
Genome-Wide Identification and Characterization of Growth Regulatory Factor Family Genes in Medicago

Abstract
:1. Introduction
2. Results
2.1. Identification of GRF Genes in the M. sativa and M. truncatula Genome
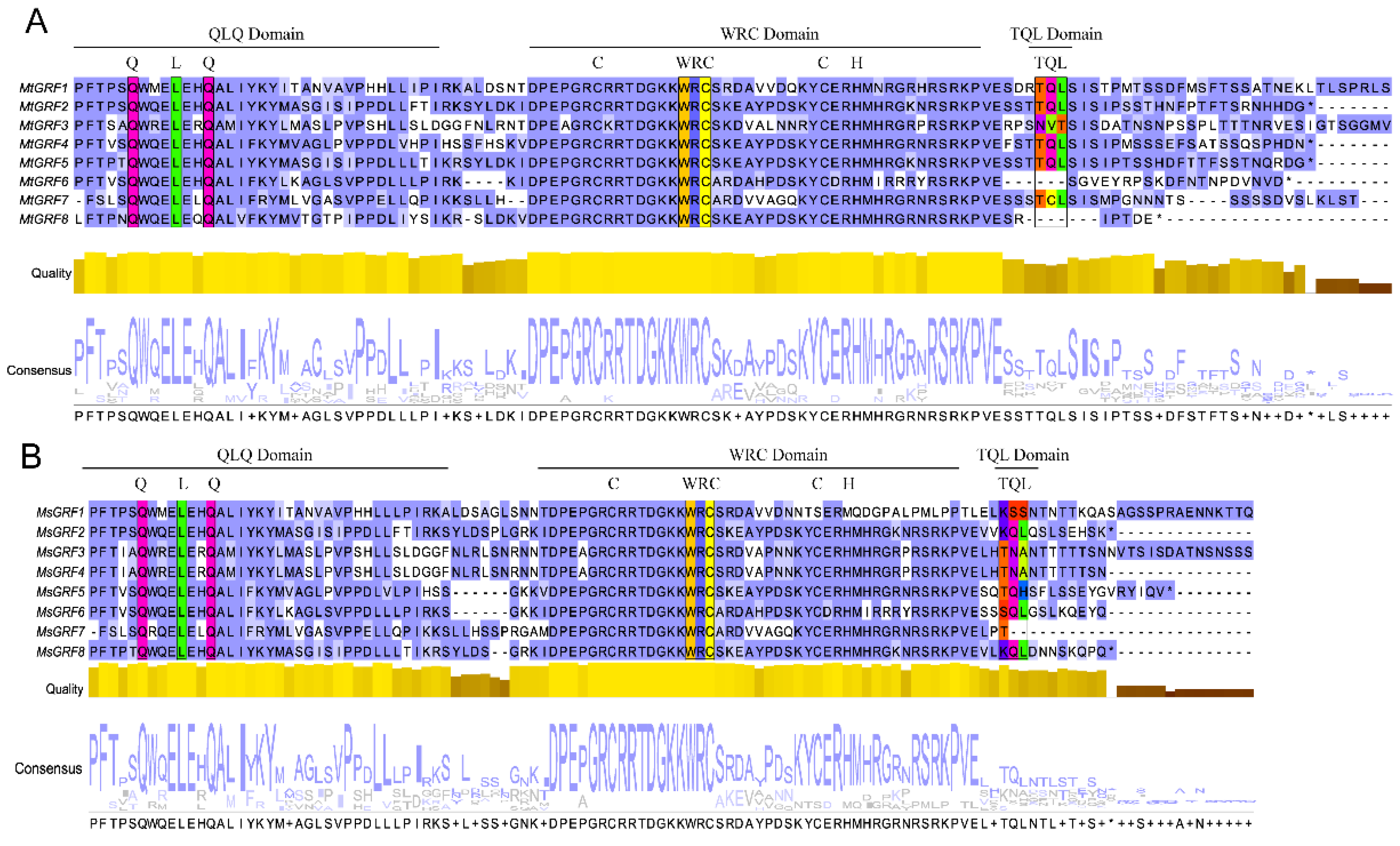
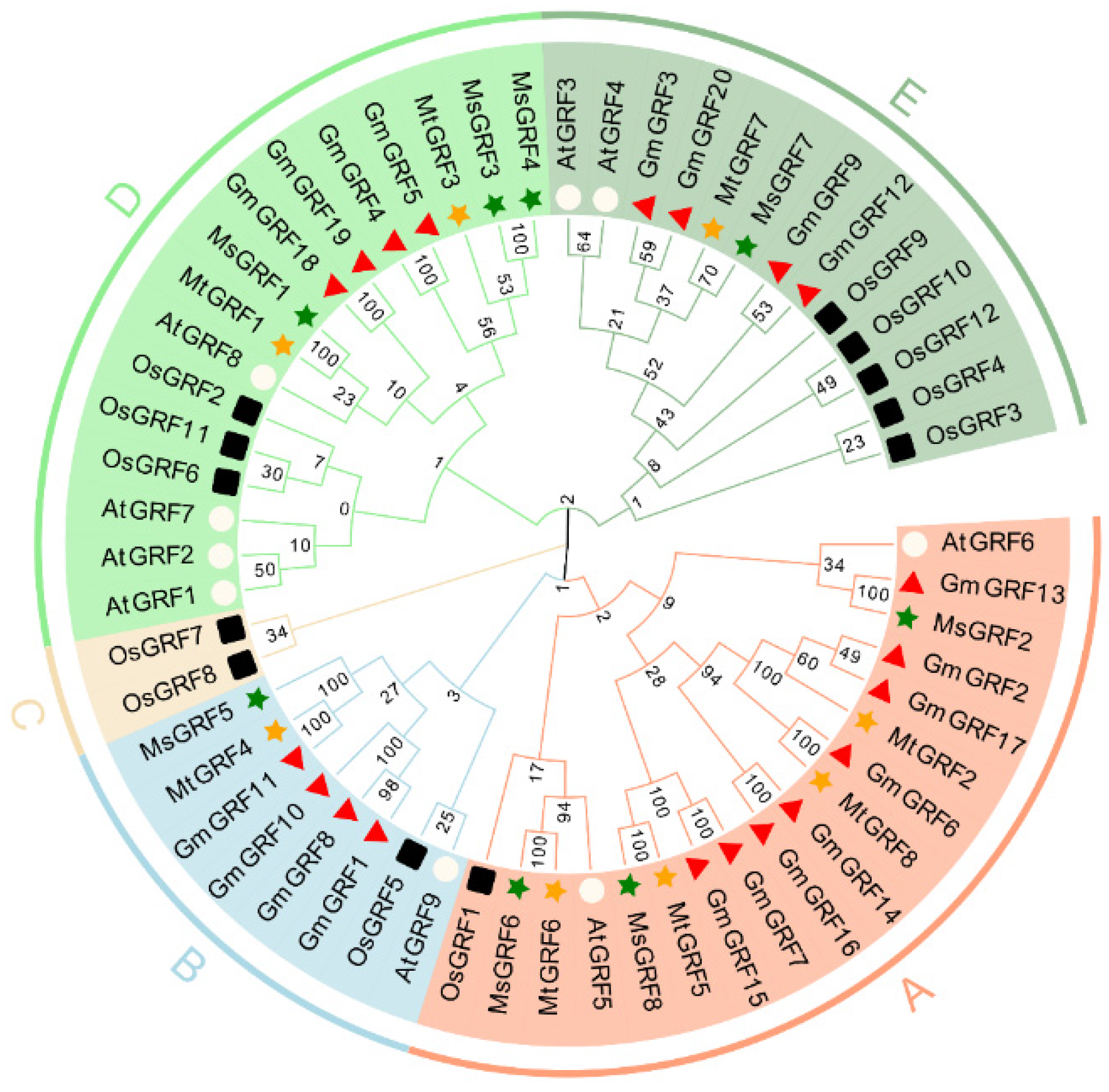
2.2. Multiple Sequence Alignment, Phylogenetic Analysis and Classification of GRF Genes in Medicago
2.3. Analyses of Conserved Motif and Gene Structure
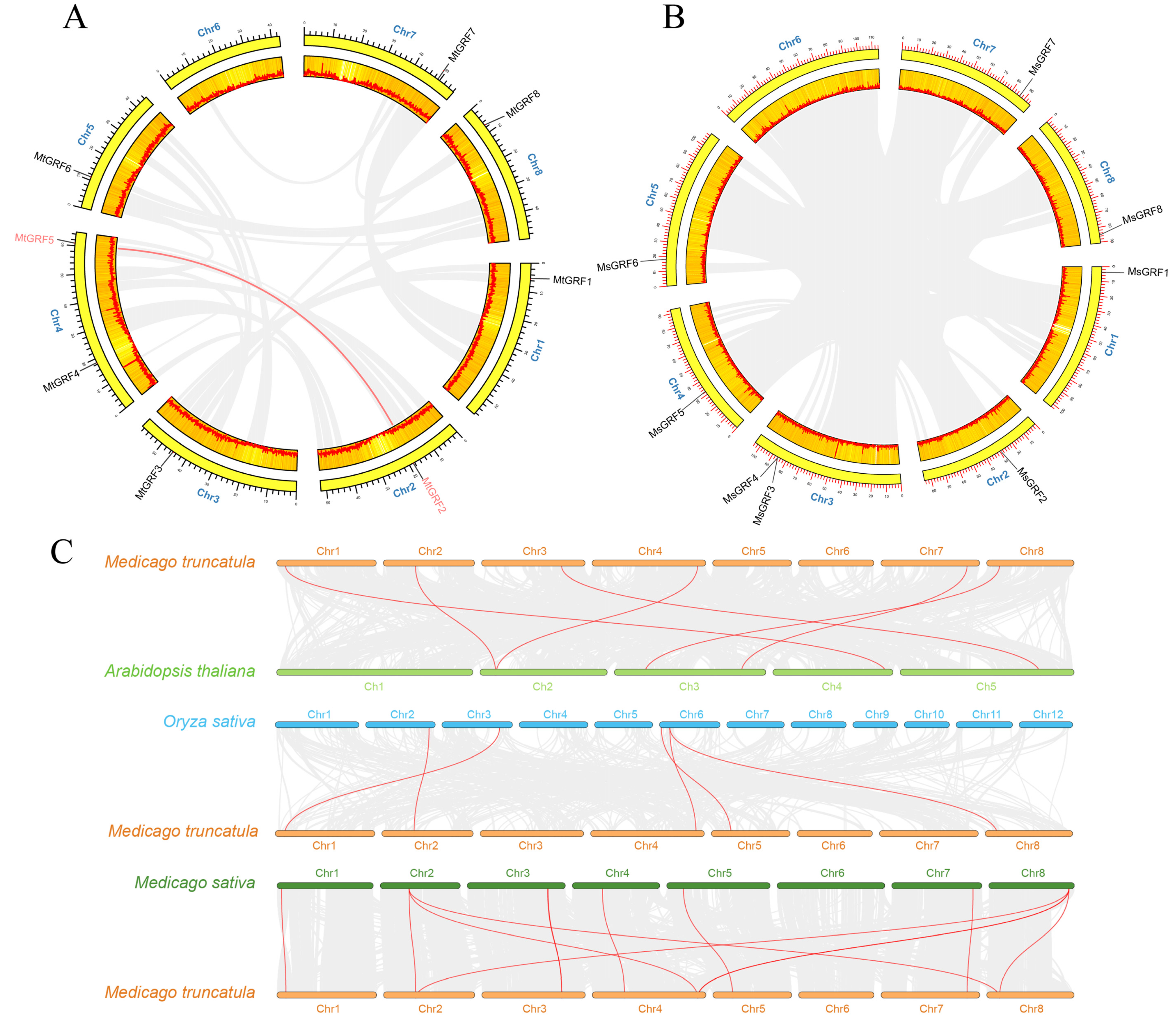
2.4. Analysis of Chromosome Location and Collinearity of GRF Genes
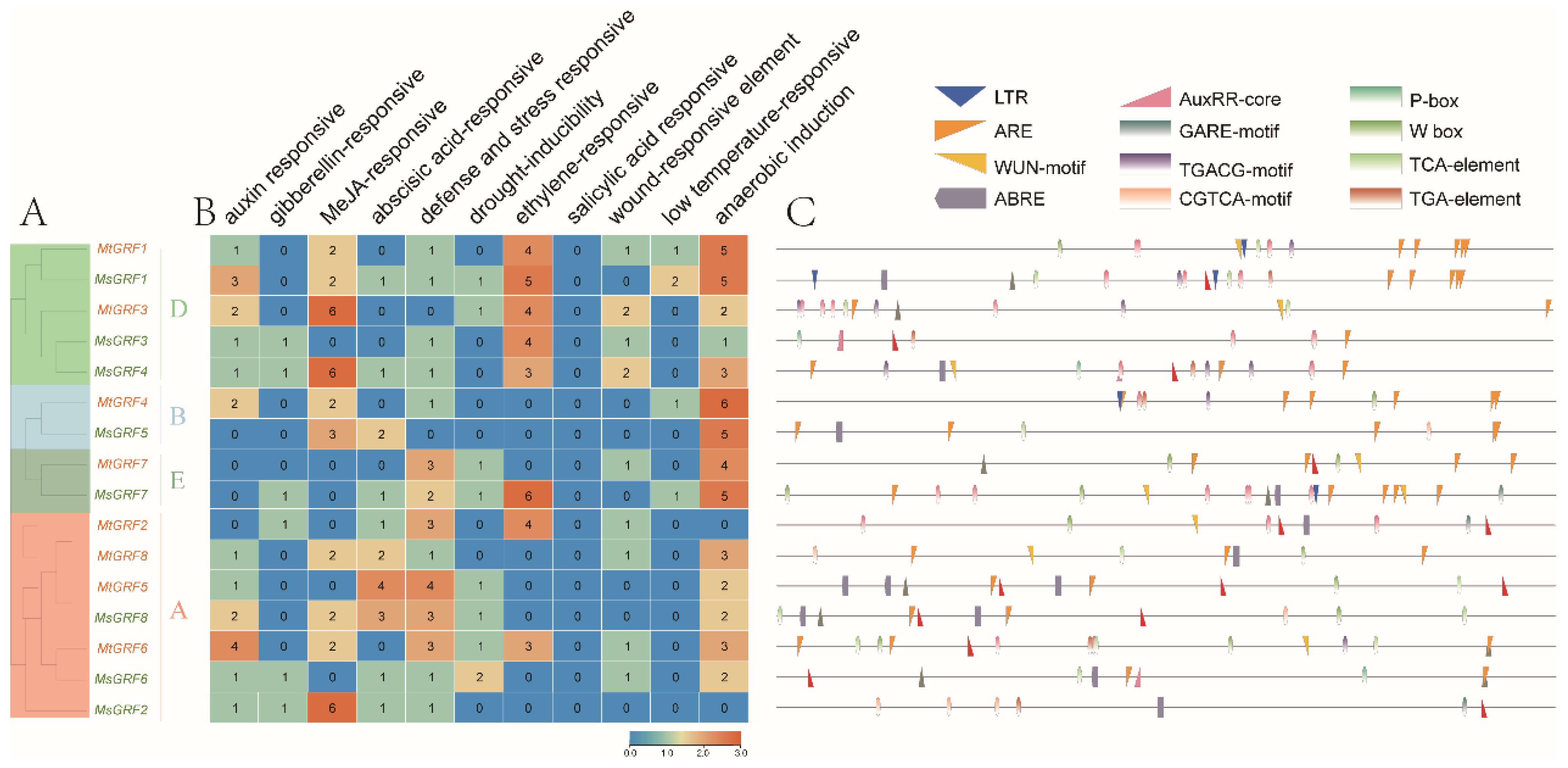
2.5. Analysis of cis-Acting Element of GRF Genes
2.6. Expression Profiles of GRF Genes in Different Tissues
2.7. Expression Profiles of MtGRF Genes under Stress Treatments
2.8. Expression Profiles of MsGRF Genes under Stress Treatments
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Identification of GRF Genes in the Medicago Genome
5.2. Analyses on Sequence and Structures of the Medicago GRF Genes
5.3. Phylogenetic Analysis and Classification of GRF Genes
5.4. Analyses of Chromosome Location and Collinearity of GRF Genes
5.5. Analyses of cis-Acting Elements and Location of GRF Genes in Medicago
5.6. Analysis of Expression Level of GRF Genes
5.7. Plant Materials and Treatments
5.8. Analysis of Gene Expression by qPCR
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kim, J.H.; Tsukaya, H. Regulation of plant growth and development by the GROWTH-REGULATING FACTOR and GRF-INTERACTING FACTOR duo. J. Exp. Bot. 2015, 66, 6093–6107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaap, E.; Kim, J.H.; Kend, H. A novel gibberellin-induced gene from rice and its potential regulatory role in stem growth. Plant Physiol. 2000, 122, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Choi, D.; Kende, H. The AtGRF family of putative transcription factors is involved in leaf and cotyledon growth in Arabidopsis. Plant J. 2003, 36, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kende, H. A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morphology in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 13374–13379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.S.; Kim, J.H.; Kende, H. Whole genome analysis of the OsGRF gene family encoding plantspecifc putative transcription activators in rice (Oryza sativa L.). Plant Cell Physiol. 2004, 45, 897–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Guo, S.; Xu, Y.; Li, C.; Zhang, Z.; Zhang, D.; Xu, S.; Zhang, C.; Chong, K. OsmiR396d-regulated OsGRFs function in floral organogenesis in rice through binding to their targets OsJMJ706 and OsCR4. Plant Physiol. 2014, 165, 160–174. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Rice, J.H.; Chen, N.; Baum, T.J.; Hewezi, T. Synchronization of developmental processes and defense signaling by growth regulating transcription factors. PLoS ONE 2014, 9, e98477. [Google Scholar] [CrossRef]
- Zhang, D.F.; Li, B.; Jia, G.Q.; Zhang, T.F.; Dai, J.R.; Li, J.S.; Wang, S.C. Isolation and characterization of genes encoding GRF transcription factors and GIF transcriptional coactivators in Maize (Zea mays L.). Plant Sci. 2008, 175, 809–817. [Google Scholar] [CrossRef]
- Bazin, J.; Khan, G.A.; Combier, J.P.; Bustos-Sanmamed, P.; Debernardi, J.M.; Rodriguez, R.; Sorin, C.; Palatnik, J.; Hartmann, C.; Crespi, M.; et al. miR396 affects mycorrhization and root meristem activity in the legume Medicago truncatula. Plant J. 2013, 74, 920–934. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, D.; Xue, M.; Qian, J.; He, Y.; Wang, S. Overexpression of the maize GRF10, an endogenous truncated growth-regulating factor protein, leads to reduction in leaf size and plant height. J. Integr. Plant Biol. 2014, 56, 1053–1063. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Mecchia, M.A.; Vercruyssen, L.; Smaczniak, C.; Kaufmann, K.; Inze, D.; Rodriguez, R.E.; Palatnik, J.F. Post-transcriptional control of GRF transcription factors by microRNA miR396 and GIF co-activator affects leaf size and longevity. Plant J. 2014, 79, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, G.; Kim, G.T.; Tsukaya, H. The transcription factor AtGRF5 and the transcription coactivator AN3 regulate cell proliferation in leaf primordia of Arabidopsis thaliana. Plant J. 2005, 43, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, B.H. GROWTH-REGULATING FACTOR4 of Arabidopsis thaliana is required for development of leaves, cotyledons, and shoot apical meristem. J. Plant Biol. 2006, 49, 463–468. [Google Scholar] [CrossRef]
- Kuijt, S.J.; Greco, R.; Agalou, A.; Shao, J.; Hoen, C.C.; Overnas, E.; Osnato, M.; Curiale, S.; Meynard, D.; van Gulik, R.; et al. Interaction between the GRF and KNOX families of transcription factors. Plant Physiol. 2014, 164, 1952–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaba, S.; Fukumoto, M.; Honda, C.; Yamaguchi, I.; Sakamoto, T.; Kano-Murakami, Y. Decreased GA1 content caused by the overexpression of OSH1 is accompanied by suppression of GA 20-oxidase gene expression. Plant Physiol. 1998, 117, 1179–1184. [Google Scholar] [CrossRef] [Green Version]
- Bolduc, N.; Hake, S. The maize transcription factor KNOTTED1 directly regulates the gibberellin catabolism gene ga2ox1. Plant Cell 2009, 21, 1647–1658. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; He, H.; Li, Y.; Wang, F.; Yu, D. Molecular mechanism of microRNA396 mediating pistil development in Arabidopsis. Plant Physiol. 2014, 164, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Schommer, C.; Debernardi, J.M.; Bresso, E.G.; Rodriguez, R.E.; Palatnik, J.F. Repression of cell proliferation by miR319-regulated TCP4. Mol. Plant 2014, 7, 1533–1544. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.E.; Mecchia, M.A.; Debernardi, J.M.; Schommer, C.; Weigel, D.; Palatnik, J.F. Control of cell proliferation in Arabidopsis thaliana by microRNA miR396. Development 2010, 137, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajoro, A.; Madrigal, P.; Muiño, J.M.; Matus, J.T.; Jin, J.; Mecchia, M.A.; Debernardi, J.M.; Palatnik, J.F.; Balazadeh, S.; Arif, M.; et al. Dynamics of chromatin accessibility and gene regulation by MADS domain transcription factors in flower development. Genome Biol. 2014, 15, R41. [Google Scholar] [CrossRef]
- Nelissen, H.; Eeckhout, D.; Demuynck, K.; Persiau, G.; Walton, A.; van Bel, M.; Vervoort, M.; Candaele, J.; De Block, J.; Aesaert, S.; et al. Dynamic Changes in ANGUSTIFOLIA3 Complex Composition Reveal a Growth Regulatory Mechanism in the Maize Leaf. Plant Cell 2015, 27, 1605–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Mizoi, J.; Kidokoro, S.; Maruyama, K.; Nakajima, J.; Nakashima, K.; Mitsuda, N.; Takiguchi, Y.; Ohme-Takagi, M.; Kondou, Y.; et al. Arabidopsis growth-regulating factor7 functions as a transcriptional repressor of abscisic acid- and osmotic stress-responsive genes, including DREB2A. Plant Cell 2012, 24, 3393–3405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Butselaar, T.; Van den Ackerveken, G. Salicylic Acid Steers the Growth-Immunity Tradeoff. Trends Plant Sci. 2020, 25, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.B.; Caldwell, M.M. The scale of nutrient heterogeneity around individual plants and its quantification with geostatistics. Ecol. Soc. Am. 1993, 74, 612–614. [Google Scholar] [CrossRef]
- Li, X.; Dhaubhadel, S. Soybean 14-3-3 gene family: Identification and molecular characterization. Planta 2011, 233, 569–582. [Google Scholar] [CrossRef]
- Xu, W.F.; Shi, W.M. Expression profiling of the 14-3-3 gene family in response to salt stress and potassium and iron deficiencies in young tomato (Solanum lycopersicum) roots: Analysis by real-time RT-PCR. Ann. Bot. 2006, 98, 965–974. [Google Scholar] [CrossRef] [Green Version]
- Elliott, F.C.; Johnson, I.J.; Schonhorst, M.H. Breeding for forage yield and quality. In Alfalfa Science and Technology Agronomy Monographs; American Society of Agronomy: Madison, WI, USA, 1972. [Google Scholar]
- Li, X.H.; Brummer, E.C. Applied Genetics and Genomics in Alfalfa Breeding. Agronomy 2012, 2, 40–61. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.L. The evolution of functionally novel proteins after gene duplication. Proc. Biol. Sci. 1994, 256, 119–124. [Google Scholar]
- Vercruyssen, L.; Tognetti, V.B.; Gonzalez, N.; Van Dingenen, J.; De Milde, L.; Bielach, A.; De Rycke, R.; Van Breusegem, F.; Inze, D. GROWTH REGULATING FACTOR5 stimulates Arabidopsis chloroplast division, photosynthesis, and leaf longevity. Plant Physiol. 2015, 167, 817–832. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.Q.; Jian, H.J.; Yang, B.; Lu, K.; Zhang, A.X.; Liu, P.; Li, J.N. Genome-wide analysis and expression profiling of the GRF gene family in oilseed rape (Brassica napus L.). Gene 2017, 620, 36–45. [Google Scholar] [CrossRef]
- Vision, T.J.; Brown, D.G.; Tanksley, S.D. The origins of genomic duplications in Arabidopsis. Science 2000, 290, 2114–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, M.C.; Rhoades, J.D.; Draper, J.H.; Scardaci, S.C.; Spyres, M.D. Assessment of salt tolerance in rice cultivars in response to salinity problems in california. Crop Ecol. Prod. Manag. 1998, 38, 394–398. [Google Scholar] [CrossRef]
- Gao, F.; Wang, K.; Liu, Y.; Chen, Y.; Chen, P.; Shi, Z.; Luo, J.; Jiang, D.; Fan, F.; Zhu, Y.; et al. Blocking miR396 increases rice yield by shaping inflorescence architecture. Nat. Plants 2015, 2, 15196. [Google Scholar] [CrossRef] [PubMed]
- Omidbakhshfard, M.A.; Fujikura, U.; Olas, J.J.; Xue, G.P.; Balazadeh, S.; Mueller-Roeber, B. GROWTH-REGULATING FACTOR 9 negatively regulates arabidopsis leaf growth by controlling ORG3 and restricting cell proliferation in leaf primordia. PLoS Genet. 2018, 14, e1007484. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Molelular Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Tang, H.B.; Debarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.H.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.F.; Ma, L.; Jiang, W.B.; Yao, Y.; Tang, Y.H.; Pang, Y.Z. Comprehensive identification and characterization of abiotic stress and hormone responsive glycosyl hydrolase family 1 genes in Medicago truncatula. Plant Physiol. Biochem. 2021, 158, 21–33. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.X.; Oono, Y.; Zhu, J.; He, X.J.; Wu, J.M.; Iida, K.; Lu, X.Y.; Cui, X.; Jin, H.; Zhu, J.K. The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | TIGR Locus | Start Site | End Site | Homologous Gene | PI | MW (kDa) | Protein Length | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| MtGRF1 | MtrunA17Chr1g0152191 | 4965222 | 4969855 | MsG0180000350.01.T01 | 7.07 | 70.74 | 654 | Nuclear |
| MtGRF2 | MtrunA17Chr2g0300511 | 18298975 | 18301839 | MsG0280008385.01.T01 | 8.75 | 40.80 | 357 | Nuclear |
| MtGRF3 | MtrunA17Chr3g0127641 | 45643490 | 45646030 | MsG0380016593.01.T01 | 7.8 | 58.33 | 540 | Extracellular |
| MtGRF4 | MtrunA17Chr4g0021261 | 18408853 | 18413110 | MsG0480020006.01.T01 | 7.72 | 37.22 | 338 | Nuclear |
| MtGRF5 | MtrunA17Chr4g0070591 | 60177320 | 60179812 | MsG0880047345.01.T01 | 7.29 | 42.01 | 369 | Nuclear |
| MtGRF6 | MtrunA17Chr5g0409471 | 11229054 | 11232562 | MsG0580025368.01.T01 | 9.03 | 36.40 | 325 | Extracellular |
| MtGRF7 | MtrunA17Chr7g0265931 | 49112735 | 49115556 | MsG0780041090.01.T01 | 8.8 | 42.42 | 385 | Nuclear |
| MtGRF8 | MtrunA17Chr8g0343881 | 7194250 | 7196469 | MsG0880047345.01.T01 | 8.39 | 39.89 | 349 | Nuclear |
| MsGRF1 | MsG0180000350.01.T01 | 4753503 | 4757720 | MtrunA17Chr1g0152191 | 6.28 | 56.20 | 516 | Nuclear |
| MsGRF2 | MsG0280008385.01.T01 | 30489028 | 30491429 | MtrunA17Chr2g0300511 | 8.43 | 41.97 | 369 | Nuclear |
| MsGRF3 | MsG0380016593.01.T01 | 85488062 | 85491005 | MtrunA17Chr3g0127641 | 7.32 | 55.45 | 513 | Extracellular |
| MsGRF4 | MsG0380016639.01.T01 | 85977548 | 85980455 | MtrunA17Chr3g0127641 | 7.81 | 50.86 | 463 | Extracellular |
| MsGRF5 | MsG0480020006.01.T01 | 31814795 | 31818355 | MtrunA17Chr4g0021261 | 9.58 | 34.29 | 305 | Nuclear |
| MsGRF6 | MsG0580025368.01.T01 | 17548590 | 17551843 | MtrunA17Chr5g0409471 | 8.97 | 37.23 | 332 | Extracellular |
| MsGRF7 | MsG0780041090.01.T01 | 86436809 | 86436865 | MtrunA17Chr7g0265931 | 10.25 | 25.36 | 224 | Nuclear |
| MsGRF8 | MsG0880047345.01.T01 | 85029014 | 85030631 | MtrunA17Chr4g0070591 | 7.29 | 42.14 | 370 | Nuclear |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, W.; Yang, J.; Li, Q.; Su, Q.; Yi, D.; Pang, Y. Genome-Wide Identification and Characterization of Growth Regulatory Factor Family Genes in Medicago. Int. J. Mol. Sci. 2022, 23, 6905. https://doi.org/10.3390/ijms23136905
Du W, Yang J, Li Q, Su Q, Yi D, Pang Y. Genome-Wide Identification and Characterization of Growth Regulatory Factor Family Genes in Medicago. International Journal of Molecular Sciences. 2022; 23(13):6905. https://doi.org/10.3390/ijms23136905
Chicago/Turabian StyleDu, Wenxuan, Junfeng Yang, Qian Li, Qian Su, Dengxia Yi, and Yongzhen Pang. 2022. "Genome-Wide Identification and Characterization of Growth Regulatory Factor Family Genes in Medicago" International Journal of Molecular Sciences 23, no. 13: 6905. https://doi.org/10.3390/ijms23136905
APA StyleDu, W., Yang, J., Li, Q., Su, Q., Yi, D., & Pang, Y. (2022). Genome-Wide Identification and Characterization of Growth Regulatory Factor Family Genes in Medicago. International Journal of Molecular Sciences, 23(13), 6905. https://doi.org/10.3390/ijms23136905