
Connexin and Pannexin Large-Pore Channels in Microcirculation and Neurovascular Coupling Function

 ,
,
Abstract
:1. Introduction
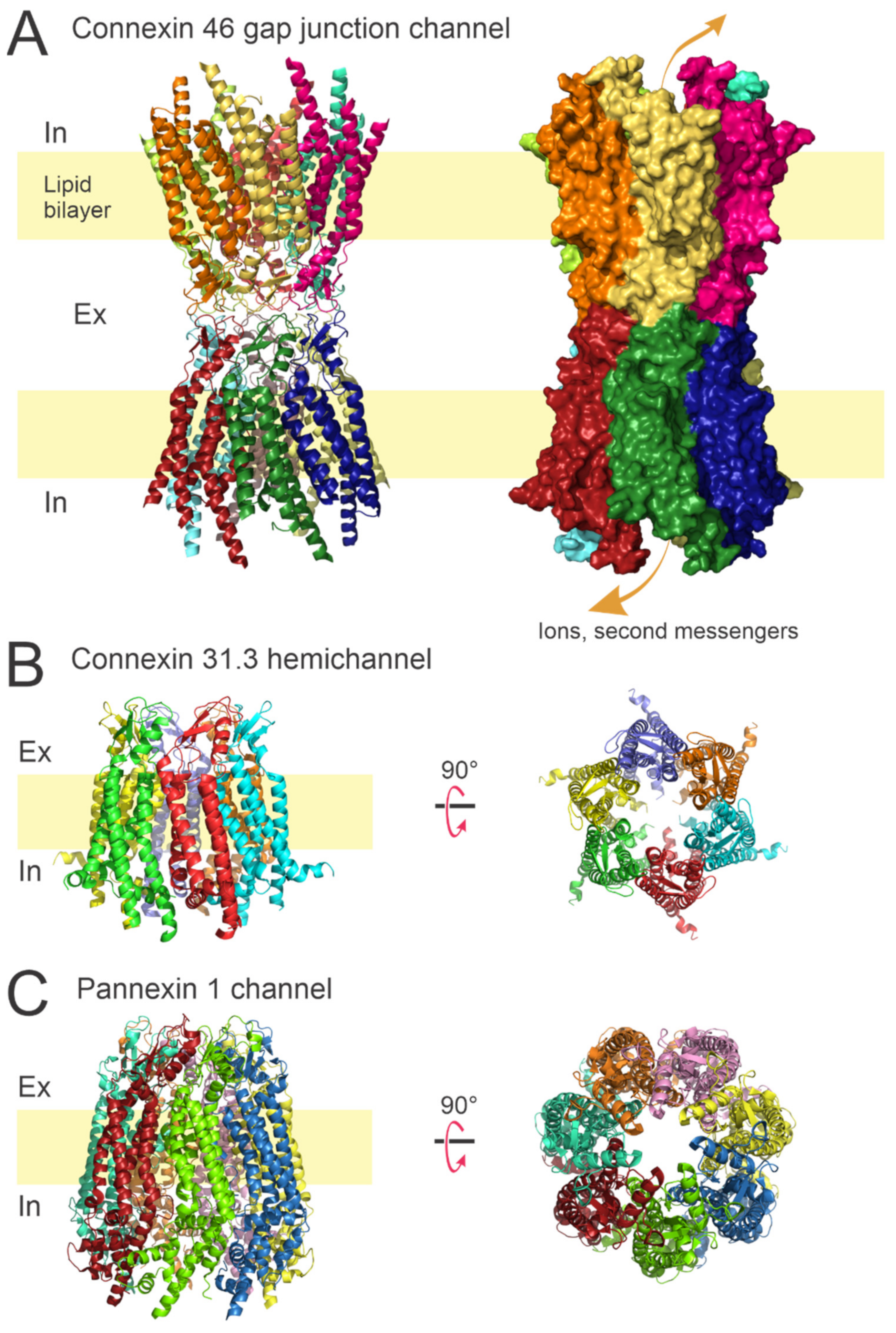
1.1. Connexin Channels
1.2. Pannexin Channels
2. Connexin Expression in Arterioles
Role of Connexin Protein Function in Arterioles
3. Pannexin Proteins in Arterioles
4. Postcapillary Venules Hyperpermeability
4.1. Connexin Proteins in Postcapillary Venules Hyperpermeability
4.2. Pannexin Proteins in Postcapillary Venules Hyperpermeability
5. Neurovascular Coupling
5.1. NVC Mediators in Astrocytes and Vascular Cells
5.2. Cxs Expression in Astrocytes and Vascular Brain Cells, and Possible Role in NVC
5.3. Panx-1 Expression in Astrocytes and Vascular Brain Cells, and Possible Role in NVC
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, W.H.; Martin, P.E. Gap junctions: Structure and function (Review). Mol. Membr. Biol. 2002, 19, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Isakson, B.E.; Ramos, S.I.; Duling, B.R. Ca2+ and inositol 1,4,5-trisphosphate-mediated signaling across the myoendothelial junction. Circ. Res. 2007, 100, 246–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saez, J.C.; Berthoud, V.M.; Branes, M.C.; Martinez, A.D.; Beyer, E.C. Plasma membrane channels formed by connexins: Their regulation and functions. Physiol. Rev. 2003, 83, 1359–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, J.E.; Saez, J.C.; Bukauskas, F.F.; Bennett, M.V. Gating and regulation of connexin 43 (Cx43) hemichannels. Proc. Natl. Acad. Sci. USA 2003, 100, 11388–11393. [Google Scholar] [CrossRef] [Green Version]
- Contreras, J.E.; Sanchez, H.A.; Eugenin, E.A.; Speidel, D.; Theis, M.; Willecke, K.; Bukauskas, F.F.; Bennett, M.V.; Saez, J.C. Metabolic inhibition induces opening of unapposed connexin 43 gap junction hemichannels and reduces gap junctional communication in cortical astrocytes in culture. Proc. Natl. Acad. Sci. USA 2002, 99, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Gaete, P.S.; Lillo, M.A.; Lopez, W.; Liu, Y.; Jiang, W.; Luo, Y.; Harris, A.L.; Contreras, J.E. A novel voltage-clamp/dye uptake assay reveals saturable transport of molecules through CALHM1 and connexin channels. J. Gen. Physiol. 2020, 152, 202–206. [Google Scholar] [CrossRef]
- Harris, A.L.; Contreras, J.E. Motifs in the permeation pathway of connexin channels mediate voltage and Ca(2+) sensing. Front. Physiol. 2014, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Lopez, W.; Ramachandran, J.; Alsamarah, A.; Luo, Y.; Harris, A.L.; Contreras, J.E. Mechanism of gating by calcium in connexin hemichannels. Proc. Natl. Acad. Sci. USA 2016, 113, E7986–E7995. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.; Lopez, W.; Ramachandran, J.; Ayad, W.A.; Liu, Y.; Lopez-Rodriguez, A.; Harris, A.L.; Contreras, J.E. Glutathione release through connexin hemichannels: Implications for chemical modification of pores permeable to large molecules. J. Gen. Physiol. 2015, 146, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Contreras, J.E.; Saez, J.C.; Bukauskas, F.F.; Bennett, M.V. Functioning of cx43 hemichannels demonstrated by single channel properties. Cell Commun. Adhes. 2003, 10, 245–249. [Google Scholar] [CrossRef]
- Saez, J.C.; Retamal, M.A.; Basilio, D.; Bukauskas, F.F.; Bennett, M.V. Connexin-based gap junction hemichannels: Gating mechanisms. Biochim. Biophys. Acta 2005, 1711, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, X.F.; Duling, B.R. Dissection of two Cx37-independent conducted vasodilator mechanisms by deletion of Cx40: Electrotonic versus regenerative conduction. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2001–H2007. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, X.F.; Duling, B.R. Gap junctions in the control of vascular function. Antioxid. Redox Signal. 2009, 11, 251–266. [Google Scholar] [CrossRef] [Green Version]
- Gaete, P.S.; Lillo, M.A.; Figueroa, X.F. Functional role of connexins and pannexins in the interaction between vascular and nervous system. J. Cell. Physiol. 2014, 229, 1336–1345. [Google Scholar] [CrossRef]
- Lillo, M.A.; Himelman, E.; Shirokova, N.; Xie, L.H.; Fraidenraich, D.; Contreras, J.E. S-nitrosylation of connexin43 hemichannels elicits cardiac stress-induced arrhythmias in Duchenne muscular dystrophy mice. JCI Insight 2019, 4, 130–136. [Google Scholar] [CrossRef]
- Vargas, A.A.; Cisterna, B.A.; Saavedra-Leiva, F.; Urrutia, C.; Cea, L.A.; Vielma, A.H.; Gutierrez-Maldonado, S.E.; Martin, A.J.; Pareja-Barrueto, C.; Escalona, Y.; et al. On Biophysical Properties and Sensitivity to Gap Junction Blockers of Connexin 39 Hemichannels Expressed in HeLa Cells. Front. Physiol. 2017, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Nieto, D.; Gomez-Hernandez, J.M.; Larrosa, B.; Gutierrez, C.; Munoz, M.D.; Fasciani, I.; O’Brien, J.; Zappala, A.; Cicirata, F.; Barrio, L.C. Regulation of neuronal connexin-36 channels by pH. Proc. Natl. Acad. Sci. USA 2008, 105, 17169–17174. [Google Scholar] [CrossRef] [Green Version]
- Skeberdis, V.A.; Rimkute, L.; Skeberdyte, A.; Paulauskas, N.; Bukauskas, F.F. pH-dependent modulation of connexin-based gap junctional uncouplers. J. Physiol. 2011, 589 Pt 14, 3495–3506. [Google Scholar] [CrossRef]
- Yamaguchi, D.T.; Ma, D. Mechanism of pH regulation of connexin 43 expression in MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 2003, 304, 736–739. [Google Scholar] [CrossRef]
- Retamal, M.A.; Schalper, K.A.; Shoji, K.F.; Bennett, M.V.; Saez, J.C. Opening of connexin 43 hemichannels is increased by lowering intracellular redox potential. Proc. Natl. Acad. Sci. USA 2007, 104, 8322–8327. [Google Scholar] [CrossRef] [Green Version]
- Bennett, B.C.; Purdy, M.D.; Baker, K.A.; Acharya, C.; McIntire, W.E.; Stevens, R.C.; Zhang, Q.; Harris, A.L.; Abagyan, R.; Yeager, M. An electrostatic mechanism for Ca(2+)-mediated regulation of gap junction channels. Nat. Commun. 2016, 7, 8770. [Google Scholar] [CrossRef] [Green Version]
- Leon-Paravic, C.G.; Figueroa, V.A.; Guzman, D.J.; Valderrama, C.F.; Vallejos, A.A.; Fiori, M.C.; Altenberg, G.A.; Reuss, L.; Retamal, M.A. Carbon monoxide (CO) is a novel inhibitor of connexin hemichannels. J. Biol. Chem. 2014, 289, 36150–36157. [Google Scholar] [CrossRef] [Green Version]
- Hirschhauser, C.; Lissoni, A.; Gorge, P.M.; Lampe, P.D.; Heger, J.; Schluter, K.D.; Leybaert, L.; Schulz, R.; Boengler, K. Connexin 43 phosphorylation by casein kinase 1 is essential for the cardioprotection by ischemic preconditioning. Basic Res. Cardiol. 2021, 116, 21. [Google Scholar] [CrossRef]
- Johnstone, S.R.; Kroncke, B.M.; Straub, A.C.; Best, A.K.; Dunn, C.A.; Mitchell, L.A.; Peskova, Y.; Nakamoto, R.K.; Koval, M.; Lo, C.W.; et al. MAPK phosphorylation of connexin 43 promotes binding of cyclin E and smooth muscle cell proliferation. Circ. Res. 2012, 111, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Slavi, N.; Toychiev, A.H.; Kosmidis, S.; Ackert, J.; Bloomfield, S.A.; Wulff, H.; Viswanathan, S.; Lampe, P.D.; Srinivas, M. Suppression of connexin 43 phosphorylation promotes astrocyte survival and vascular regeneration in proliferative retinopathy. Proc. Natl. Acad. Sci. USA 2018, 115, E5934–E5943. [Google Scholar] [CrossRef] [Green Version]
- Retamal, M.A.; Cortes, C.J.; Reuss, L.; Bennett, M.V.; Saez, J.C. S-nitrosylation and permeation through connexin 43 hemichannels in astrocytes: Induction by oxidant stress and reversal by reducing agents. Proc. Natl. Acad. Sci. USA 2006, 103, 4475–4480. [Google Scholar] [CrossRef] [Green Version]
- Vielma, A.Z.; Boric, M.P.; Gonzalez, D.R. Apocynin Treatment Prevents Cardiac Connexin 43 Hemichannels Hyperactivity by Reducing Nitroso-Redox Stress in Mdx Mice. Int. J. Mol. Sci. 2020, 21, 5415. [Google Scholar] [CrossRef]
- Delvaeye, T.; De Smet, M.A.J.; Verwaerde, S.; Decrock, E.; Czekaj, A.; Vandenbroucke, R.E.; Lemeire, K.; Goncalves, A.; Declercq, W.; Vandenabeele, P.; et al. Blocking connexin43 hemichannels protects mice against tumour necrosis factor-induced inflammatory shock. Sci. Rep. 2019, 9, 16623. [Google Scholar] [CrossRef] [Green Version]
- De Smet, M.A.; Lissoni, A.; Nezlobinsky, T.; Wang, N.; Dries, E.; Perez-Hernandez, M.; Lin, X.; Amoni, M.; Vervliet, T.; Witschas, K.; et al. Cx43 hemichannel microdomain signaling at the intercalated disc enhances cardiac excitability. J. Clin. Investig. 2021, 131, 173–179. [Google Scholar] [CrossRef]
- Kim, J.C.; Perez-Hernandez, M.; Alvarado, F.J.; Maurya, S.R.; Montnach, J.; Yin, Y.; Zhang, M.; Lin, X.; Vasquez, C.; Heguy, A.; et al. Disruption of Ca(2+)i Homeostasis and Connexin 43 Hemichannel Function in the Right Ventricle Precedes Overt Arrhythmogenic Cardiomyopathy in Plakophilin-2-Deficient Mice. Circulation 2019, 140, 1015–1030. [Google Scholar] [CrossRef]
- Yang, D.; Shen, J.; Fan, J.; Chen, Y.; Guo, X. Paracellular permeability changes induced by multi-walled carbon nanotubes in brain endothelial cells and associated roles of hemichannels. Toxicology 2020, 440, 152491. [Google Scholar] [CrossRef] [PubMed]
- Meunier, C.; Wang, N.; Yi, C.; Dallerac, G.; Ezan, P.; Koulakoff, A.; Leybaert, L.; Giaume, C. Contribution of Astroglial Cx43 Hemichannels to the Modulation of Glutamatergic Currents by D-Serine in the Mouse Prefrontal Cortex. J. Neurosci. 2017, 37, 9064–9075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosch, M.; Zindel, J.; Jebbawi, F.; Melin, N.; Sanchez-Taltavull, D.; Stroka, D.; Candinas, D.; Beldi, G. Connexin-43-dependent ATP release mediates macrophage activation during sepsis. Elife 2019, 8, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Anselmi, F.; Hernandez, V.H.; Crispino, G.; Seydel, A.; Ortolano, S.; Roper, S.D.; Kessaris, N.; Richardson, W.; Rickheit, G.; Filippov, M.A.; et al. ATP release through connexin hemichannels and gap junction transfer of second messengers propagate Ca2+ signals across the inner ear. Proc. Natl. Acad. Sci. USA 2008, 105, 18770–18775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Riquelme, M.A.; Guda, T.; Tu, C.; Xu, H.; Gu, S.; Jiang, J.X. Connexin hemichannels with prostaglandin release in anabolic function of bone to mechanical loading. Elife 2022, 11, e74365. [Google Scholar] [CrossRef]
- Flores, J.A.; Haddad, B.G.; Dolan, K.A.; Myers, J.B.; Yoshioka, C.C.; Copperman, J.; Zuckerman, D.M.; Reichow, S.L. Connexin-46/50 in a dynamic lipid environment resolved by CryoEM at 1.9 A. Nat. Commun. 2020, 11, 4331. [Google Scholar] [CrossRef]
- Lee, H.J.; Jeong, H.; Hyun, J.; Ryu, B.; Park, K.; Lim, H.H.; Yoo, J.; Woo, J.S. Cryo-EM structure of human Cx31.3/GJC3 connexin hemichannel. Sci. Adv. 2020, 6, eaba4996. [Google Scholar] [CrossRef]
- Ruan, Z.; Orozco, I.J.; Du, J.; Lu, W. Structures of human pannexin 1 reveal ion pathways and mechanism of gating. Nature 2020, 584, 646–651. [Google Scholar] [CrossRef]
- Garcia, I.E.; Maripillan, J.; Jara, O.; Ceriani, R.; Palacios-Munoz, A.; Ramachandran, J.; Olivero, P.; Perez-Acle, T.; Gonzalez, C.; Saez, J.C.; et al. Keratitis-ichthyosis-deafness syndrome-associated Cx26 mutants produce nonfunctional gap junctions but hyperactive hemichannels when co-expressed with wild type Cx43. J. Investig. Dermatol 2015, 135, 1338–1347. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, H.A.; Verselis, V.K. Aberrant Cx26 hemichannels and keratitis-ichthyosis-deafness syndrome: Insights into syndromic hearing loss. Front. Cell. Neurosci. 2014, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, H.A.; Villone, K.; Srinivas, M.; Verselis, V.K. The D50N mutation and syndromic deafness: Altered Cx26 hemichannel properties caused by effects on the pore and intersubunit interactions. J. Gen. Physiol. 2013, 142, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Nicholson, B.J. The role of connexins in ear and skin physiology—Functional insights from disease-associated mutations. Biochim. Biophys. Acta 2013, 1828, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.R.; Derosa, A.M.; White, T.W. Connexin mutations causing skin disease and deafness increase hemichannel activity and cell death when expressed in Xenopus oocytes. J. Investig. Dermatol. 2009, 129, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.R.; White, T.W. Connexin-26 mutations in deafness and skin disease. Expert Rev. Mol. Med. 2009, 11, e35. [Google Scholar] [CrossRef]
- Beyer, E.C.; Ebihara, L.; Berthoud, V.M. Connexin mutants and cataracts. Front. Pharmacol. 2013, 4, 43. [Google Scholar] [CrossRef] [Green Version]
- Vinken, M.; Decrock, E.; De Vuyst, E.; De Bock, M.; Vandenbroucke, R.E.; De Geest, B.G.; Demeester, J.; Sanders, N.N.; Vanhaecke, T.; Leybaert, L.; et al. Connexin32 hemichannels contribute to the apoptotic-to-necrotic transition during Fas-mediated hepatocyte cell death. Cell. Mol. Life Sci. 2010, 67, 907–918. [Google Scholar] [CrossRef]
- Cea, L.A.; Cisterna, B.A.; Puebla, C.; Frank, M.; Figueroa, X.F.; Cardozo, C.; Willecke, K.; Latorre, R.; Saez, J.C. De novo expression of connexin hemichannels in denervated fast skeletal muscles leads to atrophy. Proc. Natl. Acad. Sci. USA 2013, 110, 16229–16234. [Google Scholar] [CrossRef] [Green Version]
- Contreras, J.E.; Sanchez, H.A.; Veliz, L.P.; Bukauskas, F.F.; Bennett, M.V.; Saez, J.C. Role of connexin-based gap junction channels and hemichannels in ischemia-induced cell death in nervous tissue. Brain Res. Rev. 2004, 47, 290–303. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Niu, J.; Yu, G.; Ezan, P.; Yi, C.; Wang, X.; Koulakoff, A.; Gao, X.; Chen, X.; Saez, J.C.; et al. Connexin 43 deletion in astrocytes promotes CNS remyelination by modulating local inflammation. Glia 2020, 68, 1201–1212. [Google Scholar] [CrossRef]
- Himelman, E.; Lillo, M.A.; Nouet, J.; Gonzalez, J.P.; Zhao, Q.; Xie, L.H.; Li, H.; Liu, T.; Wehrens, X.H.; Lampe, P.D.; et al. Prevention of connexin-43 remodeling protects against Duchenne muscular dystrophy cardiomyopathy. J. Clin. Investig. 2020, 130, 1713–1727. [Google Scholar] [CrossRef]
- Fernandez, G.; Arias-Bravo, G.; Bevilacqua, J.A.; Castillo-Ruiz, M.; Caviedes, P.; Saez, J.C.; Cea, L.A. Myofibers deficient in connexins 43 and 45 expression protect mice from skeletal muscle and systemic dysfunction promoted by a dysferlin mutation. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165800. [Google Scholar] [CrossRef]
- Cisterna, B.A.; Vargas, A.A.; Puebla, C.; Fernandez, P.; Escamilla, R.; Lagos, C.F.; Matus, M.F.; Vilos, C.; Cea, L.A.; Barnafi, E.; et al. Active acetylcholine receptors prevent the atrophy of skeletal muscles and favor reinnervation. Nat. Commun. 2020, 11, 1073. [Google Scholar] [CrossRef] [Green Version]
- Almad, A.A.; Taga, A.; Joseph, J.; Gross, S.K.; Welsh, C.; Patankar, A.; Richard, J.P.; Rust, K.; Pokharel, A.; Plott, C.; et al. Cx43 hemichannels contribute to astrocyte-mediated toxicity in sporadic and familial ALS. Proc. Natl. Acad. Sci. USA 2022, 119, 154–169. [Google Scholar] [CrossRef]
- Qu, R.; Dong, L.; Zhang, J.; Yu, X.; Wang, L.; Zhu, S. Cryo-EM structure of human heptameric Pannexin 1 channel. Cell Res. 2020, 30, 446–448. [Google Scholar] [CrossRef]
- Michalski, K.; Syrjanen, J.L.; Henze, E.; Kumpf, J.; Furukawa, H.; Kawate, T. The Cryo-EM structure of pannexin 1 reveals unique motifs for ion selection and inhibition. Elife 2020, 9, e54670. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; He, Z.; Maksaev, G.; Bitter, R.M.; Rau, M.; Fitzpatrick, J.A.J.; Yuan, P. Cryo-EM structures of the ATP release channel pannexin 1. Nat. Struct. Mol. Biol. 2020, 27, 373–381. [Google Scholar] [CrossRef]
- Unger, V.M.; Kumar, N.M.; Gilula, N.B.; Yeager, M. Three-dimensional structure of a recombinant gap junction membrane channel. Science 1999, 283, 1176–1180. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.B.; Haddad, B.G.; O’Neill, S.E.; Chorev, D.S.; Yoshioka, C.C.; Robinson, C.V.; Zuckerman, D.M.; Reichow, S.L. Structure of native lens connexin 46/50 intercellular channels by cryo-EM. Nature 2018, 564, 372–377. [Google Scholar] [CrossRef]
- Maeda, S.; Nakagawa, S.; Suga, M.; Yamashita, E.; Oshima, A.; Fujiyoshi, Y.; Tsukihara, T. Structure of the connexin 26 gap junction channel at 3.5 A resolution. Nature 2009, 458, 597–602. [Google Scholar] [CrossRef]
- Penuela, S.; Gehi, R.; Laird, D.W. The biochemistry and function of pannexin channels. Biochim. Biophys. Acta 2013, 1828, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Penuela, S.; Bhalla, R.; Gong, X.Q.; Cowan, K.N.; Celetti, S.J.; Cowan, B.J.; Bai, D.; Shao, Q.; Laird, D.W. Pannexin 1 and pannexin 3 are glycoproteins that exhibit many distinct characteristics from the connexin family of gap junction proteins. J. Cell Sci. 2007, 120 Pt 21, 3772–3783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios-Prado, N.; Soto, P.A.; Lopez, X.; Choi, E.J.; Marquez-Miranda, V.; Rojas, M.; Duarte, Y.; Lee, J.; Gonzalez-Nilo, F.D.; Saez, J.C. Endogenous pannexin1 channels form functional intercellular cell-cell channels with characteristic voltage-dependent properties. Proc. Natl. Acad. Sci. USA 2022, 119, 315–365. [Google Scholar] [CrossRef] [PubMed]
- Beckel, J.M.; Daugherty, S.L.; Tyagi, P.; Wolf-Johnston, A.S.; Birder, L.A.; Mitchell, C.H.; de Groat, W.C. Pannexin 1 channels mediate the release of ATP into the lumen of the rat urinary bladder. J. Physiol. 2015, 593, 1857–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narahari, A.K.; Kreutzberger, A.J.; Gaete, P.S.; Chiu, Y.H.; Leonhardt, S.A.; Medina, C.B.; Jin, X.; Oleniacz, P.W.; Kiessling, V.; Barrett, P.Q.; et al. ATP and large signaling metabolites flux through caspase-activated Pannexin 1 channels. Elife 2021, 10, e64787. [Google Scholar] [CrossRef] [PubMed]
- Lohman, A.W.; Leskov, I.L.; Butcher, J.T.; Johnstone, S.R.; Stokes, T.A.; Begandt, D.; DeLalio, L.J.; Best, A.K.; Penuela, S.; Leitinger, N.; et al. Pannexin 1 channels regulate leukocyte emigration through the venous endothelium during acute inflammation. Nat. Commun. 2015, 6, 7965. [Google Scholar] [CrossRef]
- Van Kempen, M.J.; Jongsma, H.J. Distribution of connexin37, connexin40 and connexin43 in the aorta and coronary artery of several mammals. Histochem. Cell Biol. 1999, 112, 479–486. [Google Scholar] [CrossRef]
- Gustafsson, F.; Mikkelsen, H.B.; Arensbak, B.; Thuneberg, L.; Neve, S.; Jensen, L.J.; Holstein-Rathlou, N.H. Expression of connexin 37, 40 and 43 in rat mesenteric arterioles and resistance arteries. Histochem. Cell Biol. 2003, 119, 139–148. [Google Scholar] [CrossRef]
- Figueroa, X.F.; Paul, D.L.; Simon, A.M.; Goodenough, D.A.; Day, K.H.; Damon, D.N.; Duling, B.R. Central role of connexin40 in the propagation of electrically activated vasodilation in mouse cremasteric arterioles in vivo. Circ. Res. 2003, 92, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Gaete, P.S.; Lillo, M.A.; Puebla, M.; Poblete, I.; Figueroa, X.F. CGRP signalling inhibits NO production through pannexin-1 channel activation in endothelial cells. Sci. Rep. 2019, 9, 7932. [Google Scholar] [CrossRef] [Green Version]
- Lillo, M.A.; Gaete, P.S.; Puebla, M.; Burboa, P.C.; Poblete, I.; Figueroa, X.F. Novel Pannexin-1-Coupled Signaling Cascade Involved in the Control of Endothelial Cell Function and NO-Dependent Relaxation. Oxid. Med. Cell. Longev. 2021, 2021, 2678134. [Google Scholar] [CrossRef]
- Lohman, A.W.; Billaud, M.; Straub, A.C.; Johnstone, S.R.; Best, A.K.; Lee, M.; Barr, K.; Penuela, S.; Laird, D.W.; Isakson, B.E. Expression of pannexin isoforms in the systemic murine arterial network. J. Vasc. Res. 2012, 49, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Haefliger, J.A.; Nicod, P.; Meda, P. Contribution of connexins to the function of the vascular wall. Cardiovasc. Res. 2004, 62, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.E.; Rummery, N.; Hickey, H.; Sandow, S.L. Heterogeneity in the distribution of vascular gap junctions and connexins: Implications for function. Clin. Exp. Pharmacol. Physiol. 2002, 29, 620–625. [Google Scholar] [CrossRef]
- Okamoto, T.; Akiyama, M.; Takeda, M.; Gabazza, E.C.; Hayashi, T.; Suzuki, K. Connexin32 is expressed in vascular endothelial cells and participates in gap-junction intercellular communication. Biochem. Biophys. Res. Commun. 2009, 382, 264–268. [Google Scholar] [CrossRef]
- Severs, N.J.; Rothery, S.; Dupont, E.; Coppen, S.R.; Yeh, H.I.; Ko, Y.S.; Matsushita, T.; Kaba, R.; Halliday, D. Immunocytochemical analysis of connexin expression in the healthy and diseased cardiovascular system. Microsc. Res. Tech. 2001, 52, 301–322. [Google Scholar] [CrossRef]
- Van Kempen, M.J.; ten Velde, I.; Wessels, A.; Oosthoek, P.W.; Gros, D.; Jongsma, H.J.; Moorman, A.F.; Lamers, W.H. Differential connexin distribution accommodates cardiac function in different species. Microsc. Res. Tech. 1995, 31, 420–436. [Google Scholar] [CrossRef]
- Figueroa, X.F.; Isakson, B.E.; Duling, B.R. Connexins: Gaps in our knowledge of vascular function. Physiology 2004, 19, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Gabriels, J.E.; Paul, D.L. Connexin43 is highly localized to sites of disturbed flow in rat aortic endothelium but connexin37 and connexin40 are more uniformly distributed. Circ. Res. 1998, 83, 636–643. [Google Scholar] [CrossRef]
- Kwak, B.R.; Mulhaupt, F.; Veillard, N.; Gros, D.B.; Mach, F. Altered pattern of vascular connexin expression in atherosclerotic plaques. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.M.; McWhorter, A.R. Vascular abnormalities in mice lacking the endothelial gap junction proteins connexin37 and connexin40. Dev. Biol. 2002, 251, 206–220. [Google Scholar] [CrossRef] [Green Version]
- Kruger, O.; Plum, A.; Kim, J.S.; Winterhager, E.; Maxeiner, S.; Hallas, G.; Kirchhoff, S.; Traub, O.; Lamers, W.H.; Willecke, K. Defective vascular development in connexin 45-deficient mice. Development 2000, 127, 4179–4193. [Google Scholar] [CrossRef]
- Li, X.; Simard, J.M. Connexin45 gap junction channels in rat cerebral vascular smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H1890–H1898. [Google Scholar] [CrossRef]
- Li, X.; Simard, J.M. Increase in Cx45 gap junction channels in cerebral smooth muscle cells from SHR. Hypertension 2002, 40, 940–946. [Google Scholar] [CrossRef] [Green Version]
- Moller, S.; Jacobsen, J.C.B.; Holstein-Rathlou, N.H.; Sorensen, C.M. Lack of Connexins 40 and 45 Reduces Local and Conducted Vasoconstrictor Responses in the Murine Afferent Arterioles. Front. Physiol. 2020, 11, 961. [Google Scholar] [CrossRef]
- Figueroa, X.F.; Lillo, M.A.; Gaete, P.S.; Riquelme, M.A.; Saez, J.C. Diffusion of nitric oxide across cell membranes of the vascular wall requires specific connexin-based channels. Neuropharmacology 2013, 75, 471–478. [Google Scholar] [CrossRef]
- Gaete, P.S.; Lillo, M.A.; Ardiles, N.M.; Perez, F.R.; Figueroa, X.F. Ca2+-activated K+ channels of small and intermediate conductance control eNOS activation through NAD(P)H oxidase. Free Radic. Biol. Med. 2012, 52, 860–870. [Google Scholar] [CrossRef]
- Lillo, M.A.; Gaete, P.S.; Puebla, M.; Ardiles, N.M.; Poblete, I.; Becerra, A.; Simon, F.; Figueroa, X.F. Critical contribution of Na(+)-Ca(2+) exchanger to the Ca(2+)-mediated vasodilation activated in endothelial cells of resistance arteries. FASEB J. 2018, 32, 2137–2147. [Google Scholar] [CrossRef] [Green Version]
- De Wit, C.; Roos, F.; Bolz, S.S.; Pohl, U. Lack of vascular connexin 40 is associated with hypertension and irregular arteriolar vasomotion. Physiol. Genom. 2003, 13, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, I.S.; Segal, S.S. Resolution of smooth muscle and endothelial pathways for conduction along hamster cheek pouch arterioles. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H604–H612. [Google Scholar] [CrossRef] [PubMed]
- Budel, S.; Bartlett, I.S.; Segal, S.S. Homocellular conduction along endothelium and smooth muscle of arterioles in hamster cheek pouch: Unmasking an NO wave. Circ. Res. 2003, 93, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, G.G.; Segal, S.S. Endothelial cell pathway for conduction of hyperpolarization and vasodilation along hamster feed artery. Circ. Res. 2000, 86, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Segal, S.S.; Jacobs, T.L. Role for endothelial cell conduction in ascending vasodilatation and exercise hyperaemia in hamster skeletal muscle. J. Physiol. 2001, 536 Pt 3, 937–946. [Google Scholar] [CrossRef]
- Welsh, D.G.; Segal, S.S. Endothelial and smooth muscle cell conduction in arterioles controlling blood flow. Am. J. Physiol. 1998, 274, H178–H186. [Google Scholar] [CrossRef]
- Liao, Y.; Day, K.; Damon, D.; Duling, B.J.P. Endothelial cell-specific knockout of connexin 43 causes hypotension and bradycardia in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 9989–9994. [Google Scholar] [CrossRef] [Green Version]
- Dora, K.; Doyle, M.; Duling, B.J.P. Elevation of intracellular calcium in smooth muscle causes endothelial cell generation of NO in arterioles. Proc. Natl. Acad. Sci. USA 1997, 94, 6529–6534. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, C.F.; Laranjinha, J. Nitric Oxide Pathways in Neurovascular Coupling Under Normal and Stress Conditions in the Brain: Strategies to Rescue Aberrant Coupling and Improve Cerebral Blood Flow. Front. Physiol. 2021, 12, 729201. [Google Scholar] [CrossRef]
- Saliez, J.; Bouzin, C.; Rath, G.; Ghisdal, P.; Desjardins, F.; Rezzani, R.; Rodella, L.F.; Vriens, J.; Nilius, B.; Feron, O.; et al. Role of caveolar compartmentation in endothelium-derived hyperpolarizing factor-mediated relaxation: Ca2+ signals and gap junction function are regulated by caveolin in endothelial cells. Circulation 2008, 117, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; Lin, M.I.; Huang, Y.; Yu, J.; Bauer, P.M.; Giordano, F.J.; Sessa, W.C. Reexpression of caveolin-1 in endothelium rescues the vascular, cardiac, and pulmonary defects in global caveolin-1 knockout mice. J. Exp. Med. 2007, 204, 2373–2382. [Google Scholar] [CrossRef] [Green Version]
- Orellana, J.A.; Saez, P.J.; Cortes-Campos, C.; Elizondo, R.J.; Shoji, K.F.; Contreras-Duarte, S.; Figueroa, V.; Velarde, V.; Jiang, J.X.; Nualart, F.; et al. Glucose increases intracellular free Ca(2+) in tanycytes via ATP released through connexin 43 hemichannels. Glia 2012, 60, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Quan, Y.; Du, Y.; Wu, C.; Gu, S.; Jiang, J.X. Connexin hemichannels regulate redox potential via metabolite exchange and protect lens against cellular oxidative damage. Redox Biol. 2021, 46, 102102. [Google Scholar] [CrossRef]
- Moore, A.R.; Zhou, W.L.; Sirois, C.L.; Belinsky, G.S.; Zecevic, N.; Antic, S.D. Connexin hemichannels contribute to spontaneous electrical activity in the human fetal cortex. Proc. Natl. Acad. Sci. USA 2014, 111, E3919–E3928. [Google Scholar] [CrossRef] [Green Version]
- Feletou, M.; Vanhoutte, P.M. Endothelium-derived hyperpolarizing factor. Clin. Exp. Pharmacol. Physiol. 1996, 23, 1082–1090. [Google Scholar] [CrossRef]
- Morin, E.E.; Salbato, S.; Walker, B.R.; Naik, J.S. Endothelial cell membrane cholesterol content regulates the contribution of TRPV4 channels in ACh-induced vasodilation in rat gracilis arteries. Microcirculation 2022, e12774. [Google Scholar] [CrossRef]
- Vanhoutte, P.M. Endothelium-dependent hyperpolariz.zations: The history. Pharmacol. Res. 2004, 49, 503–508. [Google Scholar] [CrossRef]
- Edwards, G.; Dora, K.A.; Gardener, M.J.; Garland, C.J.; Weston, A.H. K+ is an endothelium-derived hyperpolarizing factor in rat arteries. Nature 1998, 396, 269–272. [Google Scholar] [CrossRef]
- Archer, S.L.; Gragasin, F.S.; Wu, X.; Wang, S.; McMurtry, S.; Kim, D.H.; Platonov, M.; Koshal, A.; Hashimoto, K.; Campbell, W.B.; et al. Endothelium-derived hyperpolarizing factor in human internal mammary artery is 11,12-epoxyeicosatrienoic acid and causes relaxation by activating smooth muscle BK(Ca) channels. Circulation 2003, 107, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I. Cytochrome P450 epoxygenases as EDHF synthase(s). Pharmacol. Res. 2004, 49, 525–533. [Google Scholar] [CrossRef]
- Shimokawa, H.; Morikawa, K. Hydrogen peroxide is an endothelium-derived hyperpolarizing factor in animals and humans. J. Mol. Cell. Cardiol. 2005, 39, 725–732. [Google Scholar] [CrossRef]
- Ahluwalia, A.; Hobbs, A.J. Endothelium-derived C-type natriuretic peptide: More than just a hyperpolarizing factor. Trends Pharmacol. Sci. 2005, 26, 162–167. [Google Scholar] [CrossRef]
- Chauhan, S.D.; Nilsson, H.; Ahluwalia, A.; Hobbs, A.J. Release of C-type natriuretic peptide accounts for the biological activity of endothelium-derived hyperpolarizing factor. Proc. Natl. Acad. Sci. USA 2003, 100, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- Busse, R.; Edwards, G.; Feletou, M.; Fleming, I.; Vanhoutte, P.M.; Weston, A.H. EDHF: Bringing the concepts together. Trends Pharmacol. Sci. 2002, 23, 374–380. [Google Scholar] [CrossRef]
- Doughty, J.M.; Plane, F.; Langton, P.D. Charybdotoxin and apamin block EDHF in rat mesenteric artery if selectively applied to the endothelium. Am. J. Physiol. 1999, 276 Pt 2, H1107–H1112. [Google Scholar] [CrossRef] [PubMed]
- Eichler, I.; Wibawa, J.; Grgic, I.; Knorr, A.; Brakemeier, S.; Pries, A.R.; Hoyer, J.; Kohler, R. Selective blockade of endothelial Ca2+-activated small- and intermediate-conductance K+-channels suppresses EDHF-mediated vasodilation. Br. J. Pharmacol. 2003, 138, 594–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Félétou, M.; Vanhoutte, P.M. Endothelium-dependent hyperpolarization: No longer an f-word! J. Cardiovasc. Pharmacol. 2013, 61, 91–92. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, A.T.; Bakker, L.M.; Edwards, D.H.; Griffith, T.M. Connexin-mimetic peptides dissociate electrotonic EDHF-type signalling via myoendothelial and smooth muscle gap junctions in the rabbit iliac artery. Br. J. Pharmacol. 2005, 144, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vriese, A.S.; Van de Voorde, J.; Lameire, N.H. Effects of connexin-mimetic peptides on nitric oxide synthase- and cyclooxygenase-independent renal vasodilation. Kidney Int. 2002, 61, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagiannis, J.; Rand, M.; Li, C.G. Role of gap junctions in endothelium-derived hyperpolarizing factor-mediated vasodilatation in rat renal artery. Acta Pharmacol. Sin. 2004, 25, 1031–1037. [Google Scholar]
- Begandt, D.; Good, M.E.; Keller, A.S.; DeLalio, L.J.; Rowley, C.; Isakson, B.E.; Figueroa, X.F. Pannexin channel and connexin hemichannel expression in vascular function and inflammation. BMC Cell Biol 2017, 18 (Suppl. 1), 2. [Google Scholar] [CrossRef] [Green Version]
- Billaud, M.; Sandilos, J.K.; Isakson, B.E. Pannexin 1 in the regulation of vascular tone. Trends Cardiovasc. Med. 2012, 22, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Billaud, M.; Lohman, A.W.; Straub, A.C.; Looft-Wilson, R.; Johnstone, S.R.; Araj, C.A.; Best, A.K.; Chekeni, F.B.; Ravichandran, K.S.; Penuela, S.; et al. Pannexin1 regulates alpha1-adrenergic receptor- mediated vasoconstriction. Circ. Res. 2011, 109, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Isakson, B.E.; Thompson, R.J. Pannexin-1 as a potentiator of ligand-gated receptor signaling. Channels 2014, 8, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Good, M.E.; Chiu, Y.H.; Poon, I.K.H.; Medina, C.B.; Butcher, J.T.; Mendu, S.K.; DeLalio, L.J.; Lohman, A.W.; Leitinger, N.; Barrett, E.; et al. Pannexin 1 Channels as an Unexpected New Target of the Anti-Hypertensive Drug Spironolactone. Circ. Res. 2018, 122, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Angus, J.A.; Wright, C.E. Novel alpha1-adrenoceptor antagonism by the fluroquinolone antibiotic trovafloxacin. Eur. J. Pharmacol. 2016, 791, 179–184. [Google Scholar] [CrossRef]
- Lohman, A.W.; Weaver, J.L.; Billaud, M.; Sandilos, J.K.; Griffiths, R.; Straub, A.C.; Penuela, S.; Leitinger, N.; Laird, D.W.; Bayliss, D.A.; et al. S-nitrosylation inhibits pannexin 1 channel function. J. Biol. Chem. 2012, 287, 39602–39612. [Google Scholar] [CrossRef] [Green Version]
- Gaynullina, D.; Shestopalov, V.I.; Panchin, Y.; Tarasova, O.S. Pannexin 1 facilitates arterial relaxation via an endothelium-derived hyperpolarization mechanism. FEBS Lett. 2015, 589, 1164–1170. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, F.A.; Rana, R.; Kim, D.D.; Iwahashi, T.; Zheng, R.; Lal, B.K.; Gordon, D.M.; Meininger, C.J.; Durán, W.N. Internalization of eNOS and NO delivery to subcellular targets determine agonist-induced hyperpermeability. Proc. Natl. Acad. Sci. USA 2009, 106, 6849–6853. [Google Scholar] [CrossRef] [Green Version]
- Claesson-Welsh, L. Vascular permeability—The essentials. Ups. J. Med. Sci. 2015, 120, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Rigor, R.R.; Pivetti, C.D.; Wu, M.H.; Yuan, S.Y. Myosin light chain kinase in microvascular endothelial barrier function. Cardiovasc. Res. 2010, 87, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Tietz, S.; Engelhardt, B. Brain barriers: Crosstalk between complex tight junctions and adherens junctions. J. Cell Biol. 2015, 209, 493–506. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Huang, Y. Mechanisms of Mechanical Force Induced Pulmonary Vascular Endothelial Hyperpermeability. Front. Physiol. 2021, 12, 714064. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Narumi, R.; Akiyama, R.; Vitiello, E.; Shirai, T.; Tanimura, N.; Kuromiya, K.; Ishikawa, S.; Kajita, M.; Tada, M.; et al. Calcium Wave Promotes Cell Extrusion. Curr. Biol. 2020, 30, 670–681e6. [Google Scholar] [CrossRef]
- Matsui, T. Calcium wave propagation during cell extrusion. Curr. Opin. Cell Biol. 2022, 76, 102083. [Google Scholar] [CrossRef]
- Nagy, J.A.; Benjamin, L.; Zeng, H.; Dvorak, A.M.; Dvorak, H.F. Vascular permeability, vascular hyperpermeability and angiogenesis. Angiogenesis 2008, 11, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Joffre, J.; Hellman, J.; Ince, C.; Ait-Oufella, H. Endothelial Responses in Sepsis. Am. J. Respir. Crit. Care Med. 2020, 202, 361–370. [Google Scholar] [CrossRef]
- Claesson-Welsh, L.; Dejana, E.; McDonald, D.M. Permeability of the Endothelial Barrier: Identifying and Reconciling Controversies. Trends Mol. Med. 2021, 27, 314–331. [Google Scholar] [CrossRef]
- Colunga Biancatelli, R.M.L.; Solopov, P.A.; Sharlow, E.R.; Lazo, J.S.; Marik, P.E.; Catravas, J.D. The SARS-CoV-2 spike protein subunit S1 induces COVID-19-like acute lung injury in Kappa18-hACE2 transgenic mice and barrier dysfunction in human endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321, L477–L484. [Google Scholar] [CrossRef]
- Inai, T.; Shibata, Y. Heterogeneous expression of endothelial connexin (Cx) 37, Cx40, and Cx43 in rat large veins. Anat. Sci. Int. 2009, 84, 237–245. [Google Scholar] [CrossRef]
- Chang, C.J.; Wu, L.S.; Hsu, L.A.; Chang, G.J.; Chen, C.F.; Yeh, H.I.; Ko, Y.S. Differential endothelial gap junction expression in venous vessels exposed to different hemodynamics. J. Histochem. Cytochem. 2010, 58, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
- Munger, S.J.; Kanady, J.D.; Simon, A.M. Absence of venous valves in mice lacking Connexin37. Dev. Biol. 2013, 373, 338–348. [Google Scholar] [CrossRef]
- Munger, S.J.; Geng, X.; Srinivasan, R.S.; Witte, M.H.; Paul, D.L.; Simon, A.M. Segregated Foxc2, NFATc1 and Connexin expression at normal developing venous valves, and Connexin-specific differences in the valve phenotypes of Cx37, Cx43, and Cx47 knockout mice. Dev. Biol. 2016, 412, 173–190. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.S.; Li, M.; Sui, B.D.; Wei, L.; Hou, R.; Chen, W.S.; Li, Q.; Bi, S.H.; Zhang, J.Z.; Yi, D.H. Lipopolysaccharide impairs permeability of pulmonary microvascular endothelial cells via Connexin40. Microvasc. Res. 2018, 115, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Deglise, S.; Martin, D.; Probst, H.; Saucy, F.; Hayoz, D.; Waeber, G.; Nicod, P.; Ris, H.B.; Corpataux, J.M.; Haefliger, J.A. Increased connexin43 expression in human saphenous veins in culture is associated with intimal hyperplasia. J. Vasc. Surg. 2005, 41, 1043–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bol, M.; Van Geyt, C.; Baert, S.; Decrock, E.; Wang, N.; De Bock, M.; Gadicherla, A.K.; Randon, C.; Evans, W.H.; Beele, H.; et al. Inhibiting connexin channels protects against cryopreservation-induced cell death in human blood vessels. Eur. J. Vasc. Endovasc. Surg. 2013, 45, 382–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longchamp, A.; Allagnat, F.; Alonso, F.; Kuppler, C.; Dubuis, C.; Ozaki, C.K.; Mitchell, J.R.; Berceli, S.; Corpataux, J.M.; Deglise, S.; et al. Connexin43 Inhibition Prevents Human Vein Grafts Intimal Hyperplasia. PLoS ONE 2015, 10, e0138847. [Google Scholar] [CrossRef] [Green Version]
- Lyons, O.; Saha, P.; Seet, C.; Kuchta, A.; Arnold, A.; Grover, S.; Rashbrook, V.; Sabine, A.; Vizcay-Barrena, G.; Patel, A.; et al. Human venous valve disease caused by mutations in FOXC2 and GJC2. J. Exp. Med. 2017, 214, 2437–2452. [Google Scholar] [CrossRef] [Green Version]
- Chadjichristos, C.E.; Scheckenbach, K.E.; van Veen, T.A.; Richani Sarieddine, M.Z.; de Wit, C.; Yang, Z.; Roth, I.; Bacchetta, M.; Viswambharan, H.; Foglia, B.; et al. Endothelial-specific deletion of connexin40 promotes atherosclerosis by increasing CD73-dependent leukocyte adhesion. Circulation 2010, 121, 123–131. [Google Scholar] [CrossRef] [Green Version]
- De Bock, M.; Culot, M.; Wang, N.; Bol, M.; Decrock, E.; De Vuyst, E.; da Costa, A.; Dauwe, I.; Vinken, M.; Simon, A.M.; et al. Connexin channels provide a target to manipulate brain endothelial calcium dynamics and blood-brain barrier permeability. J. Cereb. Blood Flow Metab. 2011, 31, 1942–1957. [Google Scholar] [CrossRef] [Green Version]
- Shiojiri, N.; Niwa, T.; Sugiyama, Y.; Koike, T. Preferential expression of connexin37 and connexin40 in the endothelium of the portal veins during mouse liver development. Cell Tissue Res. 2006, 324, 547. [Google Scholar] [CrossRef]
- Nagasawa, K.; Chiba, H.; Fujita, H.; Kojima, T.; Saito, T.; Endo, T.; Sawada, N. Possible involvement of gap junctions in the barrier function of tight junctions of brain and lung endothelial cells. J. Cell. Physiol. 2006, 208, 123–132. [Google Scholar] [CrossRef]
- Hwan Seul, K.; Beyer, E.C. Heterogeneous localization of connexin40 in the renal vasculature. Microvasc. Res. 2000, 59, 140–148. [Google Scholar] [CrossRef]
- Yang, X.; Chu, H.; Tang, Y.; Dong, Q. The role of connexin43 in hemorrhagic transformation after thrombolysis in vivo and in vitro. Neuroscience 2016, 329, 54–65. [Google Scholar] [CrossRef]
- Kandasamy, K.; Escue, R.; Manna, J.; Adebiyi, A.; Parthasarathi, K. Changes in endothelial connexin 43 expression inversely correlate with microvessel permeability and VE-cadherin expression in endotoxin-challenged lungs. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L584–L592. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yang, G.M.; Zhu, Y.; Peng, X.Y.; Li, T.; Liu, L.M. Role of connexin 43 in vascular hyperpermeability and relationship to Rock1-MLC20 pathway in septic rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L1323–L1332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Yang, G.; Zhu, Y.; Peng, X.; Li, T.; Liu, L. Relationship of Cx43 regulation of vascular permeability to osteopontin-tight junction protein pathway after sepsis in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R1–R11. [Google Scholar] [CrossRef]
- Tien, T.; Barrette, K.F.; Chronopoulos, A.; Roy, S. Effects of high glucose-induced Cx43 downregulation on occludin and ZO-1 expression and tight junction barrier function in retinal endothelial cells. Invest. Ophthalmol. Vis. Sci. 2013, 54, 6518–6525. [Google Scholar] [CrossRef]
- Kim, D.; Mouritzen, U.; Larsen, B.D.; Roy, S. Inhibition of Cx43 gap junction uncoupling prevents high glucose-induced apoptosis and reduces excess cell monolayer permeability in retinal vascular endothelial cells. Exp. Eye Res. 2018, 173, 85–90. [Google Scholar] [CrossRef]
- Ivanova, E.; Kovacs-Oller, T.; Sagdullaev, B.T. Domain-specific distribution of gap junctions defines cellular coupling to establish a vascular relay in the retina. J. Comp. Neurol. 2019, 527, 2675–2693. [Google Scholar] [CrossRef]
- Liu, X.; Sun, W.; Zhao, Y.; Chen, B.; Wu, W.; Bao, L.; Qi, R. Ginkgolide B Inhibits JAM-A, Cx43, and VE-Cadherin Expression and Reduces Monocyte Transmigration in Oxidized LDL-Stimulated Human Umbilical Vein Endothelial Cells. Oxid. Med. Cell. Longev. 2015, 2015, 907926. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, Y.; Tachikawa, M.; Akaogi, R.; Fujimoto, K.; Ishibashi, M.; Uchida, Y.; Couraud, P.O.; Ohtsuki, S.; Hosoya, K.; Terasaki, T. Contribution of pannexin 1 and connexin 43 hemichannels to extracellular calcium-dependent transport dynamics in human blood-brain barrier endothelial cells. J. Pharmacol. Exp. Ther. 2015, 353, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Griffin, J.M.; Harris, P.W.; Chan, S.H.; Nicholson, L.F.; Brimble, M.A.; O’Carroll, S.J.; Green, C.R. Characterizing the mode of action of extracellular Connexin43 channel blocking mimetic peptides in an in vitro ischemia injury model. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 68–78. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, W.; Hu, X. Different concentrations of lipopolysaccharide regulate barrier function through the PI3K/Akt signalling pathway in human pulmonary microvascular endothelial cells. Sci. Rep. 2018, 8, 9963. [Google Scholar] [CrossRef]
- Ange, M.; Castanares-Zapatero, D.; De Poortere, J.; Dufeys, C.; Courtoy, G.E.; Bouzin, C.; Quarck, R.; Bertrand, L.; Beauloye, C.; Horman, S. alpha1AMP-Activated Protein Kinase Protects against Lipopolysaccharide-Induced Endothelial Barrier Disruption via Junctional Reinforcement and Activation of the p38 MAPK/HSP27 Pathway. Int. J. Mol. Sci. 2020, 21, 5581. [Google Scholar] [CrossRef]
- Saez, J.C.; Contreras-Duarte, S.; Gomez, G.I.; Labra, V.C.; Santibanez, C.A.; Gajardo-Gomez, R.; Avendano, B.C.; Diaz, E.F.; Montero, T.D.; Velarde, V.; et al. Connexin 43 Hemichannel Activity Promoted by Pro-Inflammatory Cytokines and High Glucose Alters Endothelial Cell Function. Front. Immunol. 2018, 9, 1899. [Google Scholar] [CrossRef] [Green Version]
- Tachikawa, M.; Murakami, K.; Akaogi, R.; Akanuma, S.I.; Terasaki, T.; Hosoya, K.I. Polarized hemichannel opening of pannexin 1/connexin 43 contributes to dysregulation of transport function in blood-brain barrier endothelial cells. Neurochem. Int. 2020, 132, 104600. [Google Scholar] [CrossRef]
- Maier-Begandt, D.; Comstra, H.S.; Molina, S.A.; Kruger, N.; Ruddiman, C.A.; Chen, Y.L.; Chen, X.; Biwer, L.A.; Johnstone, S.R.; Lohman, A.W.; et al. A venous-specific purinergic signaling cascade initiated by Pannexin 1 regulates TNFalpha-induced increases in endothelial permeability. Sci. Signal. 2021, 14, eaba2940. [Google Scholar] [CrossRef]
- Good, M.E.; Eucker, S.A.; Li, J.; Bacon, H.M.; Lang, S.M.; Butcher, J.T.; Johnson, T.J.; Gaykema, R.P.; Patel, M.K.; Zuo, Z.; et al. Endothelial cell Pannexin1 modulates severity of ischemic stroke by regulating cerebral inflammation and myogenic tone. JCI Insight 2018, 3, e96272. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.K.; Charles, E.J.; Zhao, Y.; Narahari, A.K.; Baderdinni, P.K.; Good, M.E.; Lorenz, U.M.; Kron, I.L.; Bayliss, D.A.; Ravichandran, K.S.; et al. Pannexin-1 channels on endothelial cells mediate vascular inflammation during lung ischemia-reperfusion injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L301–L312. [Google Scholar] [CrossRef]
- Godecke, S.; Roderigo, C.; Rose, C.R.; Rauch, B.H.; Godecke, A.; Schrader, J. Thrombin-induced ATP release from human umbilical vein endothelial cells. Am. J. Physiol. Cell Physiol. 2012, 302, C915–C923. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Delalio, L.J.; Best, A.K.; Macal, E.; Milstein, J.; Donnelly, I.; Miller, A.M.; McBride, M.; Shu, X.; Koval, M.; et al. Endothelial Pannexin 1 Channels Control Inflammation by Regulating Intracellular Calcium. J. Immunol. 2020, 204, 2995–3007. [Google Scholar] [CrossRef]
- Soon, A.S.; Chua, J.W.; Becker, D.L. Connexins in endothelial barrier function—Novel therapeutic targets countering vascular hyperpermeability. Thromb. Haemost. 2016, 116, 852–867. [Google Scholar] [CrossRef] [Green Version]
- Stamatovic, S.M.; Johnson, A.M.; Keep, R.F.; Andjelkovic, A.V. Junctional proteins of the blood-brain barrier: New insights into function and dysfunction. Tissue Barriers 2016, 4, e1154641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hautefort, A.; Pfenniger, A.; Kwak, B.R. Endothelial connexins in vascular function. Vasc. Biol. 2019, 1, H117–H124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Xu, C.; Wang, S.; Zhang, Y.; Li, W. The Role of Connexin Hemichannels in Inflammatory Diseases. Biology 2022, 11, 237. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, H.; Kobayashi, I.; Kim, D.; Takenaka, H.; Hobson, R.W., 2nd; Durán, W.N. L-type calcium channel blockers modulate the microvascular hyperpermeability induced by platelet-activating factor in vivo. J. Vasc. Surg. 1995, 22, 732–739, discussion 739–741. [Google Scholar] [CrossRef] [Green Version]
- De Bock, M.; Wang, N.; Decrock, E.; Bol, M.; Gadicherla, A.K.; Culot, M.; Cecchelli, R.; Bultynck, G.; Leybaert, L. Endothelial calcium dynamics, connexin channels and blood-brain barrier function. Prog. Neurobiol. 2013, 108, 1–20. [Google Scholar] [CrossRef]
- Vinet, R.; Cortes, M.P.; Alvarez, R.; Delpiano, M.A. Bradykinin and histamine-induced cytosolic calcium increase in capillary endothelial cells of bovine adrenal medulla. Cell Biol. Int. 2014, 38, 1023–1031. [Google Scholar] [CrossRef]
- Brailoiu, E.; Shipsky, M.M.; Yan, G.; Abood, M.E.; Brailoiu, G.C. Mechanisms of modulation of brain microvascular endothelial cells function by thrombin. Brain Res. 2017, 1657, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Brailoiu, E.; Barlow, C.L.; Ramirez, S.H.; Abood, M.E.; Brailoiu, G.C. Effects of Platelet-Activating Factor on Brain Microvascular Endothelial Cells. Neuroscience 2018, 377, 105–113. [Google Scholar] [CrossRef]
- Obata, Y.; Takeuchi, K.; Wei, J.; Hakamata, A.; Odagiri, K.; Nakajima, Y.; Watanabe, H. Interactions between bradykinin and plasmin in the endothelial Ca(2+) response. Mol. Cell. Biochem. 2018, 445, 179–186. [Google Scholar] [CrossRef]
- Dalal, P.J.; Muller, W.A.; Sullivan, D.P. Endothelial Cell Calcium Signaling during Barrier Function and Inflammation. Am. J. Pathol. 2020, 190, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Pocock, T.M.; Williams, B.; Curry, F.E.; Bates, D.O. VEGF and ATP act by different mechanisms to increase microvascular permeability and endothelial [Ca(2+)](i). Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H1625–H1634. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.; Gunduz, D.; Troidl, C.; Heger, J.; Hamm, C.W.; Schulz, R. Purinergic Regulation of Endothelial Barrier Function. Int. J. Mol. Sci. 2021, 22, 1207. [Google Scholar] [CrossRef]
- Parthasarathi, K.; Ichimura, H.; Monma, E.; Lindert, J.; Quadri, S.; Issekutz, A.; Bhattacharya, J. Connexin 43 mediates spread of Ca2+-dependent proinflammatory responses in lung capillaries. J. Clin. Investig. 2006, 116, 2193–2200. [Google Scholar] [CrossRef]
- Parthasarathi, K. Endothelial connexin43 mediates acid-induced increases in pulmonary microvascular permeability. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L33–L42. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, J.J., 3rd; Birukova, A.A.; Beyer, E.C.; Birukov, K.G. Gap junction protein connexin43 exacerbates lung vascular permeability. PLoS ONE 2014, 9, e100931. [Google Scholar] [CrossRef] [Green Version]
- Tsang, H.; Leiper, J.; Hou Lao, K.; Dowsett, L.; Delahaye, M.W.; Barnes, G.; Wharton, J.; Howard, L.; Iannone, L.; Lang, N.N.; et al. Role of asymmetric methylarginine and connexin 43 in the regulation of pulmonary endothelial function. Pulm. Circ. 2013, 3, 675–691. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, J.; Wang, W.; Liu, J.; Zhu, H.; Chen, W.; Chen, T.; Yu, S.; Wang, H.; Sun, G.; et al. Connexin40 modulates pulmonary permeability through gap junction channel in acute lung injury after thoracic gunshot wounds. J. Trauma 2010, 68, 802–809. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, W.; Sun, J.; Li, Q.; Liu, J.; Zhu, H.; Chen, T.; Wang, H.; Yu, S.; Sun, G.; et al. Gap junction channel modulates pulmonary vascular permeability through calcium in acute lung injury: An experimental study. Respiration 2010, 80, 236–245. [Google Scholar] [CrossRef]
- Yin, J.; Lv, L.; Zhai, P.; Long, T.; Zhou, Q.; Pan, H.; Botwe, G.; Wang, L.; Wang, Q.; Tan, L.; et al. Connexin 40 regulates lung endothelial permeability in acute lung injury via the ROCK1-MYPT1- MLC20 pathway. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L35–L44. [Google Scholar] [CrossRef]
- Wang, S.; Sun, Y.; Bai, Y.; Zhou, N.; Chen, N.; Miller, E.J.; Zhang, Y.; Li, W. Contribution of Connexin Hemichannels to the Pathogenesis of Acute Lung Injury. Mediat. Inflamm. 2020, 2020, 8094347. [Google Scholar] [CrossRef]
- Johnson, A.M.; Roach, J.P.; Hu, A.; Stamatovic, S.M.; Zochowski, M.R.; Keep, R.F.; Andjelkovic, A.V. Connexin 43 gap junctions contribute to brain endothelial barrier hyperpermeability in familial cerebral cavernous malformations type III by modulating tight junction structure. FASEB J. 2018, 32, 2615–2629. [Google Scholar] [CrossRef] [Green Version]
- Cronin, M.; Anderson, P.N.; Cook, J.E.; Green, C.R.; Becker, D.L. Blocking connexin43 expression reduces inflammation and improves functional recovery after spinal cord injury. Mol. Cell. Neurosci. 2008, 39, 152–160. [Google Scholar] [CrossRef]
- Yao, F.; Luo, Y.; Chen, Y.; Li, Y.; Hu, X.; You, X.; Li, Z.; Yu, S.; Tian, D.; Zheng, M.; et al. Myelin Debris Impairs Tight Junctions and Promotes the Migration of Microvascular Endothelial Cells in the Injured Spinal Cord. Cell. Mol. Neurobiol. 2022. [Google Scholar] [CrossRef]
- Raghavan, S.; Kenchappa, D.B.; Leo, M.D. SARS-CoV-2 Spike Protein Induces Degradation of Junctional Proteins That Maintain Endothelial Barrier Integrity. Front. Cardiovasc. Med. 2021, 8, 687783. [Google Scholar] [CrossRef]
- Cibelli, A.; Stout, R.; Timmermann, A.; de Menezes, L.; Guo, P.; Maass, K.; Seifert, G.; Steinhauser, C.; Spray, D.C.; Scemes, E. Cx43 carboxyl terminal domain determines AQP4 and Cx30 endfoot organization and blood brain barrier permeability. Sci. Rep. 2021, 11, 24334. [Google Scholar] [CrossRef]
- Tien, T.; Muto, T.; Barrette, K.; Challyandra, L.; Roy, S. Downregulation of Connexin 43 promotes vascular cell loss and excess permeability associated with the development of vascullar lesions in the diabetic retina. Mol. Vis. 2014, 20, 732–741. [Google Scholar]
- Strauss, R.E.; Mezache, L.; Veeraraghavan, R.; Gourdie, R.G. The Cx43 Carboxyl-Terminal Mimetic Peptide alphaCT1 Protects Endothelial Barrier Function in a ZO1 Binding-Competent Manner. Biomolecules 2021, 11, 1192. [Google Scholar] [CrossRef]
- Calder, B.W.; Matthew Rhett, J.; Bainbridge, H.; Fann, S.A.; Gourdie, R.G.; Yost, M.J. Inhibition of connexin 43 hemichannel-mediated ATP release attenuates early inflammation during the foreign body response. Tissue Eng. Part A 2015, 21, 1752–1762. [Google Scholar] [CrossRef] [Green Version]
- Ling, X.; Peng, S.; Xu, Y.; Chu, F. Beneficial effect of simvastatin on human umbilical vein endothelial cells gap junctions induced by TNF-alpha. Anim. Cells Syst. 2022, 26, 10–18. [Google Scholar] [CrossRef]
- Veliz, L.P.; Gonzalez, F.G.; Duling, B.R.; Saez, J.C.; Boric, M.P. Functional role of gap junctions in cytokine-induced leukocyte adhesion to endothelium in vivo. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1056–H1066. [Google Scholar] [CrossRef]
- Fulton, D.; Babbitt, R.; Zoellner, S.; Fontana, J.; Acevedo, L.; McCabe, T.J.; Iwakiri, Y.; Sessa, W.C. Targeting of endothelial nitric-oxide synthase to the cytoplasmic face of the Golgi complex or plasma membrane regulates Akt- versus calcium-dependent mechanisms for nitric oxide release. J. Biol. Chem. 2004, 279, 30349–30357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, T.; Pappas, P.J.; Hobson, R.W., 2nd; Boric, M.P.; Sessa, W.C.; Durán, W.N. Endothelial nitric oxide synthase regulates microvascular hyperpermeability in vivo. J. Physiol. 2006, 574 Pt 1, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, F.A.; Savalia, N.B.; Durán, R.G.; Lal, B.K.; Boric, M.P.; Durán, W.N. Functional significance of differential eNOS translocation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1058–H1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, F.A.; Kim, D.D.; Durán, R.G.; Meininger, C.J.; Durán, W.N. Internalization of eNOS via caveolae regulates PAF-induced inflammatory hyperpermeability to macromolecules. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1642–H1648. [Google Scholar] [CrossRef]
- Durán, W.N.; Breslin, J.W.; Sanchez, F.A. The NO cascade, eNOS location, and microvascular permeability. Cardiovasc. Res. 2010, 87, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Marin, N.; Zamorano, P.; Carrasco, R.; Mujica, P.; Gonzalez, F.G.; Quezada, C.; Meininger, C.J.; Boric, M.P.; Durán, W.N.; Sanchez, F.A. S-Nitrosation of beta-catenin and p120 catenin: A novel regulatory mechanism in endothelial hyperpermeability. Circ. Res. 2012, 111, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Durán, W.N.; Beuve, A.V.; Sanchez, F.A. Nitric oxide, S-nitrosation, and endothelial permeability. IUBMB Life 2013, 65, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, F.A.; Ehrenfeld, I.P.; Durán, W.N. S-nitrosation of proteins: An emergent regulatory mechanism in microvascular permeability and vascular function. Tissue Barriers 2013, 1, e23896. [Google Scholar] [CrossRef] [Green Version]
- Guequen, A.; Carrasco, R.; Zamorano, P.; Rebolledo, L.; Burboa, P.; Sarmiento, J.; Boric, M.P.; Korayem, A.; Durán, W.N.; Sanchez, F.A. S-nitrosylation regulates VE-cadherin phosphorylation and internalization in microvascular permeability. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H1039–H1044. [Google Scholar] [CrossRef] [Green Version]
- Zamorano, P.; Marin, N.; Cordova, F.; Aguilar, A.; Meininger, C.; Boric, M.P.; Golenhofen, N.; Contreras, J.E.; Sarmiento, J.; Durán, W.N.; et al. S-nitrosylation of VASP at cysteine 64 mediates the inflammation-stimulated increase in microvascular permeability. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H66–H71. [Google Scholar] [CrossRef] [Green Version]
- Guequen, A.; Zamorano, P.; Cordova, F.; Koning, T.; Torres, A.; Ehrenfeld, P.; Boric, M.P.; Salazar-Onfray, F.; Gavard, J.; Durán, W.N.; et al. Interleukin-8 Secreted by Glioblastoma Cells Induces Microvascular Hyperpermeability through NO Signaling Involving S-Nitrosylation of VE-Cadherin and p120 in Endothelial Cells. Front. Physiol. 2019, 10, 988. [Google Scholar] [CrossRef] [Green Version]
- Zamorano, P.; Koning, T.; Oyanadel, C.; Mardones, G.A.; Ehrenfeld, P.; Boric, M.P.; Gonzalez, A.; Soza, A.; Sanchez, F.A. Galectin-8 induces endothelial hyperpermeability through the eNOS pathway involving S-nitrosylation-mediated adherens junction disassembly. Carcinogenesis 2019, 40, 313–323. [Google Scholar] [CrossRef]
- Aguilar, G.; Cordova, F.; Koning, T.; Sarmiento, J.; Boric, M.P.; Birukov, K.; Cancino, J.; Varas-Godoy, M.; Soza, A.; Alves, N.G.; et al. TNF-alpha-activated eNOS signaling increases leukocyte adhesion through the S-nitrosylation pathway. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H1083–H1095. [Google Scholar] [CrossRef]
- Lan, S.H.; Lai, W.T.; Zheng, S.Y.; Yang, L.; Fang, L.C.; Zhou, L.; Tang, B.; Duan, J.; Hong, T. Upregulation of Connexin 40 Mediated by Nitric Oxide Attenuates Cerebral Vasospasm After Subarachnoid Hemorrhage via the Nitric Oxide-Cyclic Guanosine Monophosphate-Protein Kinase G Pathway. World Neurosurg. 2020, 136, e476–e486. [Google Scholar] [CrossRef]
- Sun, J.; Nguyen, T.; Aponte, A.M.; Menazza, S.; Kohr, M.J.; Roth, D.M.; Patel, H.H.; Murphy, E.; Steenbergen, C. Ischaemic preconditioning preferentially increases protein S-nitrosylation in subsarcolemmal mitochondria. Cardiovasc. Res. 2015, 106, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Kameritsch, P.; Pogoda, K. The Role of Connexin 43 and Pannexin 1 During Acute Inflammation. Front. Physiol. 2020, 11, 594097. [Google Scholar] [CrossRef]
- Bunse, S.; Schmidt, M.; Prochnow, N.; Zoidl, G.; Dermietzel, R. Intracellular cysteine 346 is essentially involved in regulating Panx1 channel activity. J. Biol. Chem. 2010, 285, 38444–38452. [Google Scholar] [CrossRef] [Green Version]
- Penuela, S.; Simek, J.; Thompson, R.J. Regulation of pannexin channels by post-translational modifications. FEBS Lett. 2014, 588, 1411–1415. [Google Scholar] [CrossRef] [Green Version]
- Poornima, V.; Vallabhaneni, S.; Mukhopadhyay, M.; Bera, A.K. Nitric oxide inhibits the pannexin 1 channel through a cGMP-PKG dependent pathway. Nitric Oxide 2015, 47, 77–84. [Google Scholar] [CrossRef]
- Filosa, J.A.; Blanco, V.M. Neurovascular coupling in the mammalian brain. Exp. Physiol. 2007, 92, 641–646. [Google Scholar] [CrossRef]
- Phillips, A.A.; Chan, F.H.; Zheng, M.M.Z.; Krassioukov, A.V.; Ainslie, P.N. Neurovascular coupling in humans: Physiology, methodological advances and clinical implications. J. Cereb. Blood Flow Metab. 2016, 36, 647–664. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Pan, Y.; Harris, S.; Billings, S.; Coca, D.; Berwick, J.; Jones, M.; Kennerley, A.; Johnston, D.; Martin, C.; et al. A dynamic model of neurovascular coupling: Implications for blood vessel dilation and constriction. NeuroImage 2010, 52, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.H.; Miller, S.L.; Castillo-Melendez, M.; Malhotra, A. The Neurovascular Unit: Effects of Brain Insults During the Perinatal Period. Front. Neurosci. 2020, 13, 1452. [Google Scholar] [CrossRef]
- Muoio, V.; Persson, P.B.; Sendeski, M.M. The neurovascular unit—Concept review. Acta Physiol. 2014, 210, 790–798. [Google Scholar] [CrossRef]
- Netto, J.P.; Iliff, J.; Stanimirovic, D.; Krohn, K.A.; Hamilton, B.; Varallyay, C.; Gahramanov, S.; Daldrup-Link, H.; D’Esterre, C.; Zlokovic, B.; et al. Neurovascular unit: Basic and clinical imaging with emphasis on advantages of ferumoxytol. Neurosurgery 2018, 82, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Filosa, J.A. Vascular tone and neurovascular coupling: Considerations toward an improved in vitro model. Front. Neuroenergetics 2010, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Girouard, H.; Iadecola, C. Neurovascular coupling in the normal brain and in hypertension, stroke, and Alzheimer disease. J. Appl. Physiol. 2006, 100, 328–335. [Google Scholar] [CrossRef]
- Filosa, J.A.; Iddings, J.A. Astrocyte regulation of cerebral vascular tone. Am. J. Physiol.-Heart Circ. Physiol. 2013, 305, H609–H619. [Google Scholar] [CrossRef] [Green Version]
- Gordon, G.R.J.; Mulligan, S.J.; MacVicar, B.A. Astrocyte Control of the Cerebrovasculature GRANT. Glia 2007, 55, 1214–1221. [Google Scholar] [CrossRef]
- Haydon, P.G.; Carmignoto, G. Astrocyte control of synaptic transmission and neurovascular coupling. Physiol. Rev. 2006, 86, 1009–1031. [Google Scholar] [CrossRef] [Green Version]
- Nuriya, M.; Hirase, H. Involvement of astrocytes in neurovascular communication. Prog. Brain Res. 2016, 225, 41–62. [Google Scholar] [PubMed]
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Butt, A.M. ATP: A ubiquitous gliotransmitter integrating neuron-glial networks. Semin. Cell Dev. Biol. 2011, 22, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Filosa, J.A.; Bonev, A.D.; Nelson, M.T. Calcium dynamics in cortical astrocytes and arterioles during neurovascular coupling. Circ. Res. 2004, 95, e73–e81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girouard, H.; Bonev, A.D.; Hannah, R.M.; Meredith, A.; Aldrich, R.W.; Nelson, M.T. Astrocytic endfoot Ca2+ and BK channels determine both arteriolar dilation and constriction. Proc. Natl. Acad. Sci. USA 2010, 107, 3811–3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarrete, M.; Perea, G.; Maglio, L.; Pastor, J.; García De Sola, R.; Araque, A. Astrocyte calcium signal and gliotransmission in human brain tissue. Cereb. Cortex 2013, 23, 1240–1246. [Google Scholar] [CrossRef] [Green Version]
- Otsu, Y.; Couchman, K.; Lyons, D.G.; Collot, M.; Agarwal, A.; Mallet, J.-M.; Pfrieger, F.W.; Bergles, D.E.; Charpak, S. Calcium dynamics in astrocyte processes during neurovascular coupling. Nat. Neurosci. 2015, 18, 210–218. [Google Scholar] [CrossRef]
- Straub, S.V.; Nelson, M.T. Astrocytic Calcium Signaling: The Information Currency Coupling Neuronal Activity to the Cerebral Microcirculation. Trends Cardiovasc. Med. 2007, 17, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.L.; Pelligrino, D.A. ATP release and hydrolysis contribute to rat pial arteriolar dilatation elicited by neuronal activation. Exp. Physiol. 2007, 92, 647–651. [Google Scholar] [CrossRef] [Green Version]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [Green Version]
- Petroff, O.A.C. Book Review: GABA and Glutamate in the Human Brain. Neuroscience 2002, 8, 562–573. [Google Scholar] [CrossRef]
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J. Neural Transm. 2014, 121, 799–817. [Google Scholar] [CrossRef] [Green Version]
- Bradley, S.J.; Challiss, R.A.J. G protein-coupled receptor signalling in astrocytes in health and disease: A focus on metabotropic glutamate receptors. Biochem. Pharmacol. 2012, 84, 249–259. [Google Scholar] [CrossRef] [Green Version]
- D’Antoni, S.; Berretta, A.; Bonaccorso, C.M.; Bruno, V.; Aronica, E.; Nicoletti, F.; Catania, M.V. Metabotropic Glutamate Receptors in Glial Cells. Neurochem. Res. 2008, 33, 2436–2443. [Google Scholar] [CrossRef]
- Seifert, G.; Steinhäuser, C. Ionotropic glutamate receptors in astrocytes. Prog. Brain Res. 2001, 132, 287–299. [Google Scholar]
- Verkhratsky, A.; Chvátal, A. NMDA Receptors in Astrocytes. Neurochem. Res. 2020, 45, 122–133. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Linden, D.J.; Sapirstein, A. Astrocyte Inositol Triphosphate Receptor Type 2 and Cytosolic Phospholipase A2 Alpha Regulate Arteriole Responses in Mouse Neocortical Brain Slices. PLoS ONE 2012, 7, e42194. [Google Scholar] [CrossRef] [Green Version]
- Zur Nieden, R.; Deitmer, J.W. The Role of Metabotropic Glutamate Receptors for the Generation of Calcium Oscillations in Rat Hippocampal Astrocytes in situ. Cereb. Cortex 2006, 16, 676–687. [Google Scholar] [CrossRef]
- Stackhouse, T.L.; Mishra, A. Neurovascular Coupling in Development and Disease: Focus on Astrocytes. Front. Cell Dev. Biol. 2021, 9, 1745. [Google Scholar] [CrossRef]
- Dunn, K.M.; Nelson, M.T. Potassium channels and neurovascular coupling. Circ. J. 2010, 74, 608–616. [Google Scholar] [CrossRef] [Green Version]
- Gordon, G.R.J.; Howarth, C.; Macvicar, B.A. Bidirectional control of arteriole diameter by astrocytes. Exp. Physiol. 2011, 96, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, C.; Gebremedhin, D.; Hwang, S.H.; Hammock, B.D.; Falck, J.R.; Roman, R.J.; Harder, D.R.; Koehler, R.C. Epoxyeicosatrienoic acid-dependent cerebral vasodilation evoked by metabotropic glutamate receptor activation in vivo. Am. J. Physiol.-Heart Circ. Physiol. 2011, 301, H373–H381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medhora, M. Dual Regulation of the Cerebral Microvasculature by Epoxyeicosatrienoic Acids. Trends Cardiovasc. Med. 2001, 11, 38–42. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, X.; Gebremedhin, D.; Falck, J.R.; Harder, D.R.; Koehler, R.C. Interaction of Mechanisms Involving Epoxyeicosatrienoic Acids, Adenosine Receptors, and Metabotropic Glutamate Receptors in Neurovascular Coupling in Rat Whisker Barrel Cortex. J. Cereb. Blood Flow Metab. 2008, 28, 111–125. [Google Scholar] [CrossRef]
- Earley, S.; Brayden, J.E. Transient Receptor Potential Channels in the Vasculature. Physiol. Rev. 2015, 95, 645–690. [Google Scholar] [CrossRef] [Green Version]
- Iliff, J.J.; Jia, J.; Nelson, J.; Goyagi, T.; Klaus, J.; Alkayed, N.J. Epoxyeicosanoid signaling in CNS function and disease. Prostaglandins Other Lipid Mediat. 2010, 91, 68–84. [Google Scholar] [CrossRef] [Green Version]
- Filosa, J.A.; Morrison, H.W.; Iddings, J.A.; Du, W.; Kim, K.J. Beyond neurovascular coupling, role of astrocytes in the regulation of vascular tone. Neuroscience 2016, 323, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Imig, J.D.; Simpkins, A.N.; Renic, M.; Harder, D.R. Cytochrome P450 eicosanoids and cerebral vascular function. Expert Rev. Mol. Med. 2011, 13, e7. [Google Scholar] [CrossRef]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; MacVicar, B.A.; Newman, E.A. Glial and neuronal control of brain blood flow. Nature 2010, 468, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Filosa, J.A.; Bonev, A.D.; Straub, S.V.; Meredith, A.L.; Wilkerson, M.K.; Aldrich, R.W.; Nelson, M.T. Local potassium signaling couples neuronal activity to vasodilation in the brain. Nat. Neurosci. 2006, 9, 1397–1403. [Google Scholar] [CrossRef]
- Koizumi, S. Synchronization of Ca2+ oscillations: Involvement of ATP release in astrocytes. FEBS J 2010, 277, 286–292. [Google Scholar] [CrossRef]
- Farr, H.; David, T. Models of neurovascular coupling via potassium and EET signalling. J. Theor. Biol. 2011, 286, 13–23. [Google Scholar] [CrossRef]
- Macvicar, B.A.; Newman, E.A. Astrocyte regulation of blood flow in the brain. Cold Spring Harb. Perspect. Biol. 2015, 7, a020388. [Google Scholar] [CrossRef]
- Gordon, G.R.J.; Choi, H.B.; Rungta, R.L.; Ellis-Davies, G.C.R.; MacVicar, B.A. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature 2008, 456, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Metea, M.R. Glial Cells Dilate and Constrict Blood Vessels: A Mechanism of Neurovascular Coupling. J. Neurosci. 2006, 26, 2862–2870. [Google Scholar] [CrossRef]
- Roman, R.J. Molecular mechanisms and cell signaling of 20-hydroxyeicosatetraenoic acid in vascular pathophysiology. Front. Biosci. 2016, 21, 4465. [Google Scholar] [CrossRef] [Green Version]
- Newman, E.A. Glial cell regulation of neuronal activity and blood flow in the retina by release of gliotransmitters. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140195. [Google Scholar] [CrossRef]
- Pelligrino, D.A.; Vetri, F.; Xu, H.L. Purinergic mechanisms in gliovascular coupling. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2011; Volume 22, pp. 229–236. [Google Scholar]
- Wells, J.A.; Christie, I.N.; Hosford, P.S.; Huckstepp, R.T.R.; Angelova, P.R.; Vihko, P.; Cork, S.C.; Abramov, A.Y.; Teschemacher, A.G.; Kasparov, S.; et al. A Critical Role for Purinergic Signalling in the Mechanisms Underlying Generation of BOLD fMRI Responses. J. Neurosci. 2015, 35, 5284–5292. [Google Scholar] [CrossRef] [Green Version]
- Hørlyck, S.; Cai, C.; Helms, H.C.C.; Lauritzen, M.; Brodin, B. ATP induces contraction of cultured brain capillary pericytes via activation of P2Y-type purinergic receptors. Am. J. Physiol.-Heart Circ. Physiol. 2021, 320, H699–H712. [Google Scholar] [CrossRef]
- Li, Y.; Baylie, R.L.; Tavares, M.J.; Brayden, J.E. TRPM4 Channels Couple Purinergic Receptor Mechanoactivation and Myogenic Tone Development in Cerebral Parenchymal Arterioles. J. Cereb. Blood Flow Metab. 2014, 34, 1706–1714. [Google Scholar] [CrossRef] [Green Version]
- Beamer, E.; Conte, G.; Engel, T. ATP release during seizures—A critical evaluation of the evidence. Brain Res. Bull. 2019, 151, 65–73. [Google Scholar] [CrossRef]
- Dunn, K.M.; Nelson, M.T. Neurovascular signaling in the brain and the pathological consequences of hypertension. Am. J. Physiol.-Heart Circ. Physiol. 2014, 306, H1–H14. [Google Scholar] [CrossRef] [Green Version]
- Fitz, J.G. Regulation of cellular ATP release. Trans. Am. Clin. Climatol. Assoc. 2007, 118, 199–208. [Google Scholar]
- Vetri, F.; Xu, H.; Mao, L.; Paisansathan, C.; Pelligrino, D.A. ATP hydrolysis pathways and their contributions to pial arteriolar dilation in rats. Am. J. Physiol.-Heart Circ. Physiol. 2011, 301, H1369–H1377. [Google Scholar] [CrossRef] [Green Version]
- Braun, N.; Sévigny, J.; Robson, S.C.; Enjyoji, K.; Guckelberger, O.; Hammer, K.; Di Virgilio, F.; Zimmermann, H. Assignment of ecto-nucleoside triphosphate diphosphohydrolase-1/cd39 expression to microglia and vasculature of the brain. Eur. J. Neurosci. 2000, 12, 4357–4366. [Google Scholar]
- Grković, I.; Drakulić, D.; Martinović, J.; Mitrović, N. Role of Ectonucleotidases in Synapse Formation During Brain Development: Physiological and Pathological Implications. Curr. Neuropharmacol. 2018, 17, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.M.; Buchakjian, M.R.; Dubyak, G.R. Colocalization of ATP Release Sites and Ecto-ATPase Activity at the Extracellular Surface of Human Astrocytes. J. Biol. Chem. 2003, 278, 23331–23342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, K.A.; Gao, Z.-G. Adenosine receptors as therapeutic targets. Nat. Rev. Drug Discov. 2006, 5, 247–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusano, Y.; Echeverry, G.; Miekisiak, G.; Kulik, T.B.; Aronhime, S.N.; Chen, J.F.; Winn, H.R. Role of Adenosine A2 Receptors in Regulation of Cerebral Blood Flow during Induced Hypotension. J. Cereb. Blood Flow Metab. 2010, 30, 808–815. [Google Scholar] [CrossRef]
- Ngai, A.C.; Coyne, E.F.; Meno, J.R.; West, G.A.; Winn, H.R. Receptor subtypes mediating adenosine-induced dilation of cerebral arterioles. Am. J. Physiol.-Heart Circ. Physiol. 2001, 280, H2329–H2335. [Google Scholar] [CrossRef]
- Paisansathan, C.; Xu, H.; Vetri, F.; Hernandez, M.; Pelligrino, D.A. Interactions between adenosine and K+ channel-related pathways in the coupling of somatosensory activation and pial arteriolar dilation. Am. J. Physiol.-Heart Circ. Physiol. 2010, 299, H2009–H2017. [Google Scholar] [CrossRef] [Green Version]
- Rosenblum, W.I. ATP-Sensitive Potassium Channels in the Cerebral Circulation. Stroke 2003, 34, 1547–1552. [Google Scholar] [CrossRef] [Green Version]
- Hein, T.W.; Xu, W.; Ren, Y.; Kuo, L. Cellular signalling pathways mediating dilation of porcine pial arterioles to adenosine A2A receptor activation. Cardiovasc. Res. 2013, 99, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Mills, J.H.; Alabanza, L.; Weksler, B.B.; Couraud, P.-O.; Romero, I.A.; Bynoe, M.S. Human brain endothelial cells are responsive to adenosine receptor activation. Purinergic Signal. 2011, 7, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.K.; Shin, Y.W.; Hong, K.W. Role of adenosine A 2B receptors in vasodilation of rat pial artery and cerebral blood flow autoregulation. Am. J. Physiol.-Heart Circ. Physiol. 2000, 278, H339–H344. [Google Scholar] [CrossRef]
- Sáez, J.C.; Nicholson, B. Connexin and Pannexin Based Channels in the Nervous System. In From Molecules to Networks; Elsevier: Amsterdam, The Netherlands, 2014; pp. 257–283. [Google Scholar]
- Kar, R.; Batra, N.; Riquelme, M.A.; Jiang, J.X. Biological role of connexin intercellular channels and hemichannels. Arch Biochem Biophys 2012, 524, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Giaume, C.; Theis, M. Pharmacological and genetic approaches to study connexin-mediated channels in glial cells of the central nervous system. Brain Res. Rev. 2010, 63, 160–176. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J.I.; Ionescu, A.V.; Lynn, B.D.; Rash, J.E. Coupling of astrocyte connexins Cx26, Cx30, Cx43 to oligodendrocyte Cx29, Cx32, Cx47: Implications from normal and connexin32 knockout mice. Glia 2003, 44, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.I.; Lynn, B.D.; Tress, O.; Willecke, K.; Rash, J.E. Connexin26 expression in brain parenchymal cells demonstrated by targeted connexin ablation in transgenic mice. Eur. J. Neurosci. 2011, 34, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, B.S.; Hansen, D.B.; Ransom, B.R.; Nielsen, M.S.; MacAulay, N. Connexin Hemichannels in Astrocytes: An Assessment of Controversies Regarding Their Functional Characteristics. Neurochem. Res. 2017, 42, 2537–2550. [Google Scholar] [CrossRef]
- Xing, L.; Yang, T.; Cui, S.; Chen, G. Connexin Hemichannels in Astrocytes: Role in CNS Disorders. Front. Mol. Neurosci. 2019, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bock, M.; Decrock, E.; Wang, N.; Bol, M.; Vinken, M.; Bultynck, G.; Leybaert, L. The dual face of connexin-based astroglial Ca2+ communication: A key player in brain physiology and a prime target in pathology. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2014, 1843, 2211–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Kiyoshi, C.M.; Terman, D.; Zhou, M. Analysis of the Functional States of an Astrocyte Syncytium. In Basic Neurobiology Techniques; Wright, N., Ed.; Neuromethods; Springer: New York, NY, USA, 2020; Volume 152, pp. 285–313. [Google Scholar]
- Pacholko, A.G.; Wotton, C.A.; Bekar, L.K. Astrocytes—The Ultimate Effectors of Long-Range Neuromodulatory Networks? Front. Cell. Neurosci. 2020, 14, 581075. [Google Scholar] [CrossRef]
- Ahmadpour, N.; Kantroo, M.; Stobart, J.L. Extracellular Calcium Influx Pathways in Astrocyte Calcium Microdomain Physiology. Biomolecules 2021, 11, 1467. [Google Scholar] [CrossRef]
- Lagos-Cabré, R.; Brenet, M.; Díaz, J.; Pérez, R.; Pérez, L.; Herrera-Molina, R.; Quest, A.; Leyton, L. Intracellular Ca2+ Increases and Connexin 43 Hemichannel Opening Are Necessary but Not Sufficient for Thy-1-Induced Astrocyte Migration. Int. J. Mol. Sci. 2018, 19, 2179. [Google Scholar] [CrossRef] [Green Version]
- Turovsky, E.A.; Braga, A.; Yu, Y.; Esteras, N.; Korsak, A.; Theparambil, S.M.; Hadjihambi, A.; Hosford, P.S.; Teschemacher, A.G.; Marina, N.; et al. Mechanosensory Signaling in Astrocytes. J. Neurosci. 2020, 40, 9364–9371. [Google Scholar] [CrossRef]
- Kang, J.; Kang, N.; Lovatt, D.; Torres, A.; Zhao, Z.; Lin, J.; Nedergaard, M. Connexin 43 Hemichannels Are Permeable to ATP. J. Neurosci. 2008, 28, 4702–4711. [Google Scholar] [CrossRef]
- Stout, C.E.; Costantin, J.L.; Naus, C.C.G.; Charles, A.C. Intercellular Calcium Signaling in Astrocytes via ATP Release through Connexin Hemichannels. J. Biol. Chem. 2002, 277, 10482–10488. [Google Scholar] [CrossRef] [Green Version]
- Haddock, R.E.; Grayson, T.H.; Brackenbury, T.D.; Meaney, K.R.; Neylon, C.B.; Sandow, S.L.; Hill, C.E. Endothelial coordination of cerebral vasomotion via myoendothelial gap junctions containing connexins 37 and 40. Am. J. Physiol.-Heart Circ. Physiol. 2006, 291, 2047–2056. [Google Scholar] [CrossRef]
- Avila, M.A.; Sell, S.L.; Hawkins, B.E.; Hellmich, H.L.; Boone, D.R.; Crookshanks, J.M.; Prough, D.S.; DeWitt, D.S. Cerebrovascular connexin expression: Effects of traumatic brain injury. J. Neurotrauma 2011, 28, 1803–1811. [Google Scholar] [CrossRef] [Green Version]
- Brisset, A.C.; Isakson, B.E.; Kwak, B.R. Connexins in vascular physiology and pathology. Antioxid. Redox Signal. 2009, 11, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xin, Y.; He, Z.; Hu, W. Function of Connexins in the Interaction between Glial and Vascular Cells in the Central Nervous System and Related Neurological Diseases. Neural Plast. 2018, 2018, 6323901. [Google Scholar] [CrossRef] [PubMed]
- Bruzzone, R.; Hormuzdi, S.G.; Barbe, M.T.; Herb, A.; Monyer, H. Pannexins, a family of gap junction proteins expressed in brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13644–13649. [Google Scholar] [CrossRef] [Green Version]
- Yeung, A.K.; Patil, C.S.; Jackson, M.F. Pannexin-1 in the CNS: Emerging concepts in health and disease. J. Neurochem. 2020, 154, 468–485. [Google Scholar] [CrossRef]
- Giaume, C.; Naus, C.C.; Sáez, J.C.; Leybaert, L. Glial Connexins and Pannexins in the Healthy and Diseased Brain. Physiol. Rev. 2021, 101, 93–145. [Google Scholar] [CrossRef]
- Huang, Y.; Grinspan, J.B.; Abrams, C.K.; Scherer, S.S. Pannexin1 is expressed by neurons and glia but does not form functional gap junctions. Glia 2007, 55, 46–56. [Google Scholar] [CrossRef]
- MacVicar, B.A.; Thompson, R.J. Non-junction functions of pannexin-1 channels. Trends Neurosci. 2010, 33, 93–102. [Google Scholar] [CrossRef]
- Iglesias, R.; Dahl, G.; Qiu, F.; Spray, D.C.; Scemes, E. Pannexin 1: The Molecular Substrate of Astrocyte “Hemichannels”. J. Neurosci. 2009, 29, 7092–7097. [Google Scholar] [CrossRef]
- Suadicani, S.O.; Iglesias, R.; Wang, J.; Dahl, G.; Spray, D.C.; Scemes, E. ATP signaling is deficient in cultured pannexin1-null mouse astrocytes. Glia 2012, 60, 1106–1116. [Google Scholar] [CrossRef] [Green Version]
- Dahl, G. ATP release through pannexon channels. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140191. [Google Scholar] [CrossRef]
- Scemes, E.; Spray, D.C. Extracellular K+ and Astrocyte Signaling via Connexin and Pannexin Channels. Neurochem. Res. 2012, 37, 2310–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Protein | Species | Blood Vessel | Endothelial Cell | Smooth Muscle Cell | Reference |
|---|---|---|---|---|---|
| Rat | Coronary artery | + | + | [66] | |
| Cx37 | Arterial mesenteric bed | + | − | [67] | |
| Mouse | Cremaster muscle | + | − | [12] | |
| Hamster | Cheek pouch arterioles, retractor muscle feed arteries | + | + | [67] | |
| Pig | Coronary arteries | + | − | [66] | |
| Rat | Coronary arteries | + | + | [66] | |
| Cx40 | Arterial mesenteric bed | + | − | [67] | |
| Brain Pial arterioles | + | + | [67] | ||
| Mouse | Cremaster muscle | + | + | [12,68] | |
| Hamster | Cheek pouch arterioles, retractor muscle feed arteries | + | − | [67] | |
| Pig | Coronary artery | + | + | [66] | |
| Cow | Coronary artery | + | + | [66] | |
| Cx43 | Rat | Coronary artery | − | + | [66] |
| Arterial mesenteric bed | + | − | [67] | ||
| Mouse | Cremaster muscle | + | + | [12,68] | |
| Hamster | Cheek pouch arterioles, retractor muscle feed arteries | + | − | [67] | |
| Panx-1 | Rat | Arterial mesenteric bed | + | + | [69,70] |
| Mouse | Pinotrapezius muscle and cremaster muscle arterioles | + | + | [71] | |
| Coronary arteries | + | − | [71] | ||
| Abdominal arteries | + | + | [71] | ||
| Panx-3 | Mouse | Coronary arteries | + | + | [71] |
| Cortical kidney arterioles | + | − | [71] |
| Protein | Species | Blood Vessel or Cell Culture | Endothelial Cell | Valves | Inflammatory Model | Reference |
|---|---|---|---|---|---|---|
| Cx37 | Rat | Vena cava | + | − | − | [137,138] |
| Rat brain endothelial (RBE4) | + | − | Bradykinin acute inflammation | [147] | ||
| Pulmonary microvascular endothelial cells (PMVEC) | + | − | In vitro acute lung injury | [141] | ||
| Mouse | Vena cava, Femoral vein, Saphenous vein, Braquial vein | − | + | − | [139,140,145] | |
| Portal vein | + | − | Mouse liver development | [148] | ||
| Human | Saphenous vein | + | − | In vitro vein hyperplasia, cryopreservation | [142,143,144] | |
| Saphenous vein | − | + | − | [145] | ||
| Rabbit | Yugular vein | + | − | − | [144] | |
| Pig | Primary culture of porcine brain microvascular endothelial cells | + | - | − | [149] | |
| Cx40 | Rat | Vena cava | + | − | - | [137,138] |
| Rat brain endothelial (RBE4) | + | − | Bradykinin acute inflammation | [147] | ||
| Pulmonary microvascular endothelial cells (PMVEC) | + | − | In vitro acute lung injury | [141] | ||
| Mouse | Vena cava, Femoral vein, Saphenous vein, Braquial vein | − | + | − | [139,140,145] | |
| Portal vein | + | − | Mouse liver development | [148] | ||
| Renal Interlobular veins | + | − | − | [150] | ||
| Human | Saphenous vein | + | − | In vitro vein hyperplasia | [142,143,144] | |
| Saphenous vein | − | + | − | [145] | ||
| Rabbit | Yugular vein | + | − | − | [144] | |
| Pig | Primary culture of porcine brain microvascular endothelial cells | + | − | − | [149] | |
| Cx43 | Rat | Vena cava | + | − | − | [137,138] |
| Rat brain endothelial (RBE4) | + | − | Bradykinin acute inflammation | [147] | ||
| Rat Brain microvascular endothelial cells (RBMEC) | + | − | Thrombolysis | [151] | ||
| Pulmonary venules | + | − | Endotoxin-challenged lungs | [152] | ||
| Pulmonary vein endothelial cells | + | − | Sepsis | [153,154] | ||
| Retinal endothelial cells | + | − | In vitro diabetic retinopathy | [155,156] | ||
| Pulmonary microvascular endothelial cells (PMVEC) | + | − | In vitro acute lung injury | [141] | ||
| Mouse | Vena cava, Femoral vein, Saphenous vein, Braquial vein | − | + | − | [139,140,145] | |
| Portal vein | + | − | Mouse liver development | [148] | ||
| Retinal vein | + | − | − | [157] | ||
| Human | Saphenous vein | + | − | In vitro vein hyperplasia | [142,143,144] | |
| Saphenous vein | − | + | − | [145] | ||
| HUVEC | + | − | In vitro atherosclerosis | [158] | ||
| Human microvascular endothelial cell line (hCMVEC) | + | − | In vitro Ischemia injury and Extracellular Ca2+ variations | [159,160] | ||
| Human pulmonary microvascular endothelial cells | + | − | Acute lung injury | [161] | ||
| Human dermal microvascular endothelial cells (HMEC) | + | − | In vitro Sepsis | [162] | ||
| EAhy Cells | + | − | Inflammatory cytokines (TNF-α, IL-8) and high glucose exposure | [28,163] | ||
| Rabbit | Yugular vein | + | − | − | [144] | |
| Pig | Primary culture of porcine brain microvascular endothelial cells | + | − | − | [149] | |
| Cx47 | Mouse | Vena cava, Femoral vein, Saphenous vein, Braquial vein | − | + | Vein development | [139,140,145] |
| Human | Saphenous vein | + | + | In vitro vein hyperplasia, vein development | [142,145] | |
| Protein | Species | Blood Vessel or Cell Culture | Endothelial Cell | Valves | Inflammatory Condition | Reference |
| Panx-1 | Rat | Rat Brain Capillary endothelial cells TR-BBB13 | + | − | In vitro brain ischemia | [164] |
| Mouse | Mesenteric postcapillary venules | + | − | TNF-α acute inflammation | [65,165,166,167] | |
| Human | HUVEC | + | − | Thrombin, TNF-α inflammation | [165,168,169] | |
| Human microvascular endothelial cell line (hCMVEC) | + | − | Extracellular Ca2+ variations | [159,164] | ||
| Primary Human Saphenous vein Endothelial cells | + | − | TNF-α inflammation | [165] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burboa, P.C.; Puebla, M.; Gaete, P.S.; Durán, W.N.; Lillo, M.A. Connexin and Pannexin Large-Pore Channels in Microcirculation and Neurovascular Coupling Function. Int. J. Mol. Sci. 2022, 23, 7303. https://doi.org/10.3390/ijms23137303
Burboa PC, Puebla M, Gaete PS, Durán WN, Lillo MA. Connexin and Pannexin Large-Pore Channels in Microcirculation and Neurovascular Coupling Function. International Journal of Molecular Sciences. 2022; 23(13):7303. https://doi.org/10.3390/ijms23137303
Chicago/Turabian StyleBurboa, Pía C., Mariela Puebla, Pablo S. Gaete, Walter N. Durán, and Mauricio A. Lillo. 2022. "Connexin and Pannexin Large-Pore Channels in Microcirculation and Neurovascular Coupling Function" International Journal of Molecular Sciences 23, no. 13: 7303. https://doi.org/10.3390/ijms23137303
APA StyleBurboa, P. C., Puebla, M., Gaete, P. S., Durán, W. N., & Lillo, M. A. (2022). Connexin and Pannexin Large-Pore Channels in Microcirculation and Neurovascular Coupling Function. International Journal of Molecular Sciences, 23(13), 7303. https://doi.org/10.3390/ijms23137303