
The Analysis of a Genome-Wide Association Study (GWAS) of Overweight and Obesity in Psoriasis

 , ,
, ,  , , ,
, , ,  ,
,  ,
,  ,
,  ,
, 
 ,
,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. The Epidemiology of Overweight and Obesity among Psoriatic Patients and Control Group
2.2. The Influence of Genetic Variants on the BMI across Different Study Groups
2.2.1. Coding SNPs
2.2.2. Non-Coding SNPs
3. Discussion
4. Materials and Methods
4.1. Patient Group
4.2. Control Group
4.3. DNA Isolation
4.4. Microarrays Analysis
4.5. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, regional, and worldwide epidemiology of psoriasis: Systematic analysis and modelling study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Henseler, T.; Christophers, E. Psoriasis of early and late onset: Characterization of two types of psoriasis vulgaris. J. Am. Acad. Dermatol. 1985, 13, 450–456. [Google Scholar] [CrossRef]
- Ferrandiz, C.; Pujol, R.M.; Garcia-Patos, V.; Bordas, X.; Smandia, J.A. Psoriasis of early and late onset: A clinical and epidemiologic study from Spain. J. Am. Acad. Dermatol. 2002, 46, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.E.; Kassab, J.Y.; Rowland Payne, C.M.; Beer, W.E. Bimodality in age of onset of psoriasis, in both patients and their relatives. Dermatology 1993, 186, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Queiro, R.; Tejón, P.; Alonso, S.; Coto, P. Age at disease onset: A key factor for understanding psoriatic disease. Rheumatology 2014, 53, 1178–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samotij, D.; Nedoszytko, B.; Bartosińska, J.; Batycka-Baran, A.; Czajkowski, R.; Dobrucki, I.T.; Dobrucki, L.W.; Górecka-Sokołowska, M.; Janaszak-Jasienicka, A.; Krasowska, D.; et al. Pathogenesis of psoriasis in the “omic” era. Part I. Epidemiology, clinical manifestation, immunological and neuroendocrine disturbances. Postep. Dermatol. Alergol. 2020, 37, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Kisiel, B.; Kisiel, K.; Szymański, K.; Mackiewicz, W.; Biało-Wójcicka, E.; Uczniak, S.; Fogtman, A.; Iwanicka-Nowicka, R.; Koblowska, M.; Kossowska, H.; et al. The association between 38 previously reported polymorphisms and psoriasis in a Polish population: High predicative accuracy of a genetic risk score combining 16 loci. PLoS ONE 2017, 12, e0179348. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, K.; Okada, Y. The current landscape of psoriasis genetics in 2020. J. Dermatol. Sci. 2020, 99, 2–8. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Szczerkowska-Dobosz, A.; Stawczyk-Macieja, M.; Owczarczyk-Saczonek, A.; Reich, A.; Bartosińska, J.; Batycka- Baran, A.; Czajkowski, R.; Dobrucki, I.T.; Dobrucki, L.W.; et al. Pathogenesis of psoriasis in the “omic” era. Part II. Genetic, genomic and epigenetic changes in psoriasis. Postep. Dermatol. Alergol. 2020, 37, 283–298. [Google Scholar] [CrossRef]
- Ryan, C.; Kirby, B. Psoriasis is a systemic disease with multiple cardiovascular and metabolic comorbidities. Dermatol. Clin. 2015, 33, 41–55. [Google Scholar] [CrossRef]
- Owczarczyk-Saczonek, A.; Purzycka-Bohdan, D.; Nedoszytko, B.; Reich, A.; Szczerkowska-Dobosz, A.; Bartosiñska, J.; Batycka- Baran, A.; Czajkowski, R.; Dobrucki, I.T.; Dobrucki, L.W.; et al. Pathogenesis of psoriasis in the “omic” era. Part III. Metabolic disorders, metabolomics, nutrigenomics in psoriasis. Postep. Dermatol. Alergol. 2020, 37, 452–467. [Google Scholar] [CrossRef] [PubMed]
- Davidovici, B.B.; Sattar, N.; Prinz, J.; Puig, L.; Emery, P.; Barker, J.N.; van de Kerkhof, P.; Ståhle, M.; Nestle, F.O.; Girolomoni, G.; et al. Psoriasis and systemic inflammatory diseases: Potential mechanistic links between skin disease and co-morbid conditions. J. Investig. Dermatol. 2010, 130, 1785–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lønnberg, A.S.; Skov, L.; Skytthe, A.; Kyvik, K.O.; Pedersen, O.B.; Thomsen, S.F. Association of Psoriasis with the Risk for Type 2 Diabetes Mellitus and Obesity. JAMA Dermatol. 2016, 152, 761–767. [Google Scholar] [CrossRef]
- Rohde, K.; Keller, M.; la Cour Poulsen, L.; Blüher, M.; Kovacs, P.; Böttcher, Y. Genetics and epigenetics in obesity. Metabolism 2019, 92, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Abuabara, K.; Azfar, R.S.; Shin, D.B.; Neimann, A.L.; Troxel, A.B.; Gelfand, J.M. Cause-specific mortality in patients with severe psoriasis: A population-based cohort study in the UK. Br. J. Dermatol. 2010, 163, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Gisondi, P.; Del Giglio, M.; Girolomoni, G. Considerations for Systemic Treatment of Psoriasis in Obese Patients. Am. J. Clin. Dermatol. 2016, 17, 609–615. [Google Scholar] [CrossRef]
- Horreau, C.; Pouplard, C.; Brenaut, E.; Barnetche, T.; Misery, L.; Cribier, B.; Jullien, D.; Aractingi, S.; Aubin, F.; Joly, P.; et al. Cardiovascular morbidity and mortality in psoriasis and psoriatic arthritis: A systematic literature review. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, A.; Sanna, S.; Chen, W.M.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orrú, M.; Usala, G.; et al. Genome-wide association scan shows genetic variants in the FTO gene are associated with obesity-related traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef] [PubMed]
- Legry, V.; Cottel, D.; Ferrières, J.; Arveiler, D.; Andrieux, N.; Bingham, A.; Wagner, A.; Ruidavets, J.B.; Ducimetière, P.; Amouyel, P.; et al. Effect of an FTO polymorphism on fat mass, obesity, and type 2 diabetes mellitus in the French MONICA Study. Metabolism 2009, 58, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Liguori, R.; Labruna, G.; Alfieri, A.; Martone, D.; Farinaro, E.; Contaldo, F.; Sacchetti, L.; Pasanisi, F.; Buono, P. The FTO gene polymorphism (rs9939609) is associated with metabolic syndrome in morbidly obese subjects from southern Italy. Mol. Cell. Probes 2014, 28, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhu, Y.; Xu, F.; Ren, X.; Li, X.; Lai, M. FTO gene polymorphisms and obesity risk: A meta-analysis. BMC Med. 2011, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The International HapMap Consortium. The International HapMap Project. Nature 2003, 426, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coto-Segura, P.; González-Lara, L.; Eiris, N.; Santos-Juanes, J.; Queiro, R.; Coto, E. Effect of the FTO rs9930506 polymorphism on obesity and the main clinical outcomes in patients with psoriasis. Br. J. Dermatol. 2014, 171, 917–919. [Google Scholar] [CrossRef]
- Tupikowska-Marzec, M.; Kolačkov, K.; Zdrojowy-Wełna, A.; Słoka, N.K.; Szepietowski, J.C.; Maj, J. The Influence of FTO Polymorphism rs9939609 on Obesity, Some Clinical Features, and Disturbance of Carbohydrate Metabolism in Patients with Psoriasis. Biomed. Res. Int. 2019, 13, 7304345. [Google Scholar] [CrossRef] [Green Version]
- Ślęzak, R.; Leszczyński, P.; Warzecha, M.; Łaczmański, Ł.; Misiak, B. Assessment of the FTO gene polymorphisms in male patients with metabolic syndrome. Adv. Clin. Exp. Med. 2018, 27, 1581–1585. [Google Scholar] [CrossRef]
- Merkestein, M.; Dyan, S. Role of FTO in Adipocyte Development and Function: Recent Insights. Int. J. Endocrinol. 2015, 2015, 521381. [Google Scholar] [CrossRef]
- Tung, Y.C.L.; Yeo, G.S.H.; O’Rahilly, S.; Coll, A.P. Obesity and FTO: Changing Focus at a Complex Locus. Cell Metab. 2014, 20, 710–718. [Google Scholar] [CrossRef] [Green Version]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.P.; Els-Heindl, S.; Beck-Sickinger, A.G. Adrenomedullin—Current perspective on a peptide hormone with significant therapeutic potential. Peptides 2020, 131, 170347. [Google Scholar] [CrossRef]
- Harmancey, R.; Senard, J.M.; Rouet, P.; Pathak, A.; Smih, F. Adrenomedullin inhibits adipogenesis under transcriptional control of insulin. Diabetes 2007, 56, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jiang, C.; Wang, X.; Zhang, Y.; Shibahara, S.; Takahashi, K. Adrenomedullin is a novel adipokine: Adrenomedullin in adipocytes and adipose tissues. Peptides 2007, 28, 1129–1143. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, C.M.; Gomez-Llorente, C.; Tofe, I.; Gil-Campos, M.; Cañete, R.; Gil, Á. Genome-wide expression in visceral adipose tissue from obese prepubertal children. Int. J. Mol. Sci. 2015, 16, 7723–7737. [Google Scholar] [CrossRef] [PubMed]
- Baranova, A.; Collantes, R.; Gowder, S.J.; Elariny, H.; Schlauch, K.; Younoszai, A.; King, S.; Randhawa, M.; Pusulury, S.; Alsheddi, T.; et al. Obesity-related differential gene expression in the visceral adipose tissue. Obes. Surg. 2005, 15, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Gong, D.W.; Damcott, C.; Sabra, M.; Yang, R.; Pollin, T. Systematic analysis of omentin 1 and omentin 2 on 1q23 as candidate genes for type 2 diabetes in the Old Order Amish. Diabetes 2004, 53, A59. [Google Scholar]
- Kong, Y.; Zhang, S.; Wu, R.; Su, X.; Peng, D.; Zhao, M.; Su, Y. New insights into different adipokines in linking the pathophysiology of obesity and psoriasis. Lipids Health Dis. 2019, 18, 171. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhu, K.J.; Liu, J.L.; Xu, G.X.; Liu, W.; Jiang, F.X.; Zheng, H.F.; Quan, C. Omentin-1 plasma levels and omentin-1 expression are decreased in psoriatic lesions of psoriasis patients. Arch. Derm. Res. 2015, 307, 455–459. [Google Scholar] [CrossRef]
- Wolk, K.; Sabat, R. Adipokines in psoriasis: An important link between skin inflammation and metabolic alterations. Rev. Endocr. Metab. Disord. 2016, 17, 305–317. [Google Scholar] [CrossRef]
- Takahashi, H.; Tsuji, H.; Honma, M.; Ishida-Yamamoto, A.; Iizuka, H. Increased plasma resistin and decreased omentin levels in Japanese patients with psoriasis. Arch. Derm. Res. 2013, 305, 113–116. [Google Scholar] [CrossRef]
- Rizzo, H.E.; Escaname, E.N.; Alana, N.B.; Lavender, E.; Gelfond, J.; Fernandez, R.; Hibbs, M.A.; King, J.M.; Carr, N.R.; Blanco, C.L. Maternal diabetes and obesity influence the fetal epigenome in a largely Hispanic population. Clin. Epigenet. 2020, 12, 34. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, W.; Jiang, W.; Yao, J.; Zhang, D. An investigation of obesity susceptibility genes in Northern Han Chinese by targeted resequencing. Medicine 2017, 96, e6117. [Google Scholar] [CrossRef] [PubMed]
- Budu-Aggrey, A.; Brumpton, B.; Tyrrell, J.; Watkins, S.; Modalsli, E.H.; Celis-Morales, C.; Ferguson, L.D.; Vie, G.Å.; Palmer, T.; Fritsche, L.G.; et al. Evidence of a causal relationship between body mass index and psoriasis: A mendelian randomization study. PLoS Med. 2019, 16, e1002739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Sun, X.Y.; Miao, X.; Xu, R.; Ma, T.; Zhang, Y.N.; Li, H.J.; Li, B.; Li, X. Association between physical activity and risk of prevalent psoriasis: A MOOSE-compliant meta-analysis. Medicine 2018, 97, e11394. [Google Scholar] [CrossRef]
- Leino, M.; Mustonen, A.; Mattila, K.; Koulu, L.; Tuominen, R. Perceived impact of psoriasis on leisure-time activities. Eur. J. Dermatol. 2014, 24, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Auker, L.; Cordingley, L.; Pye, S.R.; Griffiths, C.E.M.; Young, H.S. What are the barriers to physical activity in patients with chronic plaque psoriasis? Br. J. Derm. 2020, 183, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Yeo, G.S.H. The genetics of obesity: From discovery to biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolska, S.; Michalska-Madej, J.; Słomka, M.; Sobalska-Kwapis, M.; Strapagiel, D. Biobank Łódź—population based biobank at the University of Łódź, Poland. Eur. J. Transl. Clin. Med. 2019, 1, 85–95. [Google Scholar] [CrossRef]
- Jarczak, J.; Grochowalski, Ł.; Marciniak, B.; Lach, J.; Słomka, M.; Sobalska-Kwapis, M.; Lorkiewicz, W.; Pułaski, Ł.; Strapagiel, D. Mitochondrial DNA variability of the Polish population. Eur. J. Hum. Genet. 2019, 27, 1304–1314. [Google Scholar] [CrossRef] [Green Version]
- Sobalska-Kwapis, M.; Suchanecka, A.; Słomka, M.; Siewierska-Górska, A.; Kępka, E.; Strapagiel, D. Genetic association of FTO/IRX region with obesity and overweight in the Polish population. PLoS ONE 2017, 12, e0180295. [Google Scholar] [CrossRef] [Green Version]
- Strapagiel, D.; Majewska, M.; Słomka, M.; Janik, K.; Sobalska, M.; Bartosz, G. Method for Determination of Sex. Polish Patent Pat 232653, 4 March 2019. [Google Scholar]



{kind=link}
{kind=link}
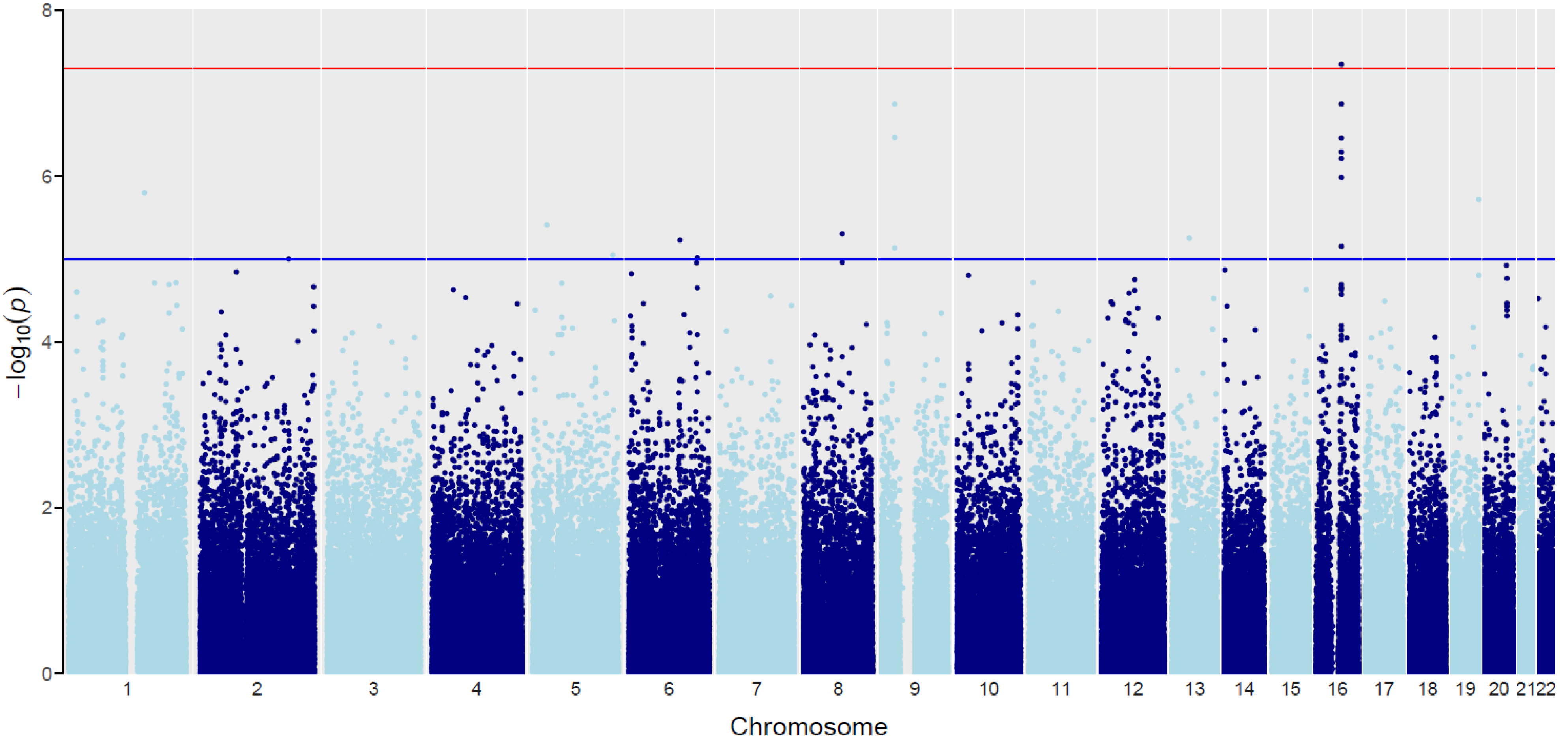
| SNP ID | Chr | Position | Gene-BMI Int: All | Gene-BMI Int: Type I | Gene-BMI Int: Type II | Gene-BMI Int Pval | Joint.Pval | Overlapped Gene | Gene Function |
|---|---|---|---|---|---|---|---|---|---|
| rs1558902 | 16 | 53803574 | 0.318627 | 0.873456933 | 0.29827713 | 0.00269419 | 4.47 × 10−8 | FTO | Protein coding |
| rs10968110 | 9 | 27792965 | 0.225899 | 1.07044773 | −0.645818448 | 5.95 × 10−5 | 1.35 × 10−7 | None | None |
| rs1556519 | 1 | 160917902 | −0.18039 | 1.477266915 | 1.211225346 | 3.73 × 10−7 | 1.58 × 10−6 | ITLN2 | Protein coding |
| rs12972098 | 19 | 58249304 | −0.40813 | 1.418581983 | 2.2106271 | 2.04 × 10−6 | 1.90 × 10−6 | AC003006.7 | Protein coding |
| rs4551082 | 5 | 29620270 | 0.134778 | 1.007787695 | 0.972212178 | 0.000176593 | 3.87 × 10−6 | None | None |
| rs12676670 | 8 | 81981261 | −0.03538 | 1.264637409 | 0.120877962 | 3.50 × 10−6 | 4.93 × 10−6 | PAG1 | Protein coding |
| rs4609724 | 13 | 55666639 | 0.308242 | 0.677809579 | −3.537824099 | 1.52 × 10−5 | 5.56 × 10−6 | None | None |
| rs9320269 | 6 | 109032557 | 0.320152 | −1.09262589 | −2.489035704 | 3.04 × 10−6 | 5.90 × 10−6 | None | None |
| rs2338833 | 5 | 169512063 | 0.0088 | 1.555408166 | 0.378795107 | 1.18 × 10−5 | 8.91 × 10−6 | None | None |
| rs1321529 | 6 | 145601560 | −0.34021 | 0.9503797 | −1.359245974 | 0.00015023 | 9.59 × 10−6 | None | None |
| rs696574 | 2 | 188228516 | 0.01579 | 1.031042583 | 2.283444931 | 1.53 × 10−5 | 9.96 × 10−6 | CALCRL | Protein coding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kisielnicka, A.; Sobalska-Kwapis, M.; Purzycka-Bohdan, D.; Nedoszytko, B.; Zabłotna, M.; Seweryn, M.; Strapagiel, D.; Nowicki, R.J.; Reich, A.; Samotij, D.; et al. The Analysis of a Genome-Wide Association Study (GWAS) of Overweight and Obesity in Psoriasis. Int. J. Mol. Sci. 2022, 23, 7396. https://doi.org/10.3390/ijms23137396
Kisielnicka A, Sobalska-Kwapis M, Purzycka-Bohdan D, Nedoszytko B, Zabłotna M, Seweryn M, Strapagiel D, Nowicki RJ, Reich A, Samotij D, et al. The Analysis of a Genome-Wide Association Study (GWAS) of Overweight and Obesity in Psoriasis. International Journal of Molecular Sciences. 2022; 23(13):7396. https://doi.org/10.3390/ijms23137396
Chicago/Turabian StyleKisielnicka, Anna, Marta Sobalska-Kwapis, Dorota Purzycka-Bohdan, Bogusław Nedoszytko, Monika Zabłotna, Michał Seweryn, Dominik Strapagiel, Roman J. Nowicki, Adam Reich, Dominik Samotij, and et al. 2022. "The Analysis of a Genome-Wide Association Study (GWAS) of Overweight and Obesity in Psoriasis" International Journal of Molecular Sciences 23, no. 13: 7396. https://doi.org/10.3390/ijms23137396
APA StyleKisielnicka, A., Sobalska-Kwapis, M., Purzycka-Bohdan, D., Nedoszytko, B., Zabłotna, M., Seweryn, M., Strapagiel, D., Nowicki, R. J., Reich, A., Samotij, D., Szczęch, J., Krasowska, D., Bartosińska, J., Narbutt, J., Lesiak, A., Barasińska, P., Owczarczyk-Saczonek, A., Czerwińska, J., Szepietowski, J. C., ... Szczerkowska-Dobosz, A. (2022). The Analysis of a Genome-Wide Association Study (GWAS) of Overweight and Obesity in Psoriasis. International Journal of Molecular Sciences, 23(13), 7396. https://doi.org/10.3390/ijms23137396