
Molecular Recognition of Surface Trans-Sialidases in Extracellular Vesicles of the Parasite Trypanosoma cruzi Using Atomic Force Microscopy (AFM)

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
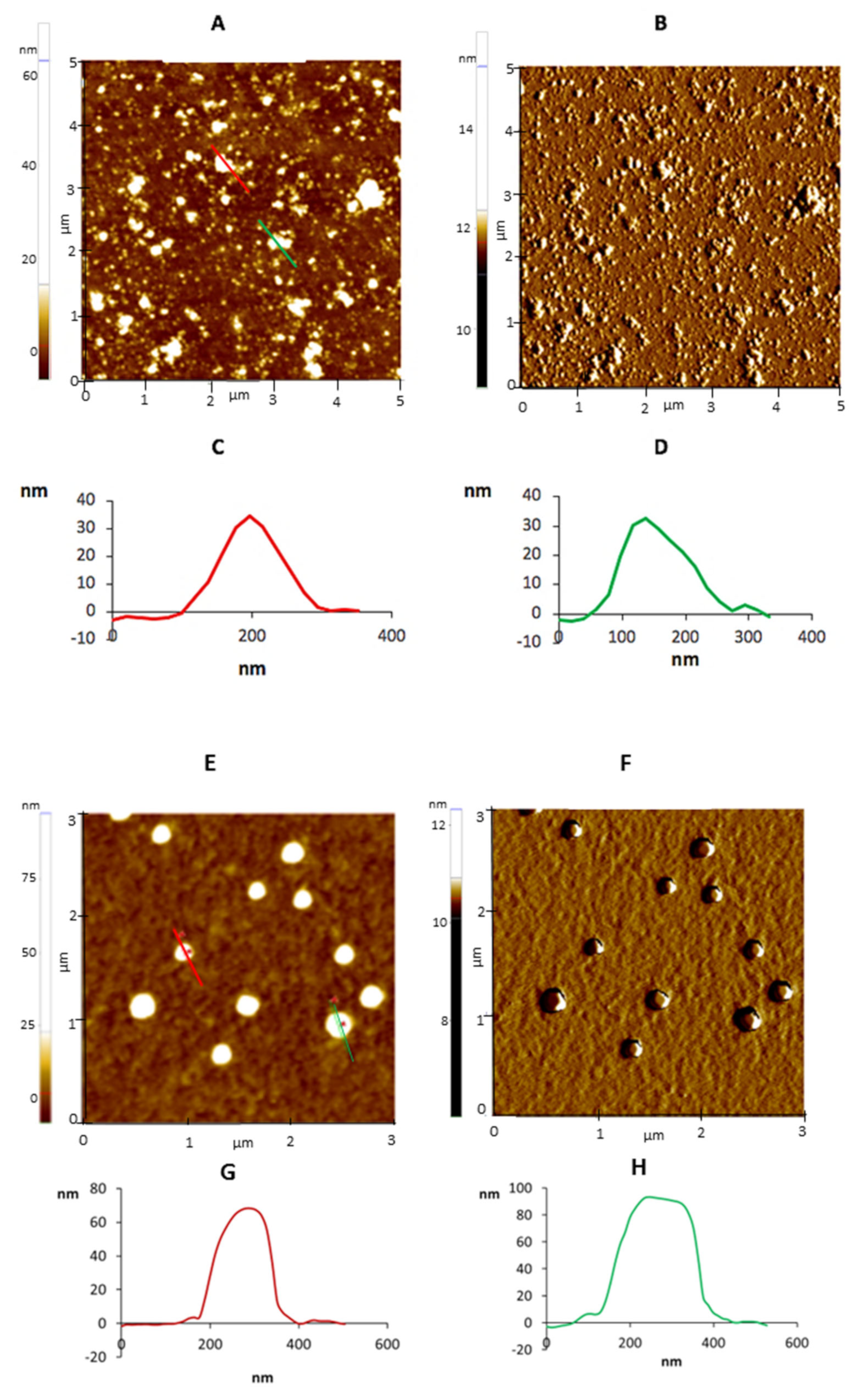
{kind=link}
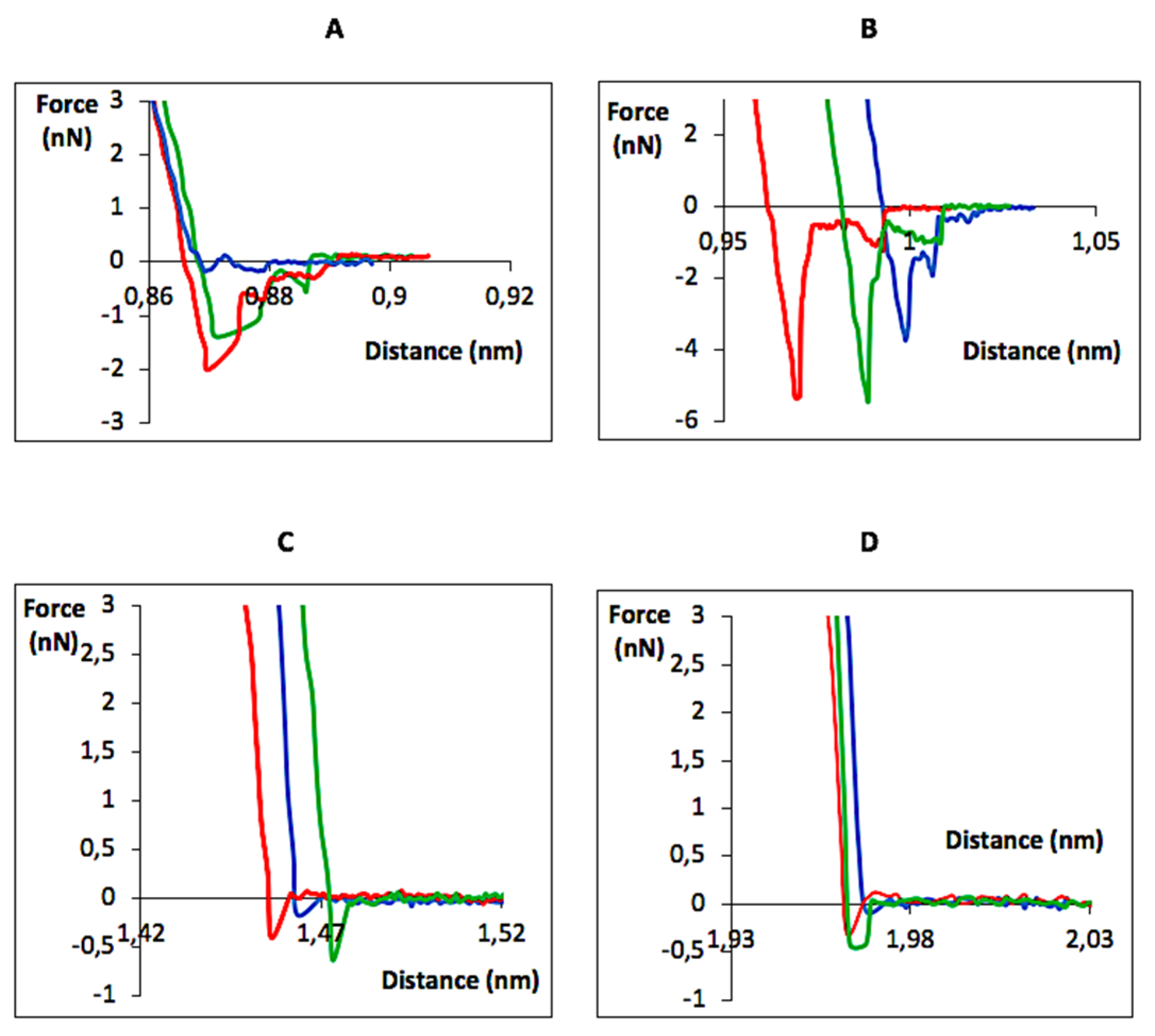
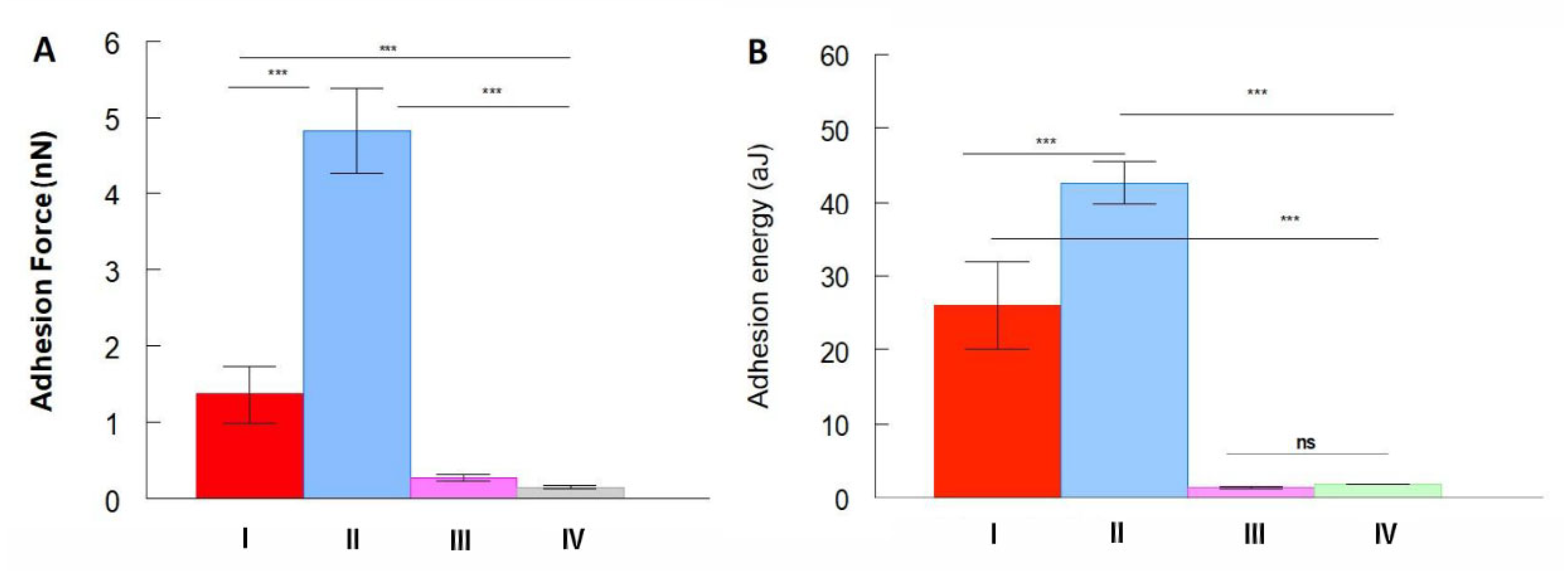{kind=link}
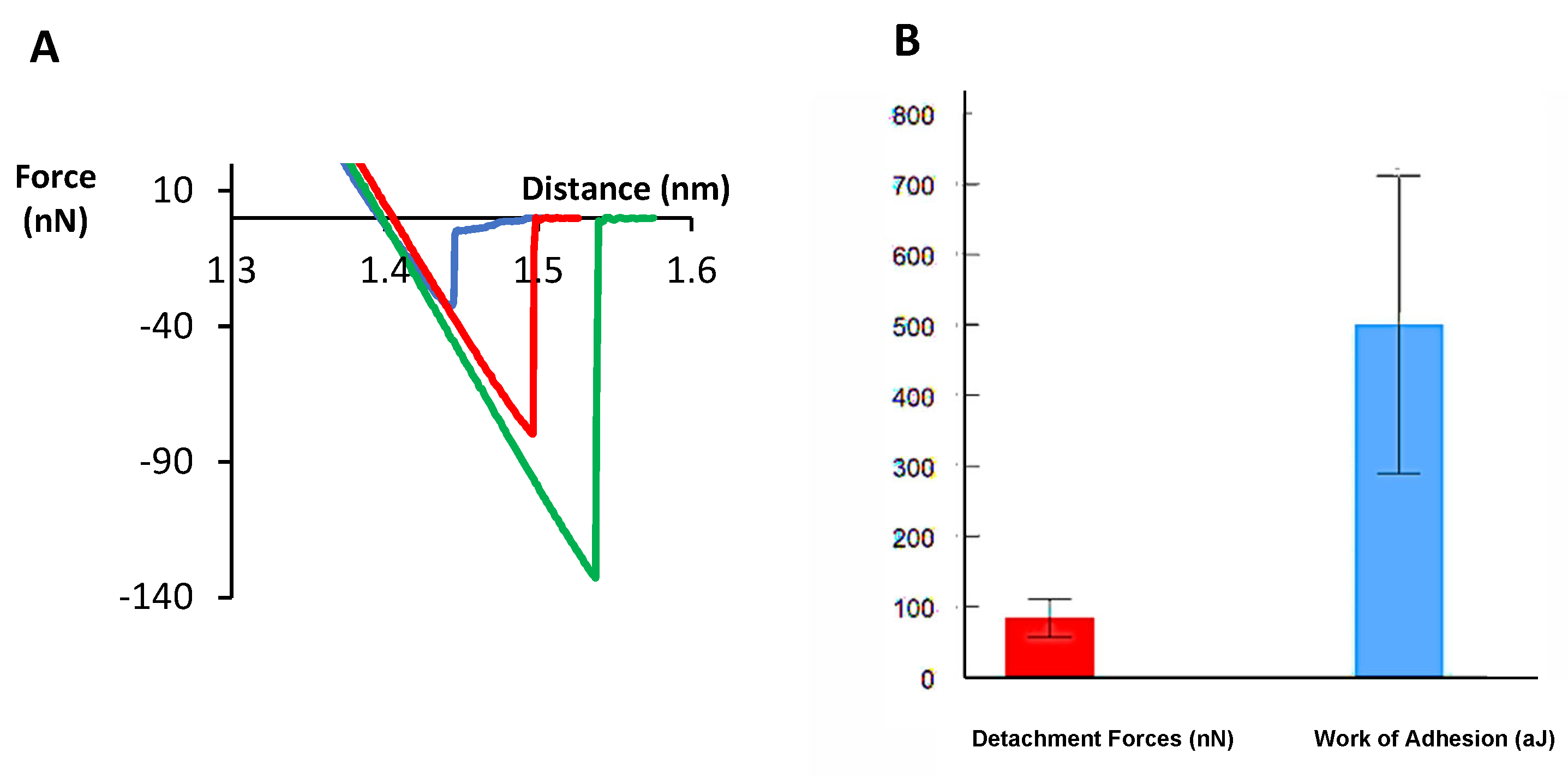{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
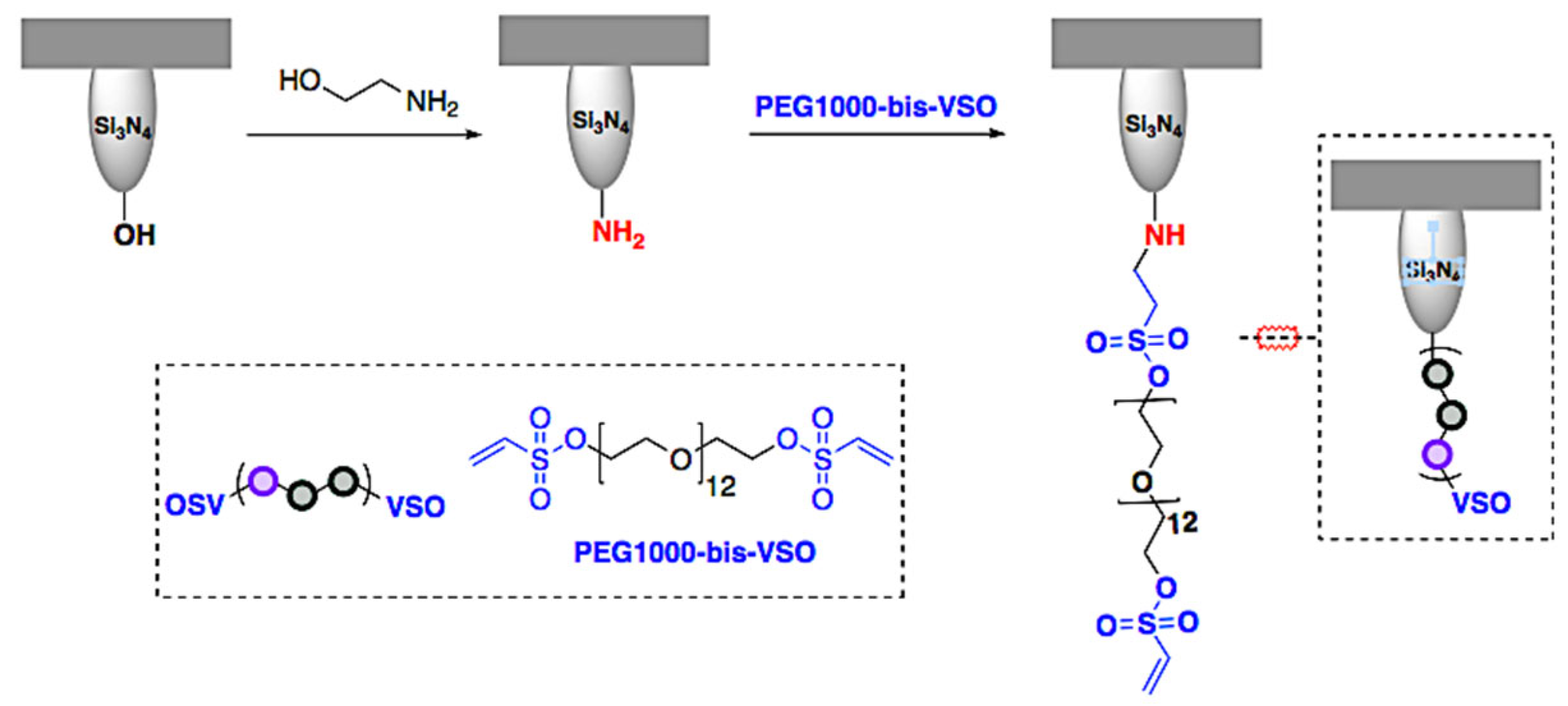
2.1. Preparation of Functionalized Cantilevers
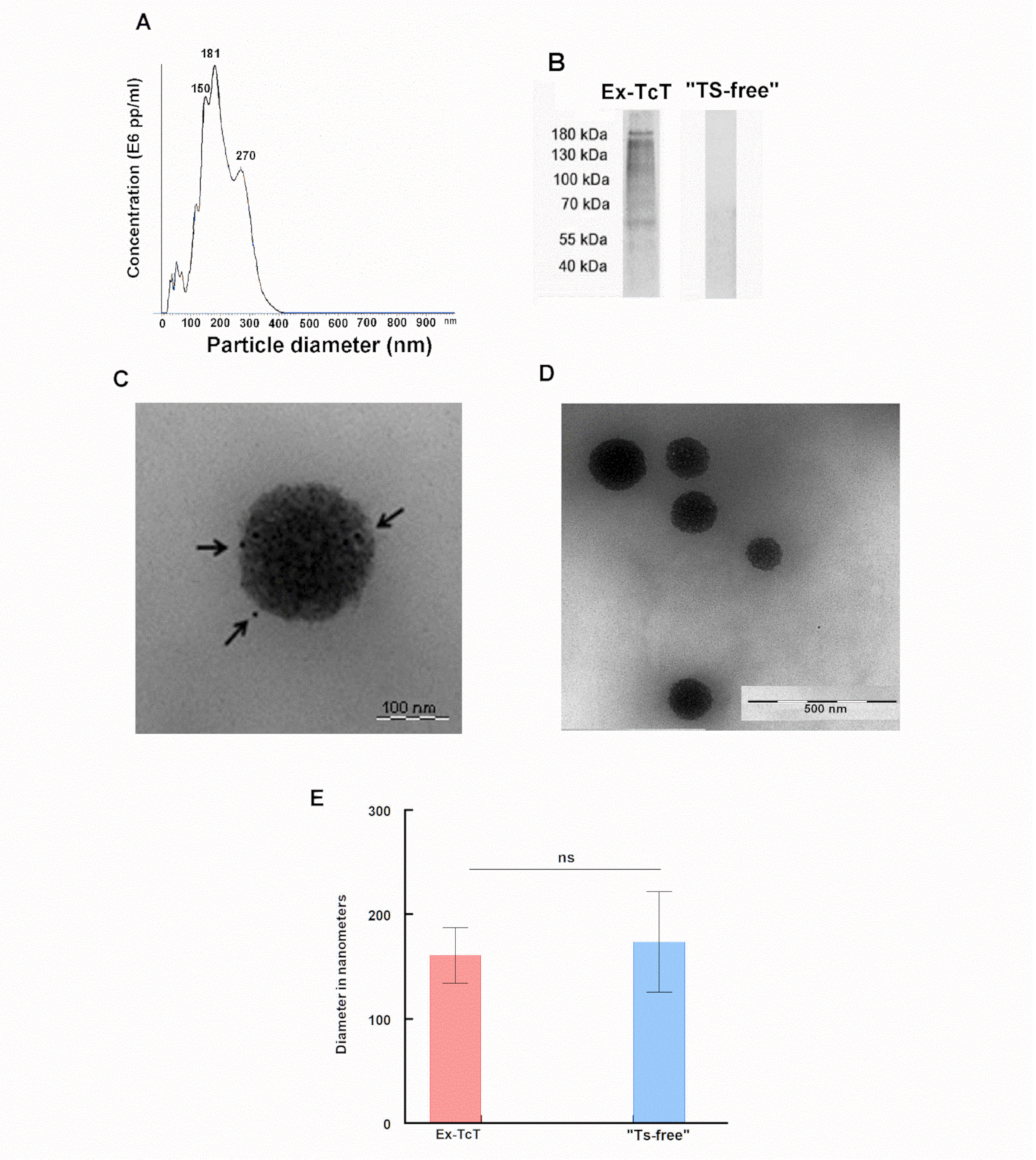
2.2. Isolation and Characterization of Ex-TcT and “TS-free” Ex-TcT
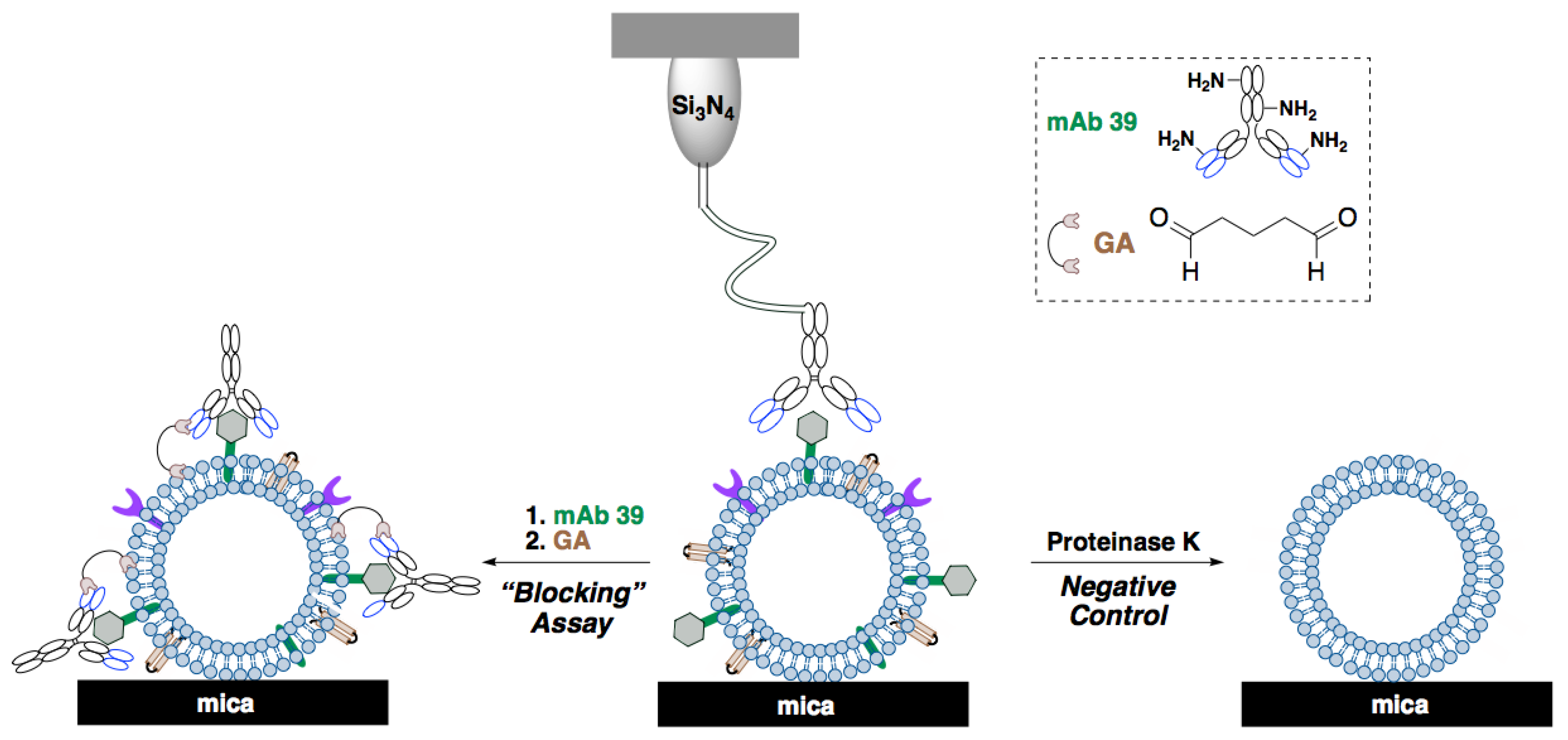
2.3. Surface Analysis of Ex-TcT, “TS-free” Ex-TcT and Ex-TcT≈mAb 39 Immune Complexes
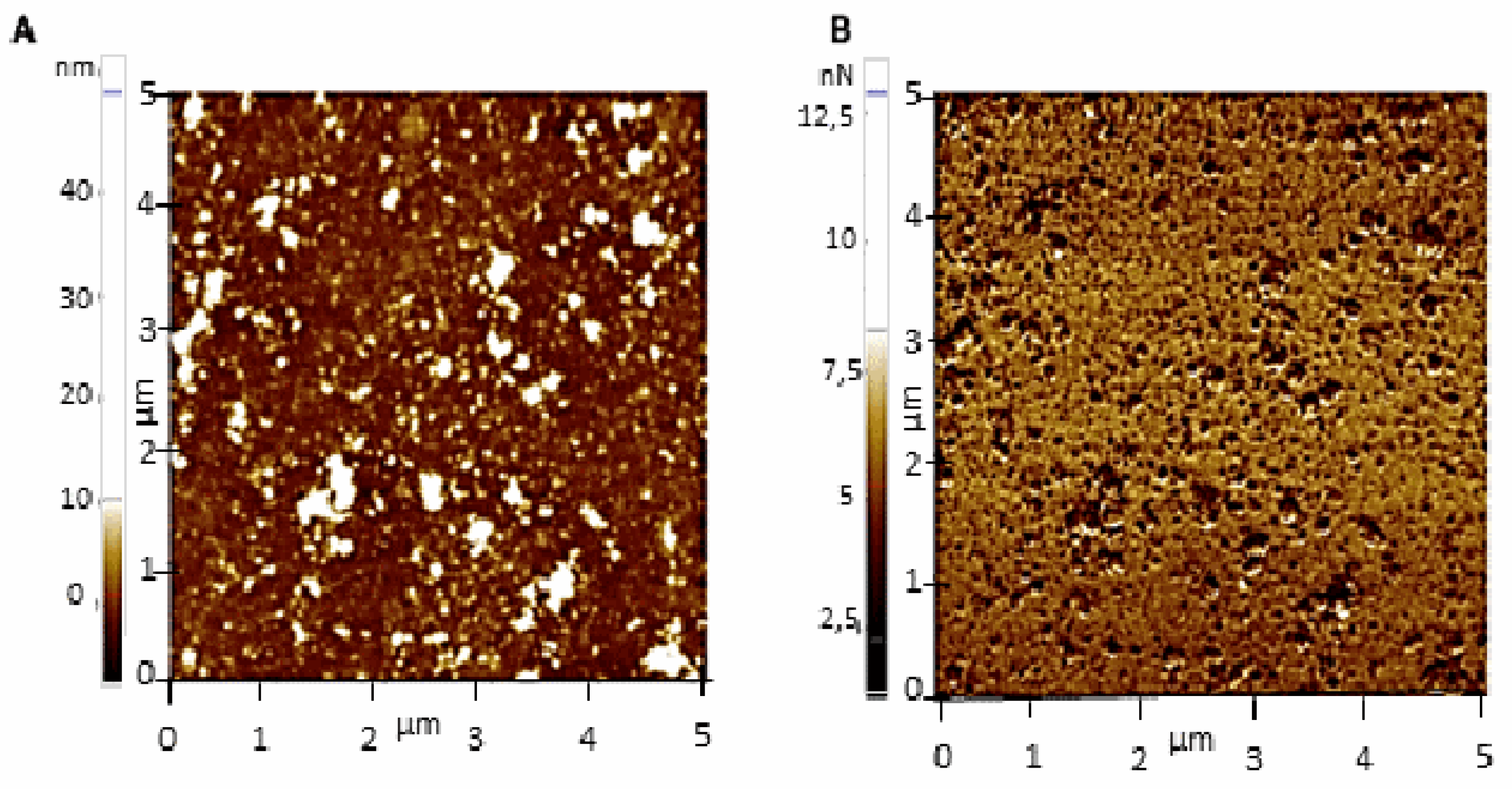
2.4. Single-Molecule Recognition Imaging of Ex-TcT of T. cruzi
3. Discussion
4. Materials and Methods
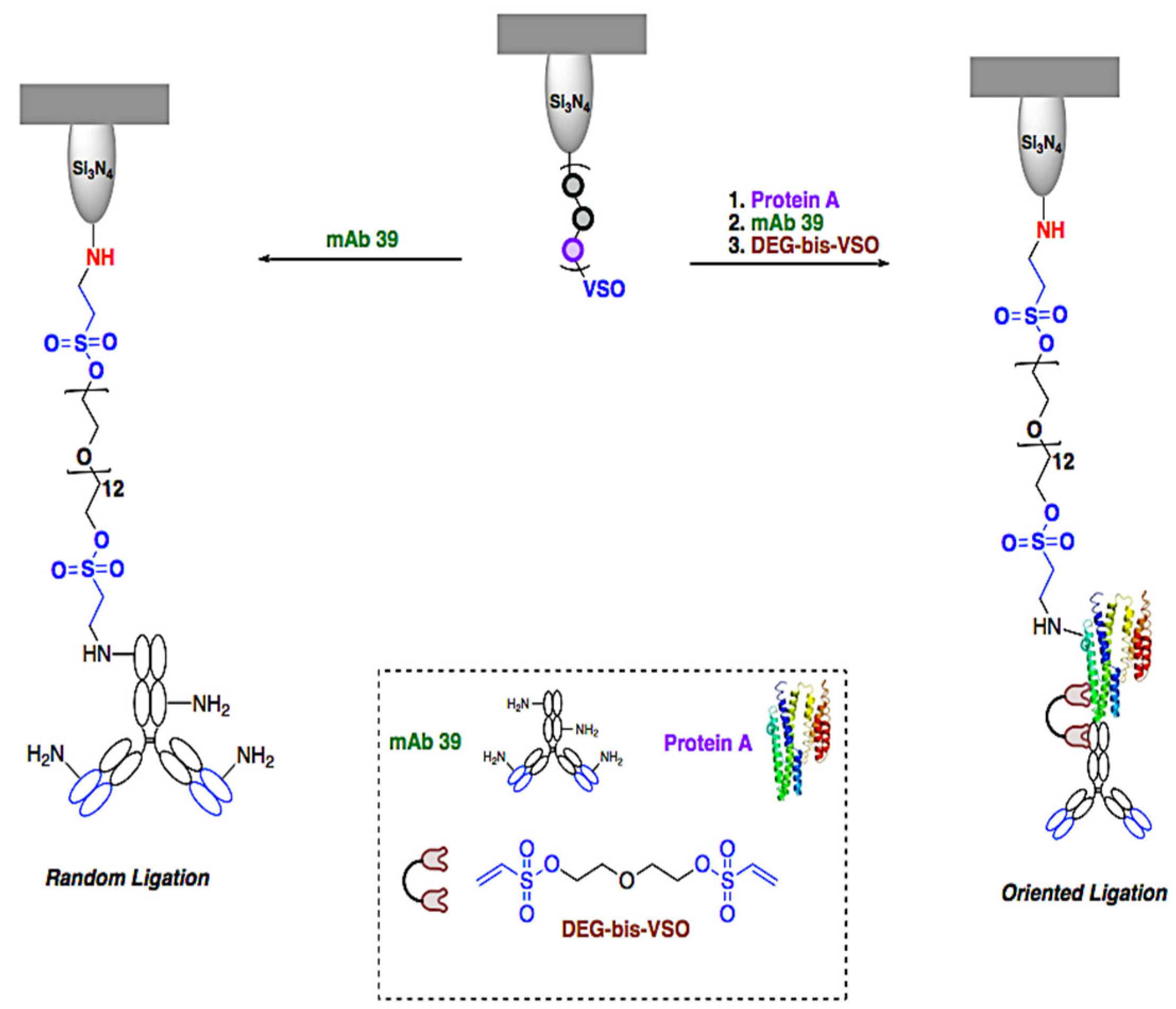
4.1. Multi-Step Conjugation of Anti-TS mAb 39 Antibodies to the Cantilever
4.1.1. Aminofunctionalization
4.1.2. Preparation of Tip-Bound PEG-VSO
4.1.3. Ligation of Anti-TS mAb 39 Antibodies
4.2. Cell Culture, Parasite Strain and Culture of Trypomastigotes of T. cruzi
4.3. Isolation of Ex-TcT of T. cruzi
4.4. Enzymatic Treatment of Ex-TcT of T. cruzi. for the Production of “TS-free” Ex-TcT
4.5. Electrophoretic Separation of Proteins from EVs from TcT of T. cruzi and Western Blot
4.6. Transmission Electron Microscopy and Immunogold Labeling
4.7. Generation of Ex-TcT≈mAb 39 Immune Complexes
4.8. Atomic Force Microscopy
4.8.1. Adsorption of Samples to Mica Sheets
4.8.2. Topographic Measurements and Imaging
4.8.3. Force Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campetella, O.; Buscaglia, C.A.; Mucci, J.; Leguizamón, M.S. Parasite-host glycan interactions during Trypanosoma cruzi infection: Trans-Sialidase rides the show. BBA-Mol. Basis Dis. 2020, 1866, 165692. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, L.M.D.; da Costa, K.M.; Chaves, V.d.S.; Freire-de-Lima, C.G.; Morrot, A.; Mendonça-Previato, L.; Previato, J.O.; Freire-de-Lima, L. Theft and Reception of Host Cell’s Sialic Acid: Dynamics of Trypanosoma Cruzi Trans-sialidases and Mucin-Like Molecules on Chagas’ Disease Immunomodulation. Front. Immunol. 2019, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lederkremer, R.M.; Giorgi, M.E.; Agusti, R. trans-Sialylation: A strategy used to incorporate sialic acid into oligosaccharides. RSC Chem. Biol. 2022, 3, 121–139. [Google Scholar] [CrossRef]
- Lantos, A.B.; Carlevaro, G.; Araoz, B.; Ruiz Diaz, P.; Camara, M.D.L.M.; Buscaglia, C.A.; Bossi, M.; Yu, H.; Chen, X.; Bertozzi, C.R.; et al. Sialic Acid Glycobiology Unveils Trypanosoma cruzi Trypomastigote Membrane Physiology. PLoS Pathog. 2016, 12, e1005559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire-De-Lima, L.; Fonseca, L.; Oeltmann, T.; Mendonça-Previato, L.; Previato, J. The trans-sialidase, the majorTrypanosoma cruzi virulence factor: Three decades of studies. Glycobiology 2015, 25, 1142–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe Costa, R.; da Silveira, J.F.; Bahia, D. Interactions between Trypanosoma cruzi Secreted Proteins and Host Cell Signaling Pathways. Front. Microbiol. 2016, 7, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossas, J.-Y.; Gulin, J.E.N.; Bisio, M.M.C.; Chapelle, M.; Marinach-Patrice, C.; Bordessoules, M.; Palazon Ruiz, G.; Vion, J.; Paris, L.; Altcheh, J.; et al. Secretome analysis of Trypanosoma cruzi by proteomics studies. PLoS ONE 2017, 12, e0185504. [Google Scholar] [CrossRef] [Green Version]
- Bayer-Santos, E.; Aguilar-Bonavides, C.; Rodrigues, S.P.; Cordero, E.M.; Marques, A.F.; Varela-Ramirez, A.; Choi, H.; Yoshida, N.; da Silveira, J.F.; Almeida, I.C. Proteomic Analysis of Trypanosoma cruzi Secretome: Characterization of Two Populations of Extracellular Vesicles and Soluble Proteins. J. Proteom. Res. 2013, 12, 883–897. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Rossi, I.V.; Ferreira Nunes, M.A.; Vargas-Otalora, S.; da Silva Ferreira, T.C.; Cortez, M.; Ramirez, M.I. Extracellular Vesicles during TriTryps infection: Complexity and future challenges. Mol. Immunol. 2021, 132, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Pereira, L.; Menna-Barreto, R.; Lannes-Vieira, J. Extracellular Vesicles: Potential Role in Remote Signaling and Inflammation in Trypanosoma cruzi-Triggered Disease. Front. Cell. Dev. Biol. 2021, 9, 3574. [Google Scholar] [CrossRef] [PubMed]
- D’Avila, H.; de Souza, N.P.; Albertoni, A.L.d.S.; Campos, L.C.; Rampinelli, P.G.; Correa, J.R.; de Almeida, P.E. Impact of the Extracellular Vesicles Derived From Trypanosoma cruzi: A Paradox in Host Response and Lipid Metabolism Modulation. Front. Cell. Dev. Biol. 2021, 11, 1039. [Google Scholar] [CrossRef] [PubMed]
- Torrecilhas, A.C.; Soares, R.P.; Schenkman, S.; Fernández-Prada, C.; Olivier, M. Extracellular Vesicles in Trypanosomatids: Host Cell Communication. Front. Cell. Dev. Biol. 2020, 10, 602502. [Google Scholar] [CrossRef]
- de Pablos Torró, L.M.; Retana Moreira, L.; Osuna, A. Extracellular Vesicles in Chagas Disease: A New Passenger for an Old Disease. Front. Microbiol. 2018, 9, 1190. [Google Scholar] [CrossRef] [Green Version]
- Retama Moreira, L.; Prescilla-Ledezma, A.; Cornet-Gomez, A.; Linares, F.; Jodar-Reyes, A.B.; Fernandez, J.; Vannucci, A.K.I.; De Pablos, L.M.; Osuna, A. Biophysical and biochemical comparison of extracellular vesicles produced by infective and non-infective stages of Trypanosoma cruzi. Int. J. Mol. Sci. 2021, 22, 5183. [Google Scholar] [CrossRef]
- de Souza, W.; Rocha, G.M. Atomic force microscopy: A tool to analyze the structural organization of pathogenic protozoa. Trends Parasitol. 2011, 27, 160–167. [Google Scholar] [CrossRef]
- Shokri, A.; Fakhar, M.; Akhtari, J.; Gill, P. Applications of nanoscopic technology in protozoal infections. J. Maz. Univ. Med. Sci. 2016, 25, 412–429. [Google Scholar]
- de Souza, W.; de Carvalho, T.U.; Barrias, E.S. 18-Ultrastructure of Trypanosoma cruzi and its interaction with host cells. In American Trypanosomiasis Chagas Disease, 2nd ed.; Telleria, J., Tibayrenc, M., Eds.; Elsevier: London, UK, 2017; pp. 401–427. [Google Scholar]
- de Souza, W.; Rocha, G.M.; Miranda, K.; Bisch, P.M.; Weissmuller, G. Atomic Force Microscopy as a Tool for the Study of the Ultrastructure of Trypanosomatid Parasites. In Atomic Force Microscopy in Biomedical Research: Methods and Protocols; Braga, P.C., Ricci, D., Eds.; Humana Press: Totowa, NJ, USA, 2011; pp. 211–221. [Google Scholar]
- Di Santo, R.; Romanò, S.; Mazzini, A.; Jovanović, S.; Nocca, G.; Campi, G.; Papi, M.; De Spirito, M.; Di Giacinto, F.; Ciasca, G. Recent advances in the label-free characterization of exosomes for cancer liquid biopsy: From scattering and spectroscopy to nanoindentation and nanodevices. Nanomaterials 2021, 11, 1476. [Google Scholar] [CrossRef]
- Malenica, M.; Vukomanović, M.; Kurtjak, M.; Masciotti, V.; Dal Zilio, S.; Greco, S.; Lazzarino, M.; Krušić, V.; Perčić, M.; Badovinac, I.J.; et al. Perspectives of microscopy methods for morphology characterisation of extracellular vesicles from human biofluids. Biomedicines 2021, 9, 603. [Google Scholar] [CrossRef]
- Sharma, S.; Leclaire, M.; Gimzewski, J.K. Ascent of atomic force microscopy as a nanoanalytical tool for exosomes and other extracellular vesicles. Nanotechnology 2018, 29, 132001. [Google Scholar] [CrossRef] [PubMed]
- Szatanek, R.; Baj-Krzyworzeka, M.; Zimoch, J.; Lekka, M.; Siedlar, M.; Baran, J. The methods of choice for extracellular vesicles (EVs) characterization. Int. J. Mol. Sci. 2017, 18, 1153. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qian, Y.; Sun, Y.; Liu, B.; Wei, G. Single-molecule force spectroscopy: A facile technique for studying the interactions between biomolecules and materials interfaces. Rev. Anal. Chem. 2020, 39, 116–129. [Google Scholar] [CrossRef]
- Li, M.; Dang, D.; Liu, L.; Xi, N.; Wang, Y. Imaging and force recognition of single molecular behaviors using atomic force microscopy. Sensors 2017, 17, 200. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, A.; Ros, R. Single-molecule force spectroscopy: A method for quantitative analysis of ligand–receptor interactions. Nanomedicine 2010, 5, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Hinterdorfer, P.; Dufrene, Y.F. Detection and localization of single molecular recognition events using atomic force microscopy. Nat. Methods 2006, 3, 347–355. [Google Scholar] [CrossRef]
- Alunda, B.O.; Lee, Y.J. Review: Cantilever-based sensors for high speed atomic force microscopy. Sensors 2020, 20, 4784. [Google Scholar] [CrossRef]
- Ebner, A.; Wildling, L.; Gruber, H.J. Functionalization of AFM Tips and Supports for Molecular Recognition Force Spectroscopy and Recognition Imaging. In Atomic Force Microscopy: Methods and Protocols; Santos, N.C., Carvalho, F.A., Eds.; Springer: New York, NY, USA, 2019; pp. 117–151. [Google Scholar]
- Ebner, A.; Wildling, L.; Zhu, R.; Rankl, C.; Haselgrübler, T.; Hinterdorfer, P.; Gruber, H.J. Functionalization of probe tips and supports for single-molecule recognition force microscopy. In Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2008; Volume 285, pp. 29–76. [Google Scholar]
- Cruz, C.M.; Ortega-Munoz, M.; Lopez-Jaramillo, F.J.; Hernandez-Mateo, F.; Blanco, V.; Santoyo-Gonzalez, F. Vinyl Sulfonates: A Click Function for Coupling-and-Decoupling Chemistry and their Applications. Adv. Synth. Catal. 2016, 358, 3394–3413. [Google Scholar] [CrossRef]
- Hinterdorfer, P.; Baumgartner, W.; Gruber, H.J.; Schilcher, K.; Schindler, H. Detection and localization of individual antibody-antigen recognition events by atomic force microscopy. Proc. Natl. Acad. Sci. USA 1996, 93, 3477–3481. [Google Scholar] [CrossRef] [Green Version]
- Ebner, A.; Hinterdorfer, P.; Gruber, H.J. Comparison of different aminofunctionalization strategies for attachment of single antibodies to AFM cantilevers. Ultramicroscopy 2007, 107, 922–927. [Google Scholar] [CrossRef]
- van der Pol, E.; Boing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, Functions, and Clinical Relevance of Extracellular Vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer-Santos, E.; Lima, F.M.; Ruiz, J.C.; Almeida, I.C.; da Silveira, J.F. Characterization of the small RNA content of Trypanosoma cruzi extracellular vesicles. Mol. Biochem. Parasit. 2014, 193, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, K.; Dutta, P.; Zareba, A.; Eldridge, M.L.; Sepaniak, M.J. Morphological and chemical optimization of microcantilever surfaces for thyroid system biosensing and beyond. Anal. Chim. Acta 2008, 625, 55–62. [Google Scholar] [CrossRef]
- Willemsen, O.H.; Snel, M.M.E.; Cambi, A.; Greve, J.; De Grooth, B.G.; Figdor, C.G. Biomolecular Interactions Measured by Atomic Force Microscopy. Biophys. J. 2000, 79, 3267–3281. [Google Scholar] [CrossRef] [Green Version]
- Riener, C.K.; Kienberger, F.; Hahn, C.D.; Buchinger, G.M.; Egwim, I.O.C.; Haselgrubler, T.; Ebner, A.; Romanin, C.; Klampfl, C.; Lackner, B.; et al. Heterobifunctional crosslinkers for tethering single ligand molecules to scanning probes. Anal. Chim. Acta 2003, 497, 101–114. [Google Scholar] [CrossRef]
- Senapati, S.; Manna, S.; Lindsay, S.; Zhang, P.M. Application of Catalyst-Free Click Reactions in Attaching Affinity Molecules to Tips of Atomic Force Microscopy for Detection of Protein Biomarkers. Langmuir 2013, 29, 14622–14630. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Jaramillo, F.J.; Hernandez-Mateo, F.; Santoyo-Gonzalez, F. Vinyl Sulfone: A Multi-Purpose Function in Proteomics. Integr. Proteom. 2012, 16, 301–326. [Google Scholar]
- Kolb, H.C.; Finn, M.G.; Sharpless, K.B. Click Chemistry: Diverse Chemical Function from a Few Good Reactions. Angew. Chem. Int. Ed. 2001, 40, 2004–2021. [Google Scholar] [CrossRef]
- McKay, C.S.; Finn, M.G. Click chemistry in complex mixtures: Bioorthogonal bioconjugation. Chem. Biol. 2014, 21, 1075–1101. [Google Scholar] [CrossRef] [Green Version]
- Nair, D.P.; Podgórski, M.; Chatani, S.; Gong, T.; Xi, W.; Fenoli, C.R.; Bowman, C.N. The Thiol-Michael addition click reaction: A powerful and widely used tool in materials chemistry. Chem. Mater. 2014, 26, 724–744. [Google Scholar] [CrossRef]
- Puntheeranurak, T.; Wildling, L.; Gruber, H.J.; Kinne, R.K.H.; Hinterdorfer, P. Ligands on the string: Single-molecule AFM studies on the interaction of antibodies and substrates with the Na+-glucose co-transporter SGLT1 in living cells. J. Cell Sci. 2006, 119, 2960–2967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puntheeranurak, T.; Wimmer, B.; Castaneda, F.; Gruber, H.J.; Hinterdorfer, P.; Kinne, R.K.H. Substrate Specificity of Sugar Transport by Rabbit SGLT1: Single-Molecule Atomic Force Microscopy versus Transport Studies. Biochemistry 2007, 46, 2797–2804. [Google Scholar] [CrossRef] [PubMed]
- Morales-Sanfrutos, J.; Lopez-Jaramillo, J.; Ortega-Munoz, M.; Megia-Fernandez, A.; Perez-Balderas, F.; Hernandez-Mateo, F.; Santoyo-Gonzalez, F. Vinyl sulfone: A versatile function for simple bioconjugation and immobilization. Org. Biomol. Chem. 2010, 8, 667–675. [Google Scholar] [CrossRef]
- Lopez-Jaramillo, F.J.; Ortega-Munoz, M.; Megia-Fernandez, A.; Hernandez-Mateo, F.; Santoyo-Gonzalez, F. Vinyl Sulfone Functionalization: A Feasible Approach for the Study of the Lectin-Carbohydrate Interactions. Bioconjugate Chem. 2012, 23, 846–855. [Google Scholar] [CrossRef]
- Kim, H.; Hoshi, M.; Iijima, M.; Kuroda, S.; Nakamura, C. Development of a universal method for the measurement of binding affinities of antibody drugs towards a living cell based on AFM force spectroscopy. Anal. Methods 2020, 12, 2922–2927. [Google Scholar] [CrossRef]
- Tan, W.; Huang, Y.; Nan, T.; Xue, C.; Li, Z.; Zhang, Q.; Wang, B. Development of protein a functionalized microcantilever immunosensors for the analyses of small molecules at parts per trillion levels. Anal. Chem. 2010, 82, 615–620. [Google Scholar] [CrossRef]
- Nugaeva, N.; Gfeller, K.Y.; Backmann, N.; Düggelin, M.; Lang, H.P.; Güntherodt, H.-J.; Hegner, M. An Antibody-Sensitized Microfabricated Cantilever for the Growth Detection of Aspergillus niger Spores. Microsc. Microanal. 2007, 13, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Stuart, J.K.; Hlady, V. Effects of Discrete Protein-Surface Interactions in Scanning Force Microscopy Adhesion Force Measurements. Langmuir 1995, 11, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Dammer, U.; Hegner, M.; Anselmetti, D.; Wagner, P.; Dreier, M.; Huber, W.; Güntherodt, H.J. Specific antigen/antibody interactions measured by force microscopy. Biophys. J. 1996, 70, 2437–2441. [Google Scholar] [CrossRef] [Green Version]
- Florin, E.L.; Radmacher, M.; Fleck, B.; Gaub, H.E. Atomic force microscope with magnetic force modulation. Rev. Sci. Instrum. 1994, 65, 639–643. [Google Scholar] [CrossRef]
- Lim, J.-S.; Kim, H.J.; Park, I.; Woo, S.; Kim, J.-H.; Park, J.W. Force Mapping Reveals the Spatial Distribution of Individual Proteins in a Neuron. Nano Lett 2022, 22, 3865–3871. [Google Scholar] [CrossRef] [PubMed]
- Ros, R.; Schwesinger, F.; Anselmetti, D.; Kubon, M.; Schafer, R.; Pluckthun, A.; Tiefenauer, L. Antigen binding forces of individually addressed single-chain Fv antibody molecules. Proc. Natl. Acad. Sci. USA 1998, 95, 7402–7405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, E.; Ritchie, K. Dynamic strength of molecular adhesion bonds. Biophys. J. 1997, 72, 1541–1555. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, J.P.; Radmacher, M.; Hansma, P.K. Atomic Scale Force Mapping with the Atomic Force Microscope. In Forces in Scanning Probe Methods; Güntherodt, H.J., Anselmetti, D., Meyer, E., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 543–549. [Google Scholar]
- Hogg, P.J.; Johnston, S.C.; Bowles, M.R.; Pond, S.M.; Winzor, D.J. Evaluation of equilibrium constants for antigen-antibody interactions by solid-phase immunoassay: The binding of paraquat to its elicited mouse monoclonal antibody. Mol. Immunol. 1987, 24, 797–801. [Google Scholar] [CrossRef]
- De Pablos, L.M.; Gonzalez, G.G.; Parada, J.S.; Hidalgo, V.S.; Lozano, I.M.D.; Samblas, M.M.G.; Bustos, T.C.; Osuna, A. Differential Expression and Characterization of a Member of the Mucin-Associated Surface Protein Family Secreted by Trypanosoma cruzi. Infect. Immun. 2011, 79, 3993–4001. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.R.; Serrano, F.R.; Osuna, A. Extracellular vesicles of Trypanosoma cruzi tissue-culture cell-derived trypomastigotes: Induction of physiological changes in non-parasitized culture cells. PLoS Neglect. Trop. Dis. 2019, 13, e0007163. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prescilla-Ledezma, A.; Linares, F.; Ortega-Muñoz, M.; Retana Moreira, L.; Jódar-Reyes, A.B.; Hernandez-Mateo, F.; Santoyo-Gonzalez, F.; Osuna, A. Molecular Recognition of Surface Trans-Sialidases in Extracellular Vesicles of the Parasite Trypanosoma cruzi Using Atomic Force Microscopy (AFM). Int. J. Mol. Sci. 2022, 23, 7193. https://doi.org/10.3390/ijms23137193
Prescilla-Ledezma A, Linares F, Ortega-Muñoz M, Retana Moreira L, Jódar-Reyes AB, Hernandez-Mateo F, Santoyo-Gonzalez F, Osuna A. Molecular Recognition of Surface Trans-Sialidases in Extracellular Vesicles of the Parasite Trypanosoma cruzi Using Atomic Force Microscopy (AFM). International Journal of Molecular Sciences. 2022; 23(13):7193. https://doi.org/10.3390/ijms23137193
Chicago/Turabian StylePrescilla-Ledezma, Alexa, Fátima Linares, Mariano Ortega-Muñoz, Lissette Retana Moreira, Ana Belén Jódar-Reyes, Fernando Hernandez-Mateo, Francisco Santoyo-Gonzalez, and Antonio Osuna. 2022. "Molecular Recognition of Surface Trans-Sialidases in Extracellular Vesicles of the Parasite Trypanosoma cruzi Using Atomic Force Microscopy (AFM)" International Journal of Molecular Sciences 23, no. 13: 7193. https://doi.org/10.3390/ijms23137193
APA StylePrescilla-Ledezma, A., Linares, F., Ortega-Muñoz, M., Retana Moreira, L., Jódar-Reyes, A. B., Hernandez-Mateo, F., Santoyo-Gonzalez, F., & Osuna, A. (2022). Molecular Recognition of Surface Trans-Sialidases in Extracellular Vesicles of the Parasite Trypanosoma cruzi Using Atomic Force Microscopy (AFM). International Journal of Molecular Sciences, 23(13), 7193. https://doi.org/10.3390/ijms23137193