
miR-1183 Is a Key Marker of Remodeling upon Stretch and Tachycardia in Human Myocardium

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
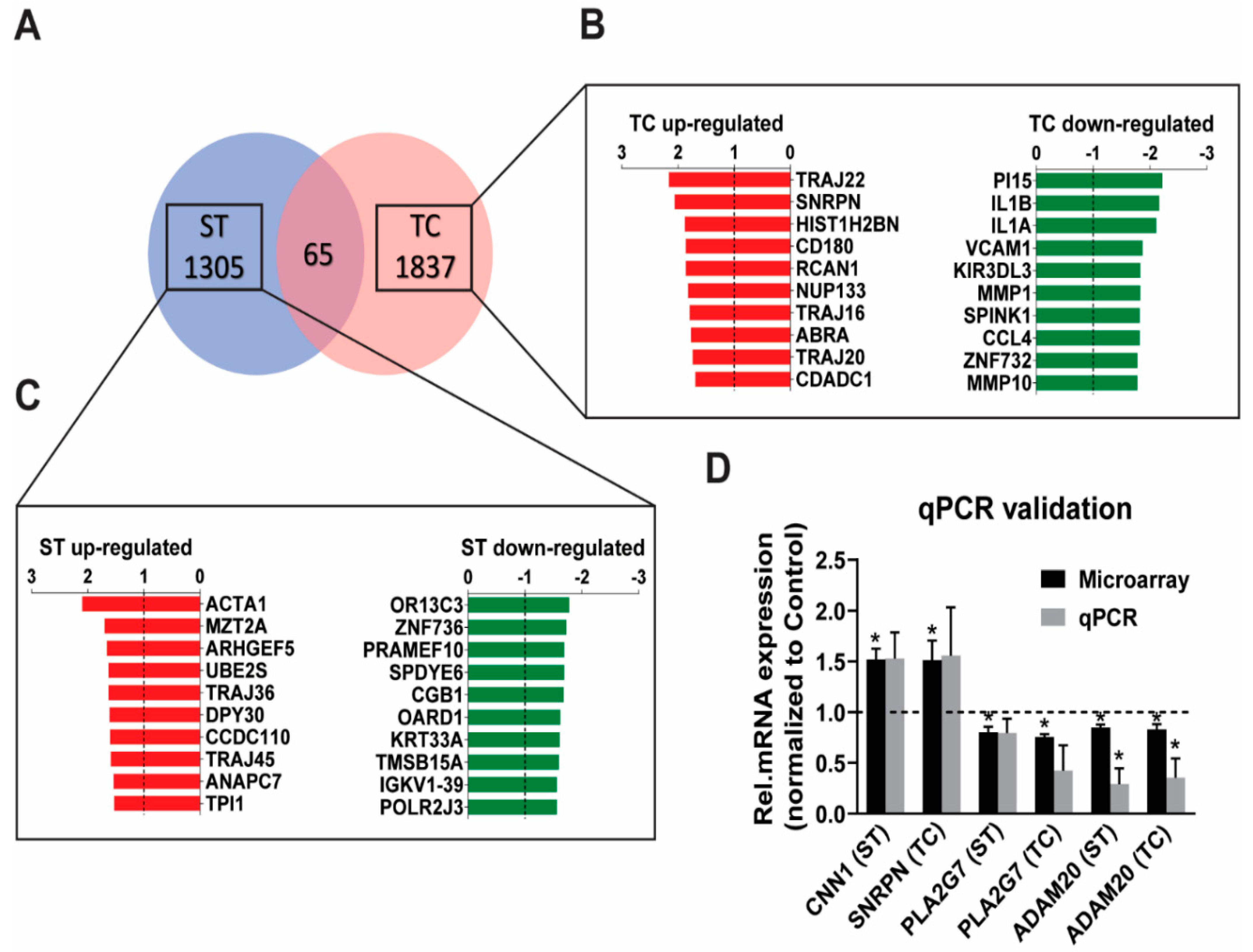
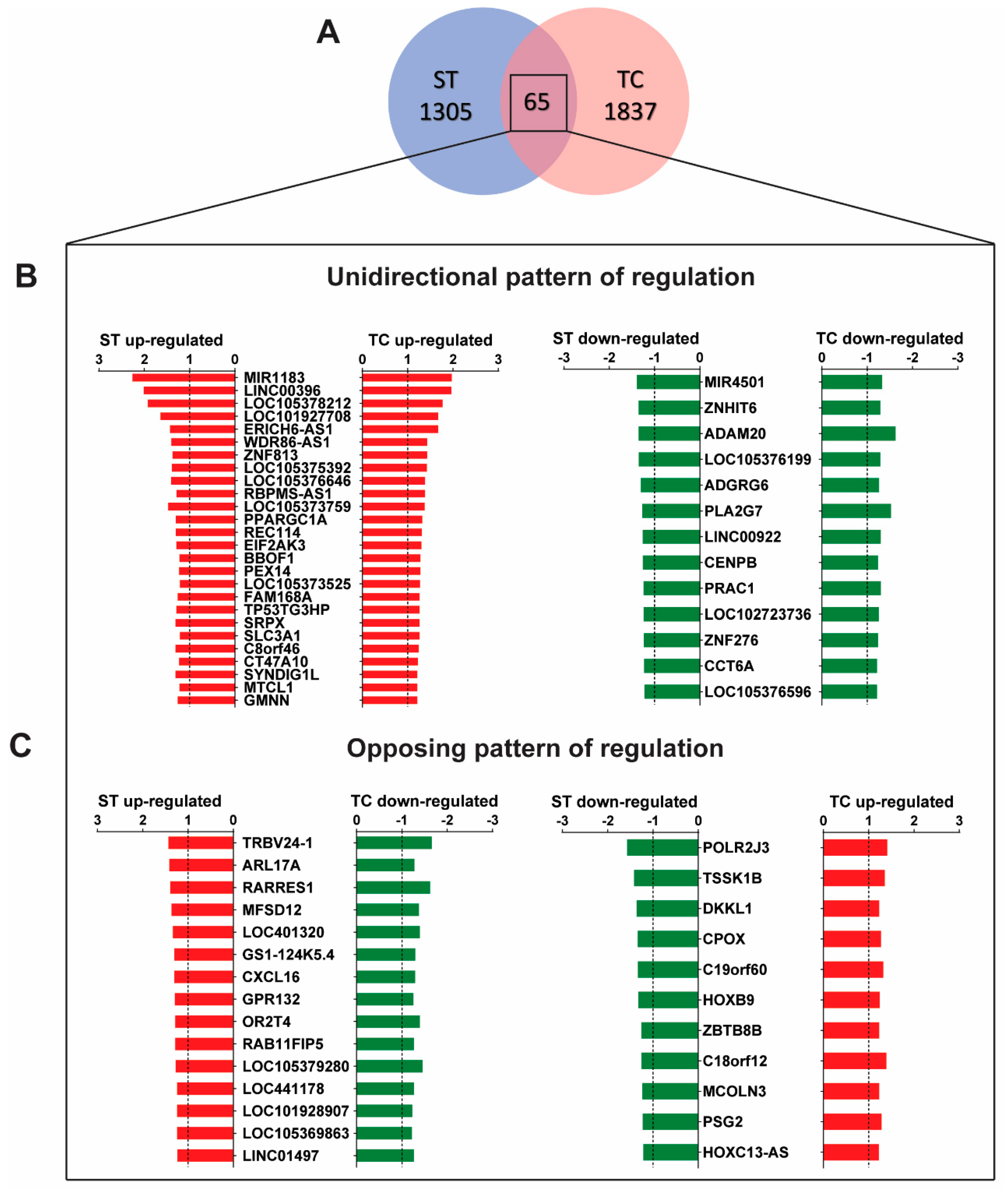
2.1. Stretch and Tachycardia Induced Differential and Common Patterns of Gene Expression
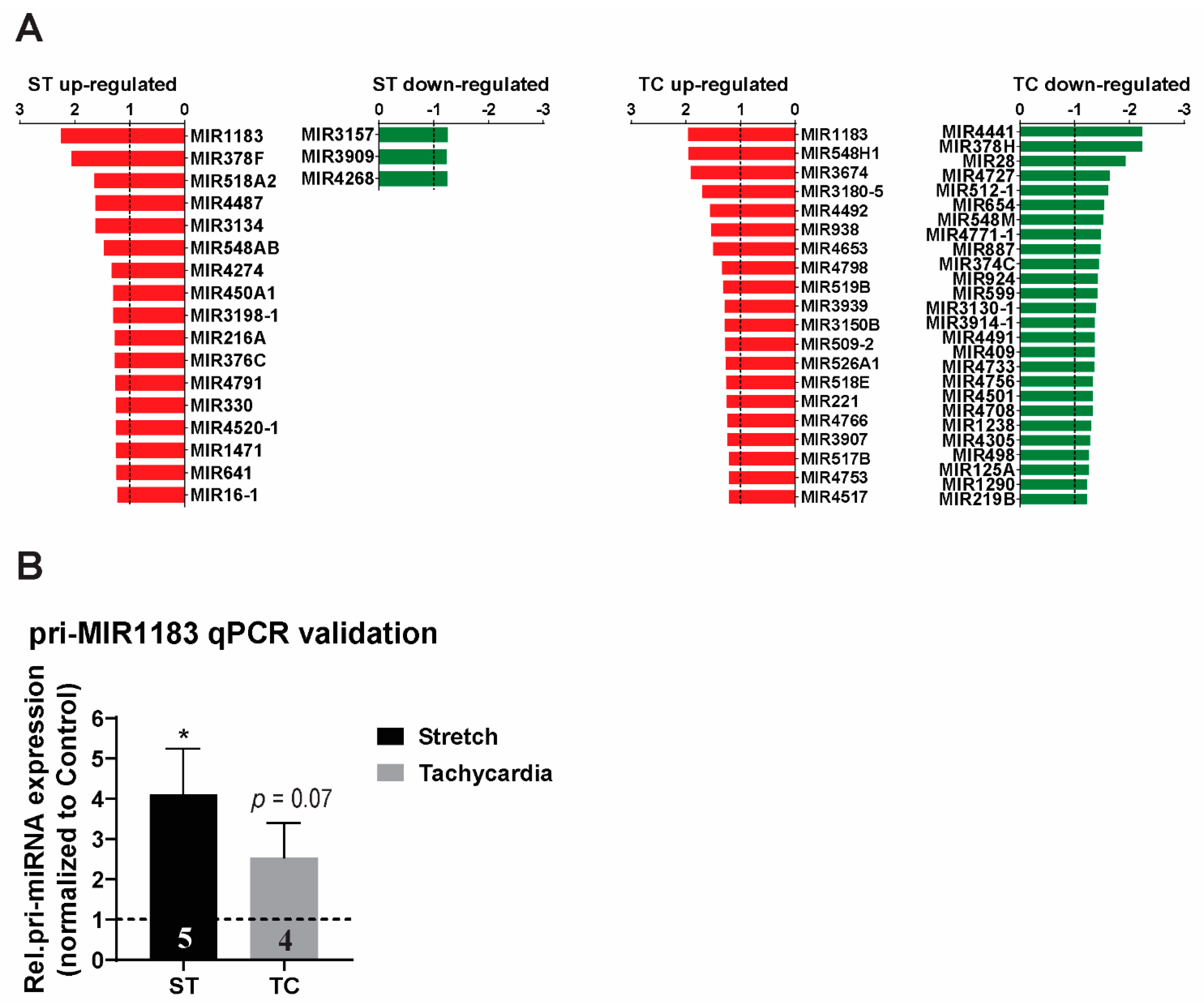
2.2. MicroRNA Expression Profile and Detection of miR-1183 Encoding Gene as the Strongest and Commonly Regulated Transcript
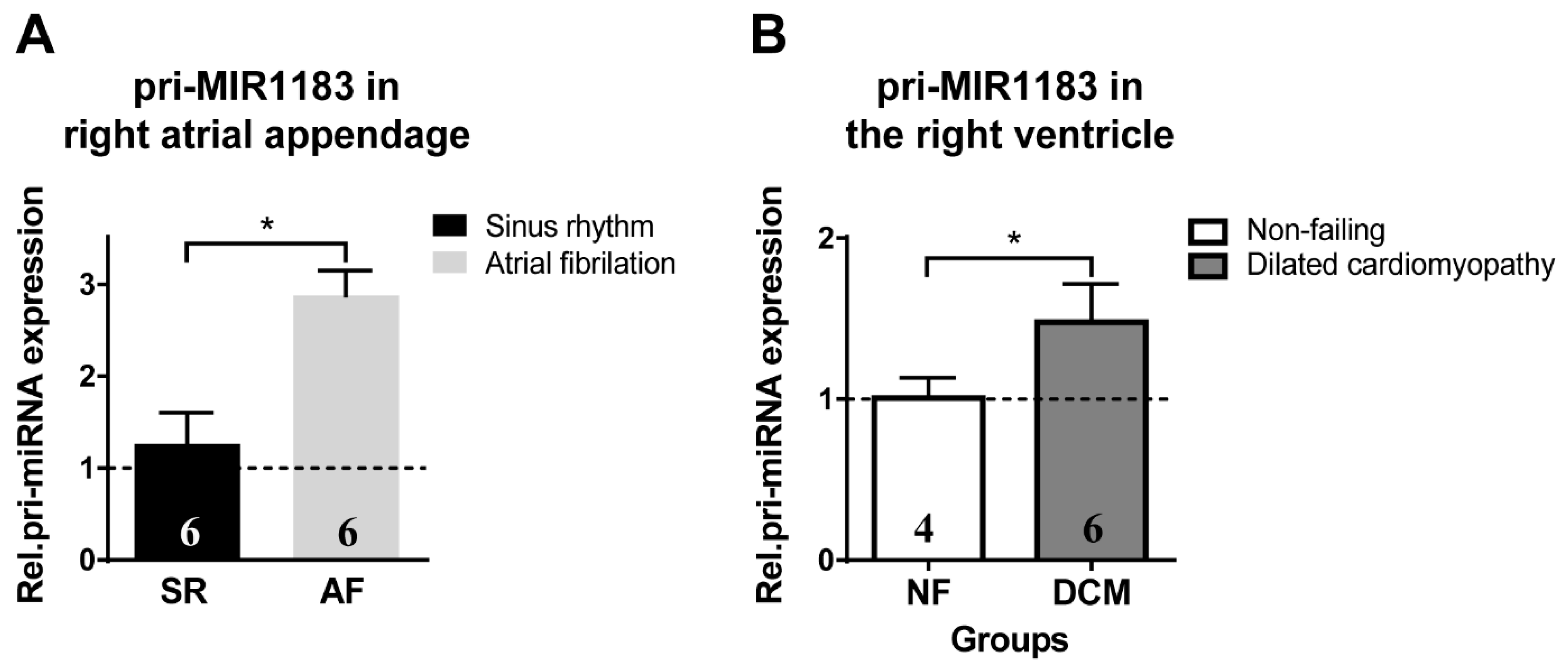
2.3. miR-1183 as a Biomarker in Human Cardiac Tissue
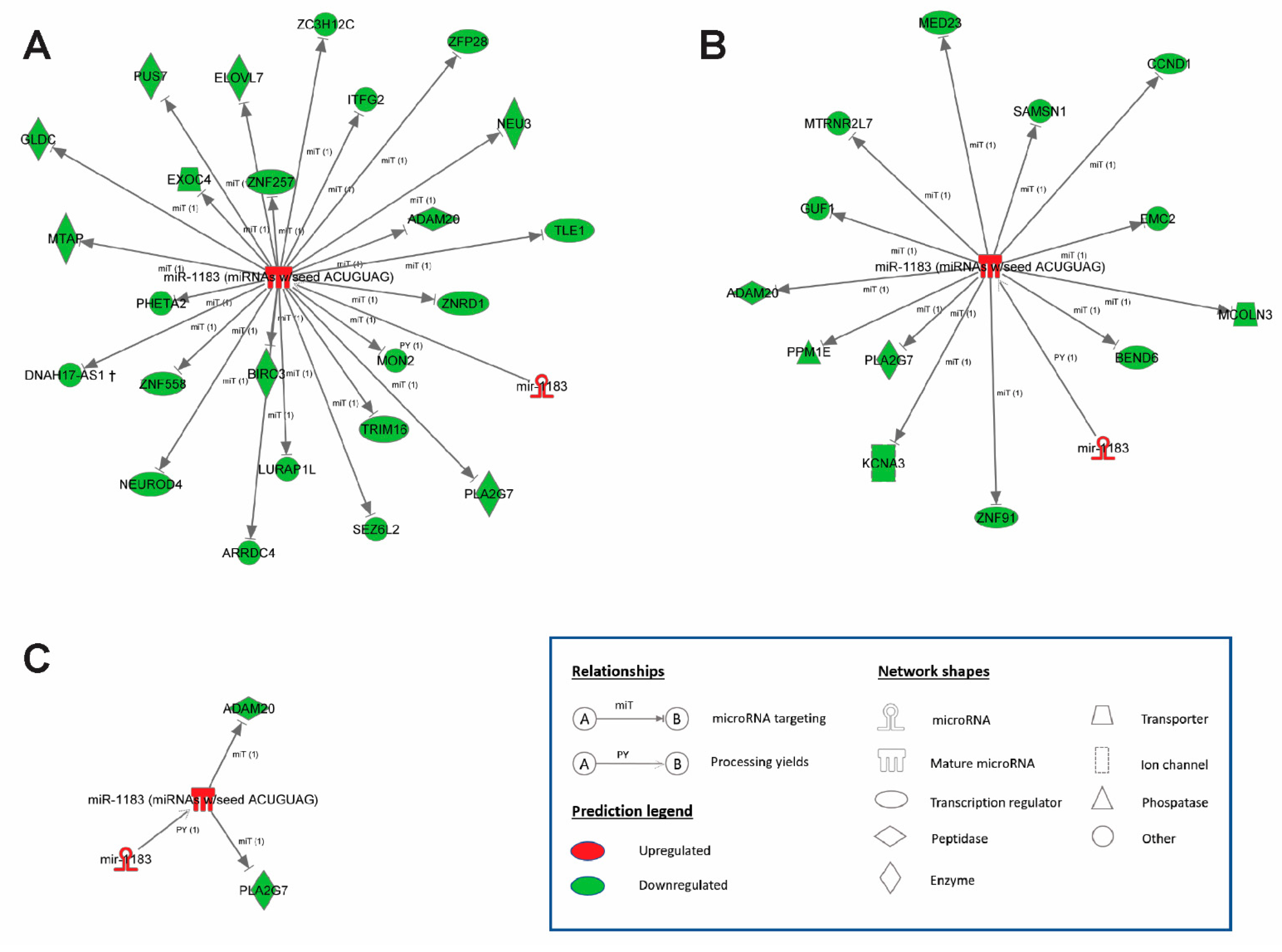
2.4. Bioinformatics Prediction of miR-1183 Downstream Targets
3. Discussion
3.1. Microarray Gene Expression Uncovers Independent Stretch and Tachycardia Gene Regulation
3.2. MicroRNA Regulation in Stretch Versus Tachycardia Induced Remodeling
3.3. miR-1183 as a Novel Marker in Atrial and Ventricular Remodeling
3.4. Upstream Regulators of miR-1183
3.5. Downstream Targets of miR-1183 and Cardiac Functional Effects
4. Material and Methods
4.1. Preparation, Processing, and Stretch and Tachycardia Induction in Human Myocardium
4.2. RNA Isolation
4.3. Microarray Hybridization
4.4. Quantitative Real-Time PCR (qPCR)
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corradi, D.; Callegari, S.; Maestri, R.; Benussi, S.; Alfieri, O. Structural remodeling in atrial fibrillation. Nat. Clin. Pract. Cardiovasc. Med. 2008, 5, 782–796. [Google Scholar] [CrossRef] [PubMed]
- Greiser, M.; Lederer, W.J.; Schotten, U. Alterations of atrial Ca2+ handling as cause and consequence of atrial fibrillation. Cardiovasc. Res. 2011, 89, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Chen, Q.; Ma, S. Left atrial fibrosis in atrial fibrillation: Mechanisms, clinical evaluation and management. J. Cell Mol. Med. 2021, 25, 2764–2775. [Google Scholar] [CrossRef]
- Al Ghamdi, B.; Hassan, W. Atrial Remodeling And Atrial Fibrillation: Mechanistic Interactions And Clinical Implications. J. Atr. Fibrillation 2009, 2, 125. [Google Scholar] [PubMed]
- Nattel, S.; Shiroshita-Takeshita, A.; Cardin, S.; Pelletier, P. Mechanisms of atrial remodeling and clinical relevance. Curr. Opin. Cardiol. 2005, 20, 21–25. [Google Scholar] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Suzuki, R.; Morishima, M.; Nakada, C.; Miyamoto, S.; Ono, K. Manifestations of gene expression profiles in human right atrial myocardium caused by mechanical stretch. Heart Vessel. 2021, 36, 577–588. [Google Scholar] [CrossRef]
- Joureau, B.; de Winter, J.M.; Conijn, S.; Bogaards, S.J.P.; Kovacevic, I.; Kalganov, A.; Persson, M.; Lindqvist, J.; Stienen, G.J.M.; Irving, T.C.; et al. Dysfunctional sarcomere contractility contributes to muscle weakness in ACTA1-related nemaline myopathy (NEM3). Ann. Neurol. 2018, 83, 269–282. [Google Scholar] [CrossRef]
- Clarkson, E.; Costa, C.F.; Machesky, L.M. Congenital myopathies: Diseases of the actin cytoskeleton. J. Pathol. 2004, 204, 407–417. [Google Scholar] [CrossRef]
- Gerçek, M.; Gerçek, M.; Kant, S.; Simsekyilmaz, S.; Kassner, A.; Milting, H.; Liehn, E.A.; Leube, R.E.; Krusche, C.A. Cardiomyocyte Hypertrophy in Arrhythmogenic Cardiomyopathy. Am. J. Pathol. 2017, 187, 752–766. [Google Scholar] [CrossRef] [Green Version]
- Karakikes, I.; Chaanine, A.H.; Kang, S.; Mukete, B.N.; Jeong, D.; Zhang, S.; Hajjar, R.J.; Lebeche, D. Therapeutic cardiac-targeted delivery of miR-1 reverses pressure overload-induced cardiac hypertrophy and attenuates pathological remodeling. J. Am. Heart Assoc. 2013, 2, e000078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kee, H.J.; Bae, E.H.; Park, S.; Lee, K.E.; Suh, S.H.; Kim, S.W.; Jeong, M.H. HDAC Inhibition Suppresses Cardiac Hypertrophy and Fibrosis in DOCA-Salt Hypertensive Rats via Regulation of HDAC6/HDAC8 Enzyme Activity. Kidney Blood Press. Res. 2013, 37, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.; Schneeberger, Y.; Schulz, S.; Krasemann, S.; Werner, T.; Piasecki, A.; Höppner, G.; Müller, C.; Morhenn, K.; Lorenz, K.; et al. Analysis of fibrosis in control or pressure overloaded rat hearts after mechanical unloading by heterotopic heart transplantation. Sci. Rep. 2019, 9, 5710. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Dong, S.; Li, Y. Effects of miRNA-455 on cardiac hypertrophy induced by pressure overload. Int. J. Mol. Med. 2015, 35, 893–900. [Google Scholar] [CrossRef] [Green Version]
- Kant, S.; Freytag, B.; Herzog, A.; Reich, A.; Merkel, R.; Hoffmann, B.; Krusche, C.A.; Leube, R.E. Desmoglein 2 mutation provokes skeletal muscle actin expression and accumulation at intercalated discs in murine hearts. J. Cell Sci. 2019, 132, jcs199612. [Google Scholar] [CrossRef] [Green Version]
- Olson, T.M.; Michels, V.V.; Thibodeau, S.N.; Tai, Y.S.; Keating, M.T. Actin mutations in dilated cardiomyopathy, a heritable form of heart failure. Science 1998, 280, 750–752. [Google Scholar] [CrossRef] [Green Version]
- Kluge, A.; Rangrez, A.Y.; Kilian, L.S.; Pott, J.; Bernt, A.; Frauen, R.; Rohrbeck, A.; Frey, N.; Frank, D. Rho-family GTPase 1 (Rnd1) is a biomechanical stress-sensitive activator of cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol. 2019, 129, 130–143. [Google Scholar] [CrossRef]
- Rysä, J.; Tokola, H.; Ruskoaho, H. Mechanical stretch induced transcriptomic profiles in cardiac myocytes. Sci. Rep. 2018, 8, 4733. [Google Scholar] [CrossRef]
- Bardosi, A.; Eber, S.W.; Hendrys, M.; Pekrun, A. Myopathy with altered mitochondria due to a triosephosphate isomerase (TPI) deficiency. Acta Neuropathol. 1990, 79, 387–394. [Google Scholar] [CrossRef]
- Smith, L.E.; Hambly, B.; Cordwell, S.; White, M. Abstract 20301: Reversible Oxidation Of Metabolic Protein Triosephosphate Isomerase in the Type 2 Diabetic Heart. Circulation 2016, 134, A20301. [Google Scholar]
- Ramsden, S.C.; Clayton-Smith, J.; Birch, R.; Buiting, K. Practice guidelines for the molecular analysis of Prader-Willi and Angelman syndromes. BMC Med. Genet. 2010, 11, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Brune, C.W.; Kistner, E.O.; Christian, S.L.; Courchesne, E.H.; Cox, N.J.; Cook, E.H. Transmission disequilibrium testing of the chromosome 15q11-q13 region in autism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147, 1116–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilk, J.B.; Walter, R.E.; Laramie, J.M.; Gottlieb, D.J.; O’Connor, G.T. Framingham Heart Study genome-wide association: Results for pulmonary function measures. BMC Med. Genet. 2007, 8, S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.J.A.; Ermak, G.; Rothermel, B.A.; Pritchard, M.; Heitman, J.; Ahnn, J.; Henrique-Silva, F.; Crawford, D.; Canaider, S.; Strippoli, P.; et al. Renaming the DSCR1/Adapt78 gene family as RCAN: Regulators of calcineurin. FASEB J. 2007, 21, 3023–3028. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.-Y.; Yang, J.M.; Choo, S.-M.; Kwon, K.-S.; Cho, K.-H. System-level investigation into the regulatory mechanism of the calcineurin/NFAT signaling pathway. Cell. Signal. 2008, 20, 1117–1124. [Google Scholar] [CrossRef]
- Vega, R.B.; Rothermel, B.A.; Weinheimer, C.J.; Kovacs, A.; Naseem, R.H.; Bassel-Duby, R.; Williams, R.S.; Olson, E.N. Dual roles of modulatory calcineurin-interacting protein 1 in cardiac hypertrophy. Proc. Natl. Acad. Sci. USA 2003, 100, 669–674. [Google Scholar] [CrossRef] [Green Version]
- Turpin, C.; Waikel, R. The role of myocyte-enriched calcineurin-inhibiting protein (MCIP1) in estrogen mediated repression of cardiac hypertrophy (795.5). FASEB J. 2014, 28, 795.5. [Google Scholar] [CrossRef]
- Sobrado, M.; Ramirez, B.G.; Neria, F.; Lizasoain, I.; Arbones, M.L.; Minami, T.; Redondo, J.M.; Moro, M.A.; Cano, E. Regulator of calcineurin 1 (Rcan1) has a protective role in brain ischemia/reperfusion injury. J. Neuroinflamm. 2012, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Bartoszewski, R.; Gebert, M.; Janaszak-Jasiecka, A.; Cabaj, A.; Króliczewski, J.; Bartoszewska, S.; Sobolewska, A.; Crossman, D.K.; Ochocka, R.; Kamysz, W.; et al. Genome-wide mRNA profiling identifies RCAN1 and GADD45A as regulators of the transitional switch from survival to apoptosis during ER stress. FEBS J. 2020, 287, 2923–2947. [Google Scholar] [CrossRef]
- Kuwahara, K.; Wang, Y.; McAnally, J.; Richardson, J.A.; Bassel-Duby, R.; Hill, J.A.; Olson, E.N. TRPC6 fulfills a calcineurin signaling circuit during pathologic cardiac remodeling. J. Clin. Investig. 2006, 116, 3114–3126. [Google Scholar] [CrossRef]
- Wang, X.; Feuerstein, G.Z.; Gu, J.-L.; Lysko, P.G.; Yue, T.-L. Interleukin-1β induces expression of adhesion molecules in human vascular smooth muscle cells and enhances adhesion of leukocytes to smooth muscle cells. Atherosclerosis 1995, 115, 89–98. [Google Scholar] [CrossRef]
- Ganesan, J.; Ramanujam, D.; Sassi, Y.; Ahles, A.; Jentzsch, C.; Werfel, S.; Leierseder, S.; Loyer, X.; Giacca, M.; Zentilin, L.; et al. MiR-378 controls cardiac hypertrophy by combined repression of mitogen-activated protein kinase pathway factors. Circulation 2013, 127, 2097–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Mi, X.; Chen, Y.; Feng, C.; Hou, Z.; Hui, R.; Zhang, W. MicroRNA-216a induces endothelial senescence and inflammation via Smad3/IkappaBalpha pathway. J. Cell Mol. Med. 2018, 22, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Wang, J.; Li, C.; Wang, W.; Yu, H.; Liu, J.; Kong, X.; Chen, Y. MiR-216a accelerates proliferation and fibrogenesis via targeting PTEN and SMAD7 in human cardiac fibroblasts. Cardiovasc. Diagn. Ther. 2019, 9, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Wang, Y.; Hu, L.; Xue, S.; Wang, Y.; Zhang, L.; Zhang, Y.; Qi, H.; Yu, H.; Aung, L.H.H.; et al. Combined detection of miR-21-5p, miR-30a-3p, miR-30a-5p, miR-155-5p, miR-216a and miR-217 for screening of early heart failure diseases. Biosci. Rep. 2020, 40, BSR20191653. [Google Scholar] [CrossRef] [Green Version]
- Greco, S.; Fasanaro, P.; Castelvecchio, S.; D’Alessandra, Y.; Arcelli, D.; Di Donato, M.; Malavazos, A.; Capogrossi, M.C.; Menicanti, L.; Martelli, F. MicroRNA dysregulation in diabetic ischemic heart failure patients. Diabetes 2012, 61, 1633–1641. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Bi, S.; Zhang, P. Long non-coding RNA MIAT regulates ox-LDL-induced cell proliferation, migration and invasion by miR-641/STIM1 axis in human vascular smooth muscle cells. BMC Cardiovasc. Disord. 2021, 21, 248. [Google Scholar] [CrossRef]
- Sandhu, H.; Cooper, S.; Hussain, A.; Mee, C.; Maddock, H. Attenuation of Sunitinib-induced cardiotoxicity through the A3 adenosine receptor activation. Eur. J. Pharmacol. 2017, 814, 95–105. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhang, H.Z.; Meng, F.R.; Han, S.Y.; Zhang, M. Differential expression of microRNA-411 and 376c is associated with hypertension in pregnancy. Braz. J. Med. Biol. Res. 2019, 52, e7546. [Google Scholar] [CrossRef] [Green Version]
- Santos-Faria, J.; Gavina, C.; Rodrigues, P.; Coelho, J.; da Costa Martins, P.; Leite-Moreira, A.; Falcao-Pires, I. MicroRNAs and ventricular remodeling in aortic stenosis. Rev. Port. Cardiol. 2020, 39, 377–387. [Google Scholar] [CrossRef]
- Li, F.; Sun, J.Y.; Wu, L.D.; Qu, Q.; Zhang, Z.Y.; Chen, X.F.; Kan, J.Y.; Wang, C.; Wang, R.X. Predictive Biomarkers for Postmyocardial Infarction Heart Failure Using Machine Learning: A Secondary Analysis of a Cohort Study. Evid. Based Complementary Altern. Med. 2021, 2021, 2903543. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, J.; Lin, C.; Fu, J.; Ren, J.; Li, L.; Guo, H.; Han, X.; Liu, J. Circulating MicroRNA Profiles Differ between Qi-Stagnation and Qi-Deficiency in Coronary Heart Disease Patients with Blood Stasis Syndrome. Evid Based Complementary Altern. Med. 2014, 2014, 926962. [Google Scholar] [CrossRef] [PubMed]
- Sanna, G.D.; Gabrielli, E.; De Vito, E.; Nusdeo, G.; Prisco, D.; Parodi, G. Atrial fibrillation in athletes: From epidemiology to treatment in the novel oral anticoagulants era. J. Cardiol. 2018, 72, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.G.; Meng, X.; Han, J.; Li, Y.; Luo, T.G.; Wang, J.; Xin, M.; Xi, J.Z. [Differential expressions of miRNAs in patients with nonvalvular atrial fibrillation]. Zhonghua Yi Xue Za Zhi 2012, 92, 1816–1819. [Google Scholar] [PubMed]
- Qiang, L.; Hong, L.; Ningfu, W.; Huaihong, C.; Jing, W. Expression of miR-126 and miR-508-5p in endothelial progenitor cells is associated with the prognosis of chronic heart failure patients. Int. J. Cardiol. 2013, 168, 2082–2088. [Google Scholar] [CrossRef]
- Fan, X.; Zhan, E.; Yao, Y.; Zhang, R.; Sun, Y.; Tian, X. MiR-599 Protects Cardiomyocytes against Oxidative Stress-Induced Pyroptosis. BioMed Res. Int. 2021, 2021, 3287053. [Google Scholar] [CrossRef]
- Chai, Q.; Zheng, M.; Wang, L.; Wei, M.; Yin, Y.; Ma, F.; Li, X.; Zhang, H.; Liu, G. Circ_0068655 Promotes Cardiomyocyte Apoptosis via miR-498/PAWR Axis. Tissue Eng. Regen. Med. 2020, 17, 659–670. [Google Scholar] [CrossRef]
- Zhu, R.Y.; Zhang, D.; Zou, H.D.; Zuo, X.S.; Zhou, Q.S.; Huang, H. MiR-28 inhibits cardiomyocyte survival through suppressing PDK1/Akt/mTOR signaling. In Vitro Cell Dev. Biol. Anim. 2016, 52, 1020–1025. [Google Scholar] [CrossRef]
- Li, S.P.; Liu, B.; Song, B.; Wang, C.X.; Zhou, Y.C. miR-28 promotes cardiac ischemia by targeting mitochondrial aldehyde dehydrogenase 2 (ALDH2) in mus musculus cardiac myocytes. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 752–758. [Google Scholar]
- Yang, L.; Liang, H.; Shen, L.; Guan, Z.; Meng, X. LncRNA Tug1 involves in the pulmonary vascular remodeling in mice with hypoxic pulmonary hypertension via the microRNA-374c-mediated Foxc1. Life Sci. 2019, 237, 116769. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, J.; Wang, L.; Pei, G.; Cheng, H.; Zhang, Q.; Wang, S.; He, C.; Fu, C.; Wei, Q. MiR-125 Family in Cardiovascular and Cerebrovascular Diseases. Front. Cell Dev. Biol. 2021, 9, 799049. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Lian, J.; Zhao, S.; Zheng, D.; Yang, X.; Huang, X.; Shi, X.; Sun, L.; Zhou, Q.; Shi, H.; et al. Detection of Differentially Expressed MicroRNAs in Rheumatic Heart Disease: MiR-1183 and miR-1299 as Potential Diagnostic Biomarkers. BioMed Res. Int. 2015, 2015, 524519. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Yang, J.; Lv, W.; Lu, Y.; Zhang, L.; Zhang, Y.; Musha, Z.; Fan, P.; Yang, B.; Zhou, X.; et al. Screening of differentially expressed microRNAs of essential hypertension in Uyghur population. Lipids Health Dis. 2019, 18, 98. [Google Scholar] [CrossRef] [Green Version]
- Slagsvold, K.H.; Johnsen, A.B.; Rognmo, O.; Hoydal, M.; Wisloff, U.; Wahba, A. Comparison of left versus right atrial myocardium in patients with sinus rhythm or atrial fibrillation-an assessment of mitochondrial function and microRNA expression. Physiol. Rep. 2014, 2, e12124. [Google Scholar] [CrossRef]
- Zayas-Arrabal, J.; Alquiza, A.; Rodriguez-de-Yurre, A.; Echeazarra, L.; Fernandez-Lopez, V.; Gallego, M.; Casis, O. Kv1.3 Channel Blockade Improves Inflammatory Profile, Reduces Cardiac Electrical Remodeling, and Prevents Arrhythmia in Type 2 Diabetic Rats. Cardiovasc. Drugs Ther. 2021. [Google Scholar] [CrossRef]
- Napoli, C.; Schiano, C.; Soricelli, A. Increasing evidence of pathogenic role of the Mediator (MED) complex in the development of cardiovascular diseases. Biochimie 2019, 165, 1–8. [Google Scholar] [CrossRef]
- Ward-Caviness, C.K.; Neas, L.M.; Blach, C.; Haynes, C.S.; LaRocque-Abramson, K.; Grass, E.; Dowdy, Z.E.; Devlin, R.B.; Diaz-Sanchez, D.; Cascio, W.E.; et al. A genome-wide trans-ethnic interaction study links the PIGR-FCAMR locus to coronary atherosclerosis via interactions between genetic variants and residential exposure to traffic. PLoS ONE 2017, 12, e0173880. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Yu, H.; Zeng, L.; Ma, H.; Cao, G. LncRNA Rian reduces cardiomyocyte pyroptosis and alleviates myocardial ischemia-reperfusion injury by regulating by the miR-17-5p/CCND1 axis. Hypertens. Res. 2022, 45, 976–989. [Google Scholar] [CrossRef]
- Bai, Q.; Li, Y.; Song, K.; Huang, J.; Qin, L. Knockdown of XIST up-regulates 263294miR-340-5p to relieve myocardial ischaemia-reperfusion injury via inhibiting cyclin D1. ESC Heart Fail. 2022, 9, 1050–1060. [Google Scholar] [CrossRef]
- Cubukcuoglu Deniz, G.; Durdu, S.; Dogan, Y.; Erdemli, E.; Ozdag, H.; Akar, A.R. Molecular Signatures of Human Chronic Atrial Fibrillation in Primary Mitral Regurgitation. Cardiovasc. Ther. 2021, 2021, 5516185. [Google Scholar] [CrossRef]
- Ghiroldi, A.; Piccoli, M.; Creo, P.; Cirillo, F.; Rota, P.; D’Imperio, S.; Ciconte, G.; Monasky, M.M.; Micaglio, E.; Garatti, A.; et al. Role of sialidase Neu3 and ganglioside GM3 in cardiac fibroblasts activation. Biochem. J. 2020, 477, 3401–3415. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, M.; Conforti, E.; Varrica, A.; Ghiroldi, A.; Cirillo, F.; Resmini, G.; Pluchinotta, F.; Tettamanti, G.; Giamberti, A.; Frigiola, A.; et al. NEU3 sialidase role in activating HIF-1alpha in response to chronic hypoxia in cyanotic congenital heart patients. Int. J. Cardiol. 2017, 230, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, H.; Wang, L.; Zhou, S.; Chang, M.; Zhou, J.; Dou, Y.; Wang, Y.; Shi, X. Association Between Single Nucleotide Polymorphisms in NFATC1 Signaling Pathway Genes and Susceptibility to Congenital Heart Disease in the Chinese Population. Pediatr. Cardiol. 2016, 37, 1548–1561. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, W.; Deng, K.Q.; Tian, S.; Liu, H.; Shi, H.; Fang, Q.; Liu, Z.; Chen, Z.; Tian, T.; et al. The E3 Ligase TRIM16 Is a Key Suppressor of Pathological Cardiac Hypertrophy. Circ. Res. 2022, 130, 1586–1600. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhu, J.; Glass, P.S.; Brink, P.R.; Rampil, I.J.; Rebecchi, M.J. Age-associated changes in cardiac gene expression after preconditioning. Anesthesiology 2009, 111, 1052–1064. [Google Scholar] [CrossRef] [Green Version]
- Alikunju, S.; Niranjan, N.; Mohsin, M.; Sayed, N.; Sayed, D. G3bp1-microRNA-1 axis regulates cardiomyocyte hypertrophy. Cell Signal. 2022, 91, 110245. [Google Scholar] [CrossRef]
- Li, S.J.; Hu, W.L.; Liu, D.T.; Sun, F.H.; Zhang, Q.; Yang, X.C.; Yu, S.Z.; Jin, L.; Wang, X.F. MTAP gene is associated with ischemic stroke in Chinese Hans. J. Neurol. Sci. 2009, 284, 103–107. [Google Scholar] [CrossRef]
- Wolfsberg, T.G.; Straight, P.D.; Gerena, R.L.; Huovila, A.P.; Primakoff, P.; Myles, D.G.; White, J.M. ADAM, a widely distributed and developmentally regulated gene family encoding membrane proteins with a disintegrin and metalloprotease domain. Dev. Biol. 1995, 169, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Hooft van Huijsduijnen, R. ADAM 20 and 21; two novel human testis-specific membrane metalloproteases with similarity to fertilin-alpha. Gene 1998, 206, 273–282. [Google Scholar] [CrossRef]
- Sudhir, K. Lipoprotein-associated phospholipase A 2, vascular inflammation and cardiovascular risk prediction. Vasc. Health Risk Manag. 2006, 2, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, R.; Fischer, C.; Kodelja, V.; Behrens, S.; Haas, S.; Vingron, M.; Timmermann, B.; Geikowski, A.; Sauer, S. Genome-wide analysis of LXRα activation reveals new transcriptional networks in human atherosclerotic foam cells. Nucleic Acids Res. 2013, 41, 3518–3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spadaro, O.; Youm, Y.; Shchukina, I.; Ryu, S.; Sidorov, S.; Ravussin, A.; Nguyen, K.; Aladyeva, E.; Predeus, A.N.; Smith, S.R.; et al. Caloric restriction in humans reveals immunometabolic regulators of health span. Science 2022, 375, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.L.; Zeng, Z.F.; Gan, W.Q.; Wang, W.Q.; Li, T.G.; Hou, Y.F.; Yan, Z.; Zhang, R.X.; Yang, M. Lp-PLA2 inhibition prevents Ang II-induced cardiac inflammation and fibrosis by blocking macrophage NLRP3 inflammasome activation. Acta Pharmacol. Sin. 2021, 42, 2016–2032. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhu, L.; Zhou, H.; Zheng, D.; Xu, G.; Sun, L.; Gao, J.; Shao, G. miRNA-1183-targeted regulation of Bcl-2 contributes to the pathogenesis of rheumatic heart disease. Biosci. Rep. 2020, 40, BSR20201573. [Google Scholar] [CrossRef]
- Chaanine, A.H.; Higgins, L.; Lauterboeck, L.; Markowski, T.; Yang, Q.; Delafontaine, P. Multiomics Approach Reveals an Important Role of BNIP3 in Myocardial Remodeling and the Pathogenesis of Heart Failure with Reduced Ejection Fraction. Cells 2022, 11, 1572. [Google Scholar] [CrossRef]
- Chaanine, A.H.; Joyce, L.D.; Stulak, J.M.; Maltais, S.; Joyce, D.L.; Dearani, J.A.; Klaus, K.; Nair, K.S.; Hajjar, R.J.; Redfield, M.M. Mitochondrial Morphology, Dynamics, and Function in Human Pressure Overload or Ischemic Heart Disease With Preserved or Reduced Ejection Fraction. Circ Heart Fail. 2019, 12, e005131. [Google Scholar] [CrossRef]
- Lammerich, A.; Bohm, J.; Schimke, I.; Wagner, K.D.; Storch, E.; Günther, J. Effects of hypoxia, simulated ischemia and reoxygenation on the contractile function of human atrial trabeculae. Mol. Cell. Biochem. 1996, 160, 143–151. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Parameters/Medical History | Medication (%) | ||
|---|---|---|---|
| Gender m/f (%) | 56.6/44.4 | ß-blockers | 44.4 |
| Age (y ± SD) | 66.8 ± 9.4 | ACE Inhibitors | 44.4 |
| BMI (% ± SD) | 29 ± 6.4 | AT1 Antagonist | 22.2 |
| SR (%) | 100 | MRA | 0 |
| AFIB | 0 | Statins | 77.7 |
| Paroxysmal AFIB | 0 | Ca2+ Antagonist | 44.4 |
| EF (% ± SD) | 54.9 ± 9.4 | ||
| CABG (%) | 77.8 | ||
| AVR (%) | 11.1 | ||
| MVR (%) | 11.1 | ||
| Groups | SR | AFIB |
|---|---|---|
| Demographic parameters | ||
| Gender m/f | 1/5 | 4/2 |
| Age (y ± SD) | 66 ± 10 | 73.7 ± 9.8 |
| BMI (% ± SD) | 29.2 ± 3.5 | 24.8 ± 2.9 |
| Cardiac function | ||
| EF (%± SD) | 60.8 ± 9.2 | 56.2 ± 1.5 |
| LVEDD | 49.2 ± 2.4 | 57.2 ± 3.1 |
| RVEDD | 33.8 ± 1.3 | 33.3 ± 0.9 |
| IVS | 11.8 ± 0.9 | 12.8 ± 0.4 |
| LA major axis | 62.8 ± 3.6 | 70.4 ± 2.3 |
| Comorbidities (%) | ||
| Hypertension | 83 | 100 |
| Diabetes | 17 | 0 |
| CABG | 17 | 67 |
| AVR | 17 | 17 |
| MVR | 100 | 100 |
| Medication (%) | ||
| ß-blockers | 67 | 83 |
| ACE Inhibitors | 50 | 33 |
| AT1 Antagonist | 17 | 33 |
| MRA | 17 | 17 |
| Statins | 50 | 33 |
| Ca2+ Antagonist | 17 | 0 |
| Groups | NF | DCM |
|---|---|---|
| Gender m/f (%) | 50/50 | 100/0 |
| Age (y ± SD) | 63 ± 16.3 | 59 ± 14 |
| BMI (% ± SD) | 25.4 ± 5.1 | 25.8 ± 2 |
| SR (%) | 75 | 33.3 |
| AFIB (%) | 25 | 66.7 |
| EF (%± SD) | 58 ± 0.13 | 22.5 ± 6.12 |
| CHD | 0 | 16.7 |
| Hypertension | 25 | 83.3 |
| Diabetes | 0 | 33.3 |
| Hyperlipidemia | 0 | 50 |
| Paroxysmal AFIB | 25 | 33.3 |
| CABG | 0 | 0 |
| AVR | 0 | 0 |
| MVR | 0 | 0 |
| ß-blockers | 25 | 33.3 |
| ACE Inhibitors | 25 | 50 |
| AT1 Antagonist | 0 | 0 |
| MRA | 0 | 50 |
| Statins | 0 | 16.7 |
| Ca2+ Antagonist | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djalinac, N.; Kolesnik, E.; Maechler, H.; Scheruebel-Posch, S.; Pelzmann, B.; Rainer, P.P.; Foessl, I.; Wallner, M.; Scherr, D.; Heinemann, A.; et al. miR-1183 Is a Key Marker of Remodeling upon Stretch and Tachycardia in Human Myocardium. Int. J. Mol. Sci. 2022, 23, 6962. https://doi.org/10.3390/ijms23136962
Djalinac N, Kolesnik E, Maechler H, Scheruebel-Posch S, Pelzmann B, Rainer PP, Foessl I, Wallner M, Scherr D, Heinemann A, et al. miR-1183 Is a Key Marker of Remodeling upon Stretch and Tachycardia in Human Myocardium. International Journal of Molecular Sciences. 2022; 23(13):6962. https://doi.org/10.3390/ijms23136962
Chicago/Turabian StyleDjalinac, Natasa, Ewald Kolesnik, Heinrich Maechler, Susanne Scheruebel-Posch, Brigitte Pelzmann, Peter P. Rainer, Ines Foessl, Markus Wallner, Daniel Scherr, Akos Heinemann, and et al. 2022. "miR-1183 Is a Key Marker of Remodeling upon Stretch and Tachycardia in Human Myocardium" International Journal of Molecular Sciences 23, no. 13: 6962. https://doi.org/10.3390/ijms23136962
APA StyleDjalinac, N., Kolesnik, E., Maechler, H., Scheruebel-Posch, S., Pelzmann, B., Rainer, P. P., Foessl, I., Wallner, M., Scherr, D., Heinemann, A., Sedej, S., Ljubojevic-Holzer, S., von Lewinski, D., & Bisping, E. (2022). miR-1183 Is a Key Marker of Remodeling upon Stretch and Tachycardia in Human Myocardium. International Journal of Molecular Sciences, 23(13), 6962. https://doi.org/10.3390/ijms23136962