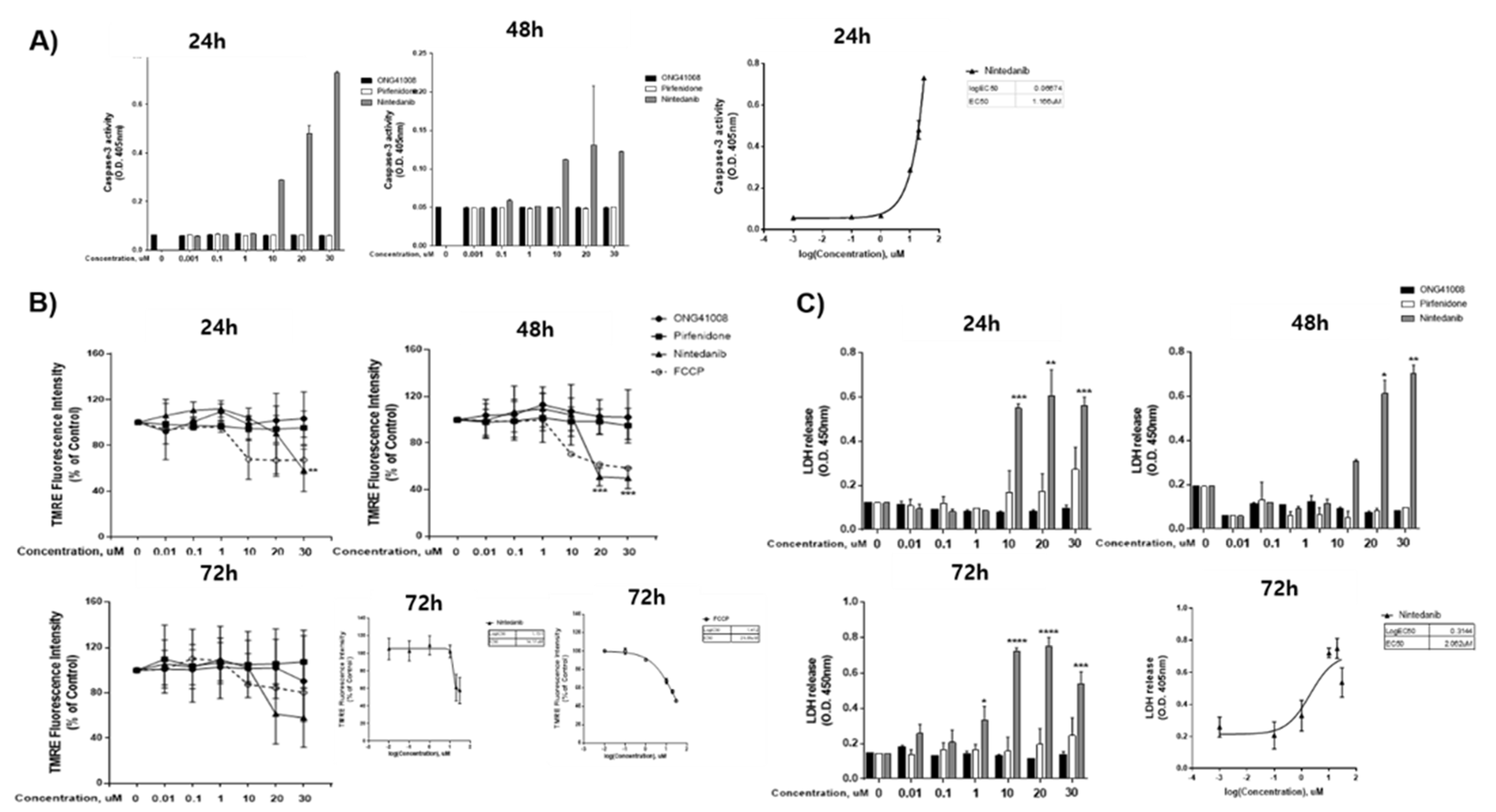
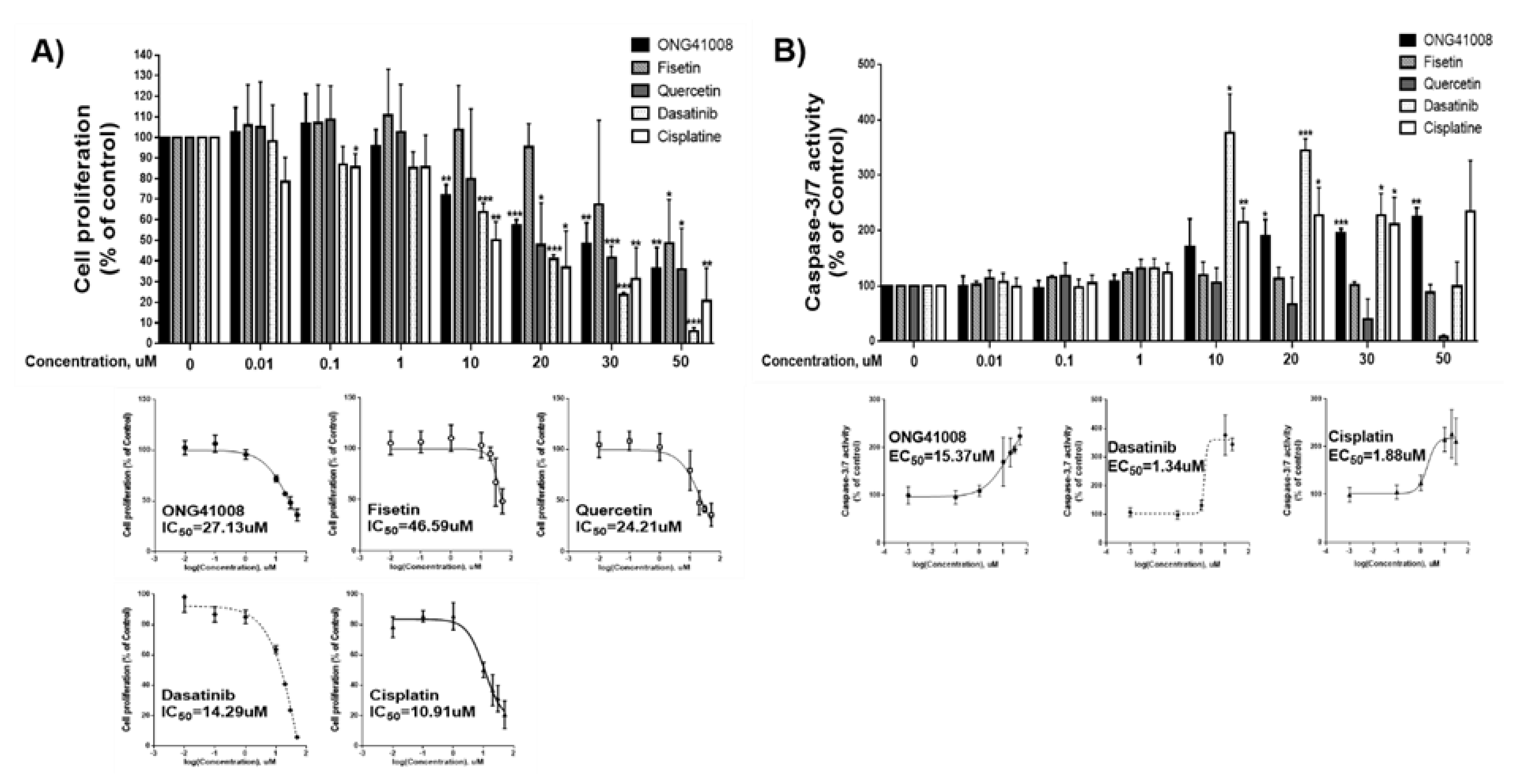
2.6. ONG41008 Is a Cancer Cell Senolytic
2.7. Biochemical Analysis of ONG41008-Mediated Cancer Cell Senolytics in A549
2.8. Recognition of the Intracellular Microenvironment by ONG41008 May Determine Strength of Cancer Cell Synolytics (CCS)
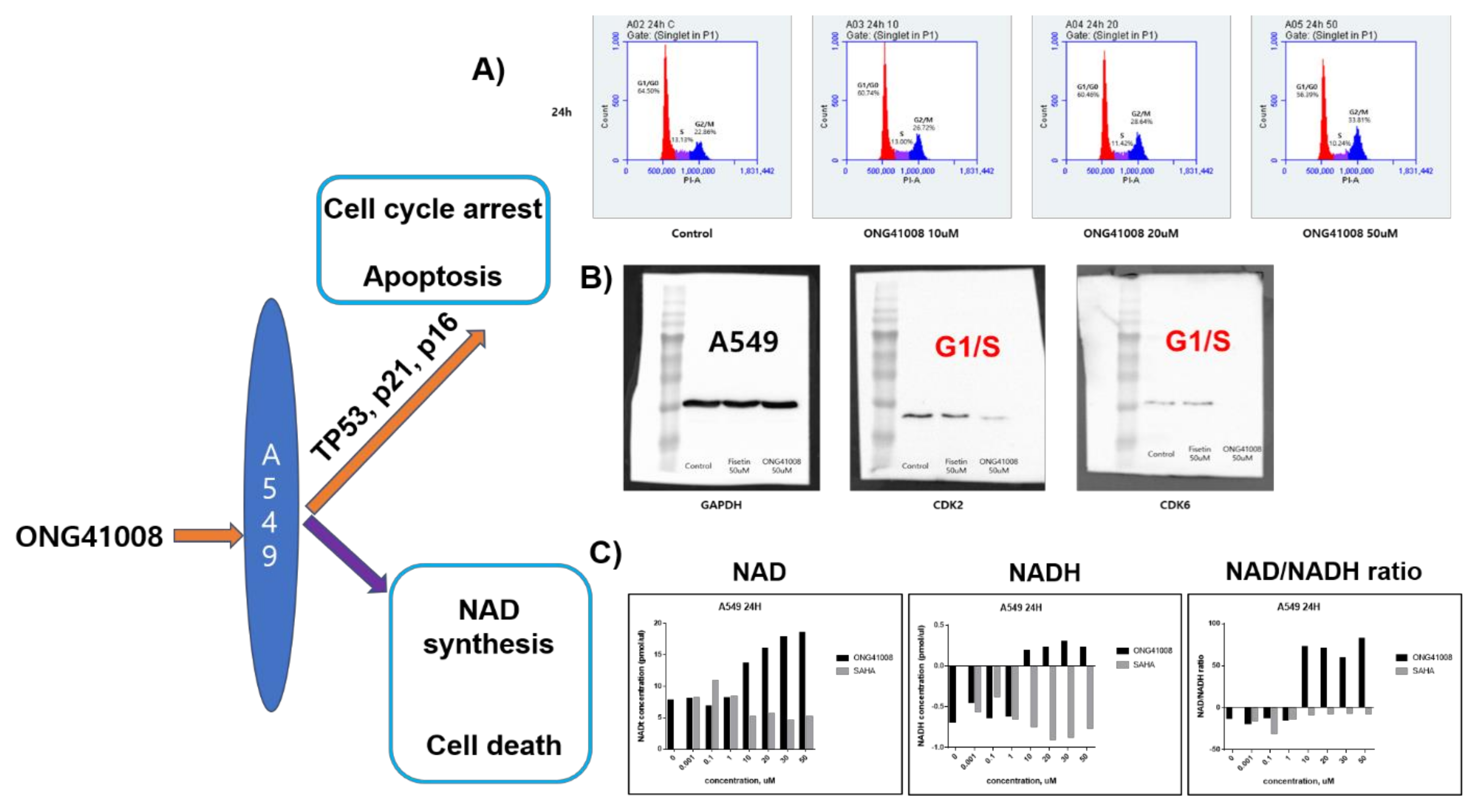
2.9. ONG41008 Induced Cell-Cycle Arrest at G2/M and Restored the NAD/NADH Ratio
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Immunocytochemistry
4.3. Western Blotting
4.4. Live Imaging
4.5. CCK-8 Assay
4.6. Caspase-3 Assay
4.7. Mitochondrial Membrane Potential Assay
4.8. LDH Assay
4.9. NAD/NADH Assay
4.10. RNA-seq, Differential Gene Expression, and Interactome Analyses
4.11. Reverse Transcriptase PCR and Real-Time PCR
5. Layman’s Summary: The Research in Context
5.1. Evidence Prior to This Study
5.2. Added Value of This Study
5.3. Implication of All the Evidence
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| CS | Chromone Scaffold |
| DHLFs | Diseased Human Lung Fibroblasts from IPF patients |
| NHLFs | Normal Human Lung Fibroblasts |
| CCS | Cancer Cell Senolytics |
| OIS | Oncogene-Induced Senescence |
| MNC | Multinucleation |
| MTMP | Mitochondrial Membrane Potential |
| NAD | Nicotinamide Adenine Dinucleotide |
| NAMPT | Nicotinamide Phosphoribosyltransferase |
References
- Wang, J.J.; Lei, K.F.; Han, F. Tumor microenvironment: Recent advances in various cancer treatments. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3855–3864. [Google Scholar] [PubMed]
- Maier, D.; Nagel, A.C.; Preiss, A. Two isoforms of the Notch antagonist Hairless are produced by differential translation initiation. Proc. Natl. Acad. Sci. USA 2002, 99, 15480–15485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallorini, M.; Cataldi, A.; di Giacomo, V. Cyclin-dependent kinase modulators and cancer therapy. BioDrugs 2012, 26, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; You, Y.; Jiang, H.; Wang, Z.Z. Epithelial-mesenchymal transition (EMT): A biological process in the development, stem cell differentiation, and tumorigenesis. J. Cell. Physiol. 2017, 232, 3261–3272. [Google Scholar] [CrossRef]
- Nowell, P.C. The clonal evolution of tumor cell populations. Science 1976, 194, 23–28. [Google Scholar] [CrossRef]
- van Meerbeeck, J.P.; Fennell, D.A.; De Ruysscher, D.K.M. Small-cell lung cancer. Lancet 2011, 378, 1741–1755. [Google Scholar] [CrossRef]
- Luengo, A.; Li, Z.; Gui, D.Y.; Sullivan, L.B.; Zagorulya, M.; Do, B.T.; Ferreira, R.; Naamati, A.; Ali, A.; Lewis, C.A.; et al. Increased demand for NAD(+) relative to ATP drives aerobic glycolysis. Mol. Cell 2021, 81, 691–707.e6. [Google Scholar] [CrossRef]
- Pavlides, S.; Tsirigos, A.; Vera, I.; Flomenberg, N.; Frank, P.G.; Casimiro, M.C.; Wang, C.; Pestell, R.G.; Martinez-Outschoorn, U.E.; Howell, A.; et al. Transcriptional evidence for the “Reverse Warburg Effect” in human breast cancer tumor stroma and metastasis: Similarities with oxidative stress, inflammation, Alzheimer’s disease, and “Neuron-Glia Metabolic Coupling”. Aging 2010, 2, 185–199. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kaur, M.; Silakari, O. Flavones: An important scaffold for medicinal chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef]
- Fischer, N.; Seo, E.J.; Efferth, T. Prevention from radiation damage by natural products. Phytomedicine 2018, 47, 192–200. [Google Scholar] [CrossRef]
- Quintão, N.L.M.; Pastor, M.V.D.; Antonialli, C.d.-S.; da Silva, G.F.; Rocha, L.W.; Berté, T.E.; de Souza, M.M.; Meyre-Silva, C.; Lucinda-Silva, R.M.; Bresolin, T.M.B.; et al. Aleurites moluccanus and its main active constituent, the flavonoid 2″-O-rhamnosylswertisin, in experimental model of rheumatoid arthritis. J. Ethnopharmacol. 2019, 235, 248–254. [Google Scholar] [CrossRef]
- Lu, Q.Q.; Chen, Y.M.; Liu, H.R.; Yan, J.Y.; Cui, P.W.; Zhang, Q.F.; Gao, X.H.; Feng, X.; Liu, Y.Z. Nitrogen-containing flavonoid and their analogs with diverse B-ring in acetylcholinesterase and butyrylcholinesterase inhibition. Drug Dev. Res. 2020, 81, 1037–1047. [Google Scholar] [CrossRef]
- Kim, H.-S.; Meang, M.K.; Ham, M.; Lee, J.Y.; Seong, B.L.; Yoon, H.-S.; Kim, I.-H.; Youn, B.-S. Discovery of a small molecule having both potent anti-fibrotic and anti-inflammatory capabilities. bioRxiv 2020, 770404. [Google Scholar] [CrossRef] [Green Version]
- Richeldi, L.; du Bois, R.M.; Raghu, G.; Azuma, A.; Brown, K.K.; Costabel, U.; Cottin, V.; Flaherty, K.R.; Hansell, D.M.; Inoue, Y.; et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N. Engl. J. Med. 2014, 370, 2071–2082. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, L.H.; de Andrade, J.A.; Zibrak, J.D.; Padilla, M.L.; Albera, C.; Nathan, S.D.; Wijsenbeek, M.S.; Stauffer, J.L.; Kirchgaessler, K.-U.; Costabel, U. Pirfenidone safety and adverse event management in idiopathic pulmonary fibrosis. Eur. Respir. Rev. 2017, 26, 170057. [Google Scholar] [CrossRef] [Green Version]
- Itahana, K.; Campisi, J.; Dimri, G.P. Mechanisms of cellular senescence in human and mouse cells. Biogerontology 2004, 5, 1–10. [Google Scholar] [CrossRef]
- Kirkland, J.L.; Tchkonia, T. Cellular Senescence: A Translational Perspective. EBioMedicine 2017, 21, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Schafer, M.J.; White, T.A.; Iijima, K.; Haak, A.J.; Ligresti, G.; Atkinson, E.J.; Oberg, A.L.; Birch, J.; Salmonowicz, H.; Zhu, Y.; et al. Cellular senescence mediates fibrotic pulmonary disease. Nat. Commun. 2017, 8, 14532. [Google Scholar] [CrossRef]
- Steinberg, M. Dasatinib: A tyrosine kinase inhibitor for the treatment of chronic myelogenous leukemia and philadelphia chromosome-positive acute lymphoblastic leukemia. Clin. Ther. 2007, 29, 2289–2308. [Google Scholar] [CrossRef]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef]
- Yousefzadeh, M.J.; Zhu, Y.; McGowan, S.J.; Angelini, L.; Fuhrmann-Stroissnigg, H.; Xu, M.; Ling, Y.Y.; Melos, K.I.; Pirtskhalava, T.; Inman, C.L.; et al. Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine 2018, 36, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Contrepois, K.; Coudereau, C.; Benayoun, B.A.; Schuler, N.; Roux, P.-F.; Bischof, O.; Courbeyrette, R.; Carvalho, C.; Thuret, J.-Y.; Ma, Z.; et al. Histone variant H2A.J accumulates in senescent cells and promotes inflammatory gene expression. Nat. Commun. 2017, 8, 14995. [Google Scholar] [CrossRef] [Green Version]
- Senderowicz, A.M. Cell cycle modulators for the treatment of lung malignancies. Clin. Lung Cancer 2003, 5, 158–168. [Google Scholar] [CrossRef]
- Carvalho, C.; L’Hôte, V.; Courbeyrette, R.; Kratassiouk, G.; Pinna, G.; Cintrat, J.C.; Denby-Wilkes, C.; Derbois, C.; Olaso, R.; Deleuze, J.F.; et al. Glucocorticoids delay RAF-induced senescence promoted by EGR. J. Cell. Sci. 2019, 132. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, J.; Bellani, M.A.; Pokharel, D.; Gichimu, J.; James, R.C.; Gali, H.; Ling, C.; Yan, Z.; Xu, D.; et al. Remodeling of Interstrand Crosslink Proximal Replisomes Is Dependent on ATR, FANCM, and FANCD. Cell Rep. 2019, 27, 1794–1808.e5. [Google Scholar] [CrossRef] [Green Version]
- Suhail, T.V.; Singh, P.; Manna, T.K. Suppression of centrosome protein TACC3 induces G1 arrest and cell death through activation of p38-p53-p21 stress signaling pathway. Eur. J. Cell Biol. 2015, 94, 90–100. [Google Scholar] [CrossRef]
- Michowski, W.; Chick, J.M.; Chu, C.; Kolodziejczyk, A.; Wang, Y.; Suski, J.M.; Abraham, B.; Anders, L.; Day, D.; Dunkl, L.M.; et al. Cdk1 Controls Global Epigenetic Landscape in Embryonic Stem Cells. Mol. Cell 2020, 78, 459–476.e13. [Google Scholar] [CrossRef]
- Dikovskaya, D.; Cole, J.J.; Mason, S.M.; Nixon, C.; Karim, S.A.; McGarry, L.; Clark, W.; Hewitt, R.N.; Sammons, M.A.; Zhu, J.; et al. Mitotic Stress Is an Integral Part of the Oncogene-Induced Senescence Program that Promotes Multinucleation and Cell Cycle Arrest. Cell Rep. 2015, 12, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.; Serrano, M. The power and the promise of oncogene-induced senescence markers. Nat. Rev. Cancer 2006, 6, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Persidis, A. Cancer multidrug resistance. Nat. Biotechnol. 1999, 17, 94–95. [Google Scholar] [CrossRef] [PubMed]
- Edgington, S.M. Conquering Multidrug Resistance in Cancer Chemotherapy. Bio/Technol. 1992, 10, 269–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of p53 in Tumor Initiation and Progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef] [PubMed]
- Buchakjian, M.R.; Kornbluth, S. The engine driving the ship: Metabolic steering of cell proliferation and death. Nat. Rev. Mol. Cell Biol. 2010, 11, 715–727. [Google Scholar] [CrossRef]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, C.; Shen, G.; Zou, B. NAD+ metabolism: Pathophysiologic mechanisms and therapeutic potential. Signal Transduct. Target. Ther. 2020, 5, 227. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Schoenfeld, A.J.; Hellmann, M.D. Acquired Resistance to Immune Checkpoint Inhibitors. Cancer Cell 2020, 37, 443–455. [Google Scholar] [CrossRef]
- Stapleton, A.E.; Walbot, V. Flavonoids can protect maize DNA from the induction of ultraviolet radiation damage. Plant Physiol. 1994, 105, 881–889. [Google Scholar] [CrossRef]
- Yan, W.; Xiaoli, L.; Guoliang, A.; Zhonghui, Z.; Di, L.; Ximeng, L.; Piye, N.; Li, C.; Lin, T. SB203580 inhibits epithelial-mesenchymal transition and pulmonary fibrosis in a rat silicosis model. Toxicol. Lett. 2016, 259, 28–34. [Google Scholar] [CrossRef]
- Longo, V.D.; Antebi, A.; Bartke, A.; Barzilai, N.; Brown-Borg, H.M.; Caruso, C.; Curiel, T.J.; De Cabo, R.; Franceschi, C.; Gems, D.; et al. Interventions to slow aging in humans: Are we ready? Aging Cell 2015, 14, 497–510. [Google Scholar] [CrossRef]
- Zhu, Y.; Doornebal, E.J.; Pirtskhalava, T.; Giorgadze, N.; Wentworth, M.; Fuhrmann-Stroissnigg, H.; Niedernhofer, L.J.; Robbins, P.D.; Tchkonia, T.; Kirkland, J.L. New agents that target senescent cells: The flavone, fisetin, and the BCL-XL inhibitors, A1331852 and A1155463. Aging 2017, 9, 955–963. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meang, M.K.; Kim, S.; Kim, I.-H.; Kim, H.-S.; Youn, B.-S. A Small Molecule That Promotes Cellular Senescence Prevents Fibrogenesis and Tumorigenesis. Int. J. Mol. Sci. 2022, 23, 6852. https://doi.org/10.3390/ijms23126852
Meang MK, Kim S, Kim I-H, Kim H-S, Youn B-S. A Small Molecule That Promotes Cellular Senescence Prevents Fibrogenesis and Tumorigenesis. International Journal of Molecular Sciences. 2022; 23(12):6852. https://doi.org/10.3390/ijms23126852
Chicago/Turabian StyleMeang, Moon Kee, Saesbyeol Kim, Ik-Hwan Kim, Han-Soo Kim, and Byung-Soo Youn. 2022. "A Small Molecule That Promotes Cellular Senescence Prevents Fibrogenesis and Tumorigenesis" International Journal of Molecular Sciences 23, no. 12: 6852. https://doi.org/10.3390/ijms23126852
APA StyleMeang, M. K., Kim, S., Kim, I.-H., Kim, H.-S., & Youn, B.-S. (2022). A Small Molecule That Promotes Cellular Senescence Prevents Fibrogenesis and Tumorigenesis. International Journal of Molecular Sciences, 23(12), 6852. https://doi.org/10.3390/ijms23126852