
The Contribution of the Minimal Promoter Element to the Activity of Synthetic Promoters Mediating CAR Expression in the Tumor Microenvironment


 and
and 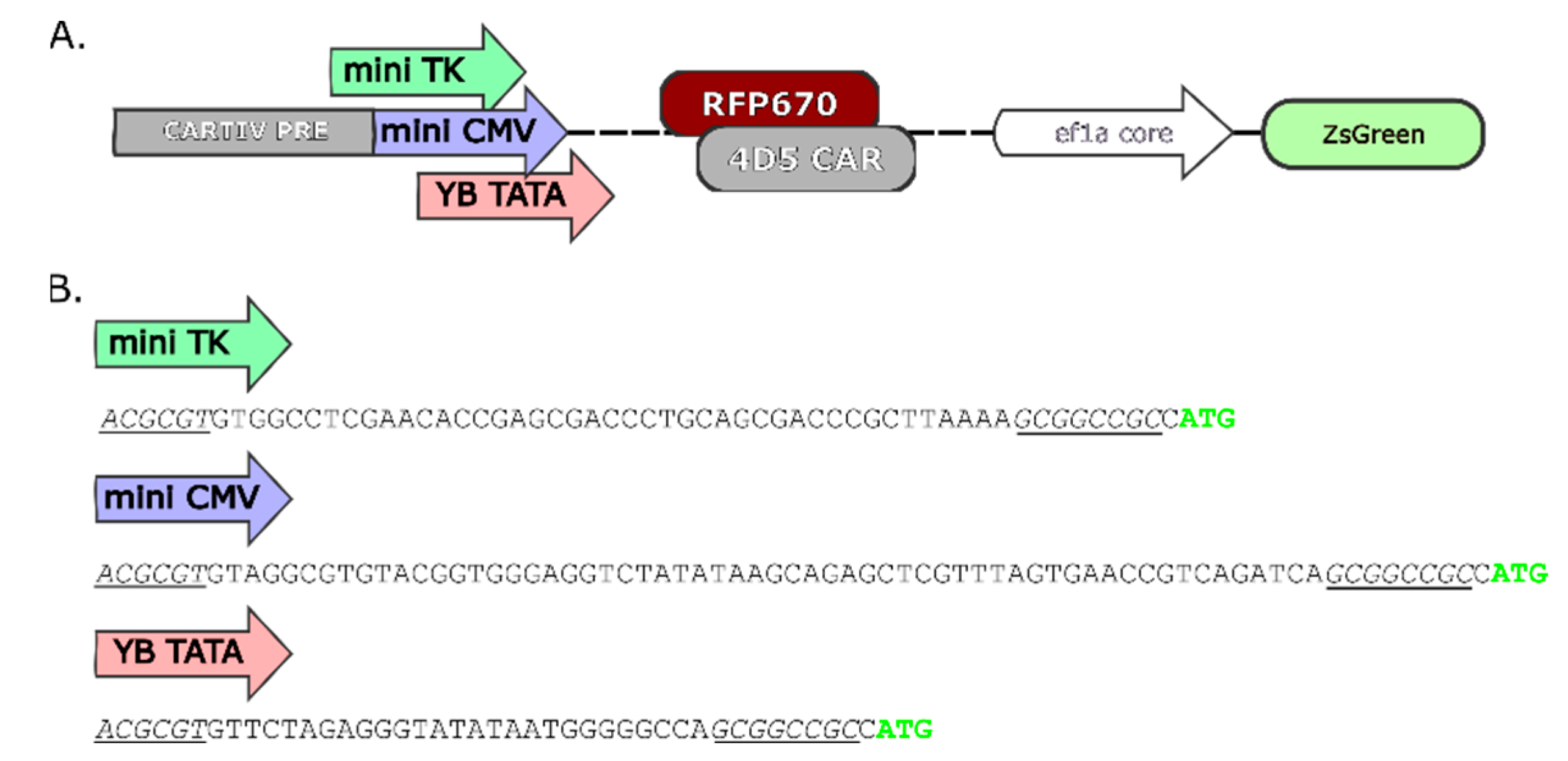{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Promoter Design and Constructs Used
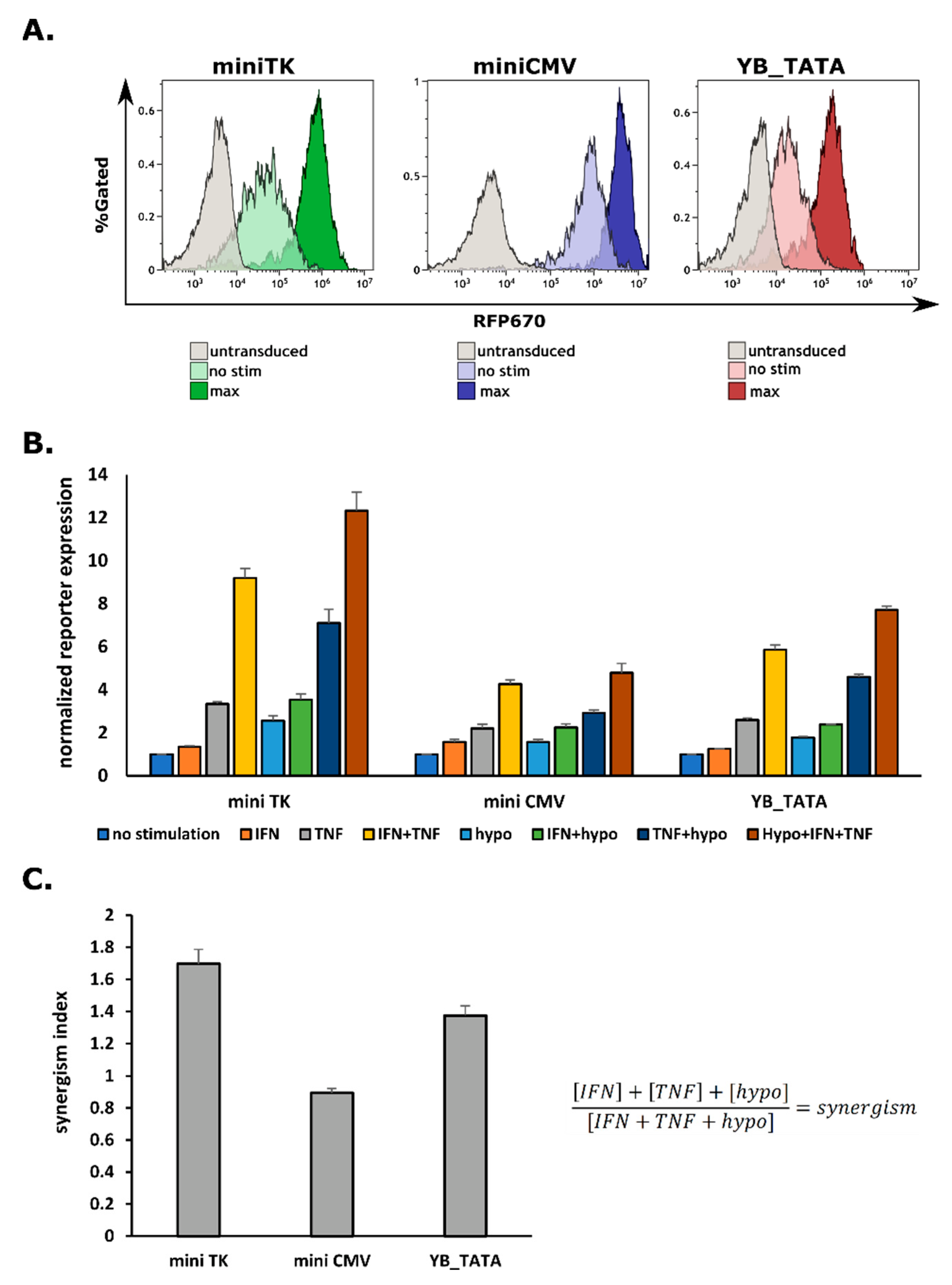
2.2. G1K0.6H1 PRE-Combination Is Functional with All Three Minimal Promoters
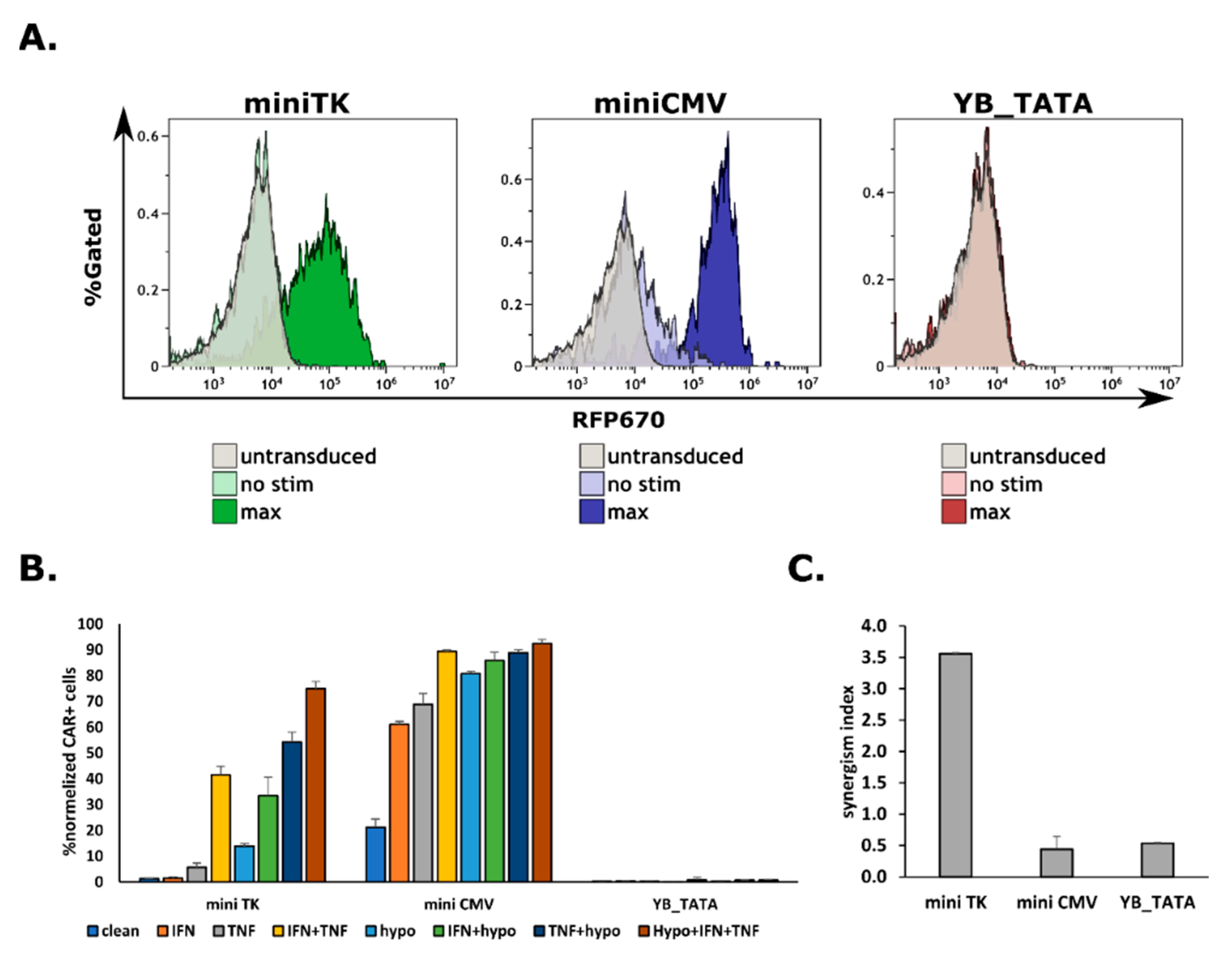
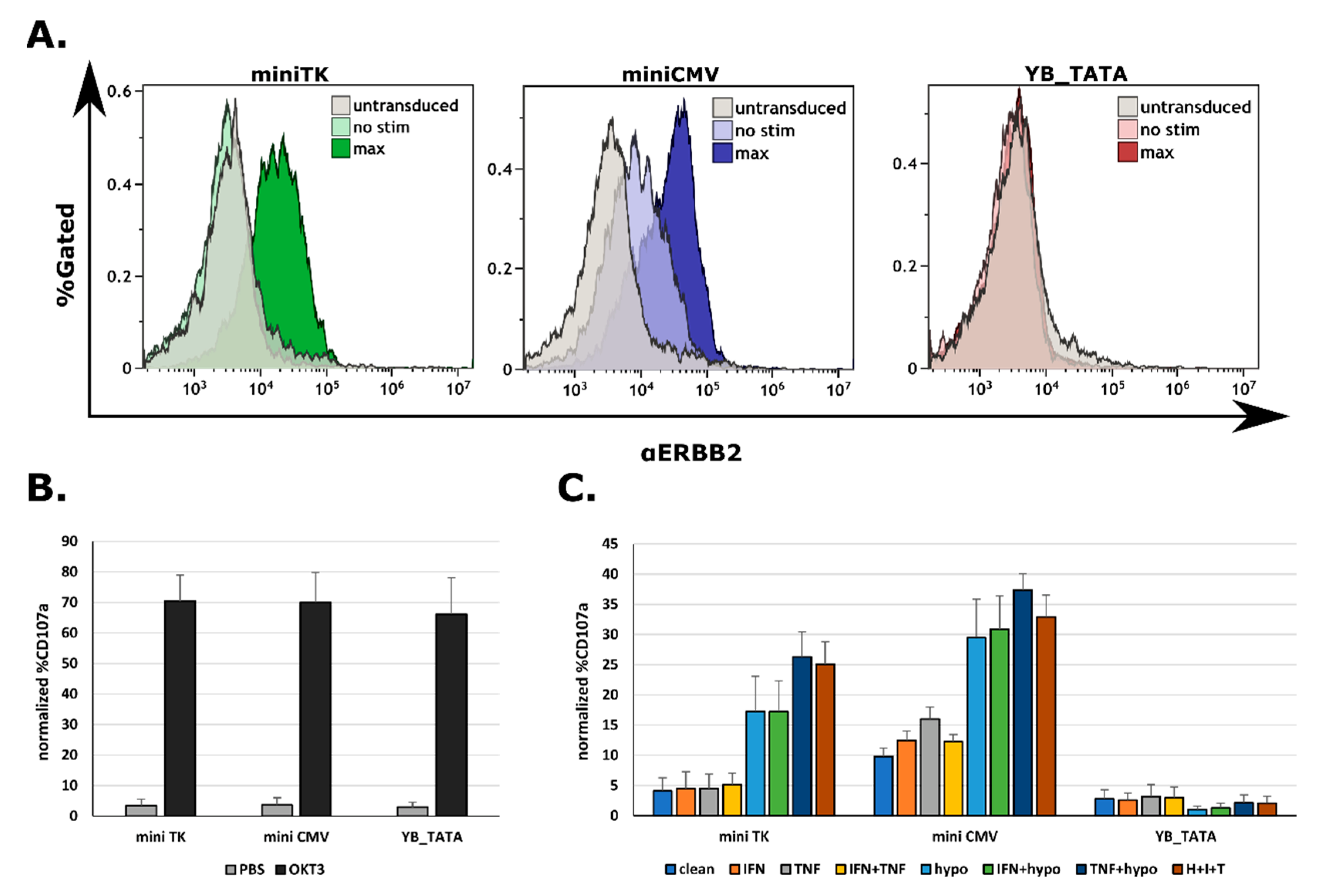
2.3. The Expression of a CAR on Cell’s Surface Is a Function of Promoter Strength
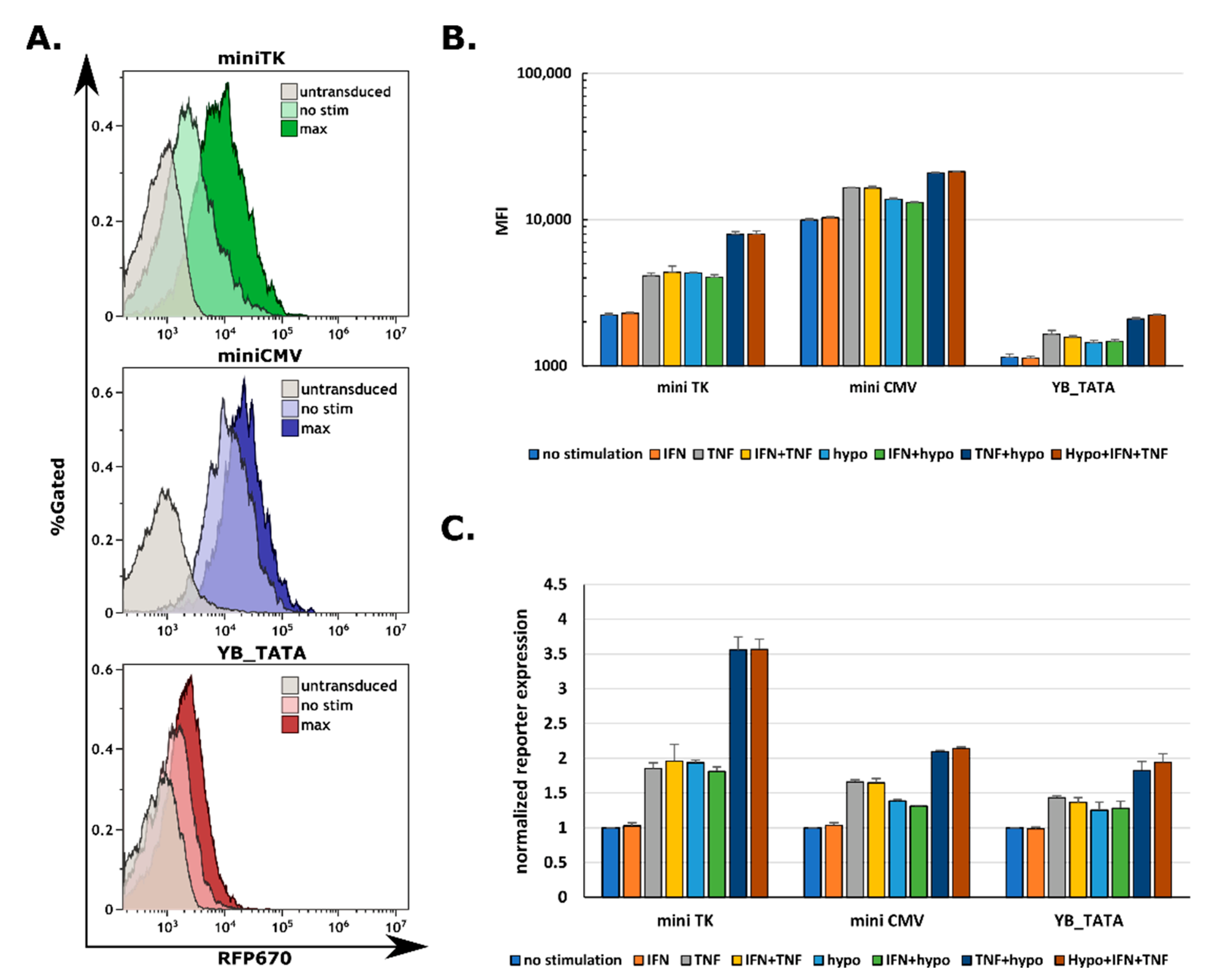
2.4. All Three Promoters Are Responsive to Combination Stimuli in Human Primary T Cells
2.5. CAR-T Response Could Be Fine-Tuned by the Promoter Driving the CAR Expression
3. Discussion
4. Materials and Methods
4.1. Cloning
4.2. Tissue Culture
4.3. Human Primary T Cells Culture and Infection
4.4. Lenti Virus Production and Concentration
4.5. Hypoxic Conditions
4.6. CARTIV Promoter Activity Assay
4.7. CAR Expression Assay
4.8. T Cell Functional Assay
4.9. Statistics and Reproducibility
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gross, G.; Eshhar, Z. Therapeutic Potential of T Cell Chimeric Antigen Receptors (CARs) in Cancer Treatment: Counteracting Off-Tumor Toxicities for Safe CAR T Cell Therapy. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Larson, R.C.; Maus, M. V Recent advances and discoveries in the mechanisms and functions of CAR T cells. Nat. Rev. Cancer 2021, 21, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Riddell, S.R. Engineering CAR-T cells: Design concepts. Trends Immunol. 2015, 36, 494–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, S.L.; Wang, W.W.; Toh, H.C. Cancer immunotherapy—The target is precisely on the cancer and also not. Ann. Acad. Med. Singap. 2018, 47, 381–387. [Google Scholar] [CrossRef]
- Holzinger, A.; Barden, M.; Abken, H. The growing world of CAR T cell trials: A systematic review. Cancer Immunol. Immunother. 2016, 65, 1433–1450. [Google Scholar] [CrossRef]
- D’Aloia, M.M.; Zizzari, I.G.; Sacchetti, B.; Pierelli, L.; Alimandi, M. CAR-T cells: The long and winding road to solid tumors. Cell Death Dis. 2018, 9, 282. [Google Scholar] [CrossRef]
- Grosser, R.; Cherkassky, L.; Chintala, N.; Adusumilli, P.S. Combination immunotherapy with CAR T cells and checkpoint blockade for the treatment of solid tumors. Cancer Cell 2019, 36, 471–482. [Google Scholar] [CrossRef]
- Ma, S.; Li, X.; Wang, X.; Cheng, L.; Li, Z.; Zhang, C.; Ye, Z.; Qian, Q. Current progress in CAR-T cell therapy for solid tumors. Int. J. Biol. Sci. 2019, 15, 2548. [Google Scholar] [CrossRef] [Green Version]
- Di Stasi, A.; Tey, S.K.; Dotti, G.; Fujita, Y.; Kennedy-Nasser, A.; Martinez, C.; Straathof, K.; Liu, E.; Durett, A.G.; Grilley, B.; et al. Inducible apoptosis as a safety switch for adoptive cell therapy. N. Engl. J. Med. 2011, 365, 1673–1683. [Google Scholar] [CrossRef] [Green Version]
- Diaconu, I.; Ballard, B.; Zhang, M.; Chen, Y.; West, J.; Dotti, G.; Savoldo, B. Inducible Caspase-9 Selectively Modulates the Toxicities of CD19-Specific Chimeric Antigen Receptor-Modified T Cells. Mol. Ther. 2017, 25, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Gargett, T.; Brown, M.P. The inducible caspase-9 suicide gene system as a “safety switch” to limit on-target, off-tumor toxicities of chimeric antigen receptor T-cells. Front. Pharmacol. 2014, 5, 235. [Google Scholar] [CrossRef]
- Khanali, J.; Azangou-Khyavy, M.; Boroomand-Saboor, M.; Ghasemi, M.; Niknejad, H. JAK/STAT-Dependent Chimeric Antigen Receptor (CAR) Expression: A Design Benefiting From a Dual AND/OR Gate Aiming to Increase Specificity, Reduce Tumor Escape and Affect Tumor Microenvironment. Front. Immunol. 2021, 12, 2181. [Google Scholar] [CrossRef]
- Fedorov, V.D.; Themeli, M.; Sadelain, M. PD-1- and CTLA-4-based inhibitory chimeric antigen receptors (iCARs) divert off-target immunotherapy responses. Sci. Transl. Med. 2013, 5, 215ra172. [Google Scholar] [CrossRef] [Green Version]
- Sharabi, O.; Greenshpan, Y.; Ofir, N.; Ottolenghi, A.; Levi, T.; Olender, L.; Adler-Agmon, Z.; Porgador, A.; Gazit, R. High throughput screen for the improvement of inducible promoters for tumor microenvironment cues. Sci. Rep. 2022, 12, 7169. [Google Scholar] [CrossRef]
- Greenshpan, Y.; Sharabi, O.; Ottolenghi, A.; Cahana, A.; Kundu, K.; Yegodayev, K.M.; Elkabets, M.; Gazit, R.; Porgador, A. Synthetic promoters to induce immune-effectors into the tumor microenvironment. Commun. Biol. 2021, 4, 143. [Google Scholar] [CrossRef]
- Hawkins, E.R.; D’Souza, R.R.; Klampatsa, A. Armored CAR T-cells: The next chapter in T-cell cancer immunotherapy. Biol. Targets Ther. 2021, 15, 95. [Google Scholar] [CrossRef]
- Narayan, V.; Barber-Rotenberg, J.S.; Jung, I.-Y.; Lacey, S.F.; Rech, A.J.; Davis, M.M.; Hwang, W.-T.; Lal, P.; Carpenter, E.L.; Maude, S.L. PSMA-targeting TGFβ-insensitive armored CAR T cells in metastatic castration-resistant prostate cancer: A phase 1 trial. Nat. Med. 2022, 28, 724–734. [Google Scholar] [CrossRef]
- Arcot, S.S.; Flemington, E.K.; Deininger, P.L. The human thymidine kinase gene promoter: Deletion analysis and specific protein binding. J. Biol. Chem. 1989, 264, 2343–2349. [Google Scholar] [CrossRef]
- Jeang, K.T.; Cho, M.S.; Hayward, G.S. Abundant constitutive expression of the immediate-early 94K protein from cytomegalovirus (Colburn) in a DNA-transfected mouse cell line. Mol. Cell. Biol. 1984, 4, 2214–2223. [Google Scholar]
- Byrne, B.J.; Davis, M.S.; Yamaguchi, J.; Bergsma, D.J.; Subramanian, K.N. Definition of the simian virus 40 early promoter region and demonstration of a host range bias in the enhancement effect of the simian virus 40 72-base-pair repeat. Proc. Natl. Acad. Sci. USA 1983, 80, 721–725. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.; Mailand, E.; Swaminathan, K.K.; Schreiber, J.; Angelici, B.; Benenson, Y. Transplantation of prokaryotic two-component signaling pathways into mammalian cells. Proc. Natl. Acad. Sci. USA 2014, 111, 15705–15710. [Google Scholar] [CrossRef] [Green Version]
- Ede, C.; Chen, X.; Lin, M.-Y.; Chen, Y.Y. Quantitative analyses of core promoters enable precise engineering of regulated gene expression in mammalian cells. ACS Synth. Biol. 2016, 5, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.; Morris, K.V.; Shevchenko, G. Hypoxia-directed tumor targeting of CRISPR-Cas9 and HSV-TK suicide gene therapy using lipid nanoparticles. Mol. Ther. Clin. Dev. 2022, 25, 158–169. [Google Scholar] [CrossRef]
- Fillat, C.; Carrio, M.; Cascante, A.; Sangro, B. Suicide gene therapy mediated by the Herpes Simplex virus thymidine kinase gene/Ganciclovir system: Fifteen years of application. Curr. Gene Ther. 2003, 3, 13–26. [Google Scholar] [CrossRef]
- Gossen, M.; Bujard, H. Tight control of gene expression in mammalian cells by tetracycline-responsive promoters. Proc. Natl. Acad. Sci. USA 1992, 89, 5547–5551. [Google Scholar] [CrossRef] [Green Version]
- Gitzinger, M.; Kemmer, C.; El-Baba, M.D.; Weber, W.; Fussenegger, M. Controlling transgene expression in subcutaneous implants using a skin lotion containing the apple metabolite phloretin. Proc. Natl. Acad. Sci. USA 2009, 106, 10638–10643. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jiang, Y.; Chen, H.; Liao, W.; Li, Z.; Weiss, R.; Xie, Z. Modular construction of mammalian gene circuits using TALE transcriptional repressors. Nat. Chem. Biol. 2015, 11, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, Q.J.; Yang, S.; Kochenderfer, J.N.; Zheng, Z.; Zhong, X.; Sadelain, M.; Eshhar, Z.; Rosenberg, S.A.; Morgan, R.A. A Herceptin-Based Chimeric Antigen Receptor with Modified Signaling Domains Leads to Enhanced Survival of Transduced T Lymphocytes and Antitumor Activity. J. Immunol. 2009, 183, 5563–5574. [Google Scholar] [CrossRef]
- Hu, C.-J.; Wang, L.-Y.; Chodosh, L.A.; Keith, B.; Simon, M.C. Differential Roles of Hypoxia-Inducible Factor 1 (HIF-1) and HIF-2 in Hypoxic Gene Regulation. Mol. Cell. Biol. 2003, 9361–9374. [Google Scholar] [CrossRef] [Green Version]
- Land, S.C.; Tee, A.R. Hypoxia-inducible factor 1α is regulated by the mammalian target of rapamycin (mTOR) via an mTOR signaling motif. J. Biol. Chem. 2007, 282, 20534–20543. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Marquez, P.; Calleja-Cervantes, M.E.; Serrano, G.; Palacios-Berraquero, M.L.; Alignani, D.; San Martin-Uriz, P.; Vilas-Zornoza, A.; Ceballos, C.; Martin-Mallo, A.; Rodríguez-Diaz, S. CAR Density Influences Antitumoral Efficacy of BCMA CAR-T Cells and Correlates with Clinical Outcome. Blood 2021, 138, 735. [Google Scholar] [CrossRef]
- Chee, J.Y.; Chin, C.F. Gateway cloning technology: Advantages and drawbacks. Cloning Transgenes 2015, 4, 138. [Google Scholar]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Lundh, S.; Kim, D.; Woodhouse, S.; Barrett, D.M.; Myers, R.M.; Grupp, S.A.; Maus, M.V.; June, C.H.; Camara, P.G. Single-cell multiomics dissection of basal and antigen-specific activation states of CD19-targeted CAR T cells. J. Immunother. Cancer 2021, 9, e002328. [Google Scholar] [CrossRef] [PubMed]
- Viaud, S.; Ma, J.S.Y.; Hardy, I.R.; Hampton, E.N.; Benish, B.; Sherwood, L.; Nunez, V.; Ackerman, C.J.; Khialeeva, E.; Weglarz, M. Switchable control over in vivo CAR T expansion, B cell depletion, and induction of memory. Proc. Natl. Acad. Sci. USA 2018, 115, E10898–E10906. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, K.; Tsunei, A.; Kusabuka, H.; Ogaki, E.; Tachibana, M.; Okada, N. Hinge and transmembrane domains of chimeric antigen receptor regulate receptor expression and signaling threshold. Cells 2020, 9, 1182. [Google Scholar] [CrossRef]
- Hirobe, S.; Imaeda, K.; Tachibana, M.; Okada, N. The Effects of Chimeric Antigen Receptor (CAR) Hinge Domain Post-Translational Modifications on CAR-T Cell Activity. Int. J. Mol. Sci. 2022, 23, 4056. [Google Scholar] [CrossRef]
- Lazar-Molnar, E.; Almo, S.C.; Nathenson, S.G. The interchain disulfide linkage is not a prerequisite but enhances CD28 costimulatory function. Cell. Immunol. 2006, 244, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.Y.; Mikolajczak, S.A.; Yoshida, T.; Yoshida, R.; Kelvin, D.J.; Ochi, A. CD28 T cell costimulatory receptor function is negatively regulated by N-linked carbohydrates. Biochem. Biophys. Res. Commun. 2004, 317, 60–67. [Google Scholar] [CrossRef]
- Shore, D.A.; Wilson, I.A.; Dwek, R.A.; Rudd, P.M. Glycosylation and the function of the T cell co-receptor CD8. In Glycobiology and Medicine; Springer: Berlin, Germany, 2005; pp. 71–84. [Google Scholar]
- June, C.H.; O’Connor, R.S.; Kawalekar, O.U.; Ghassemi, S.; Milone, M.C. CAR T cell immunotherapy for human cancer. Science 2018, 359, 1361–1365. [Google Scholar] [CrossRef] [Green Version]
- Feins, S.; Kong, W.; Williams, E.F.; Milone, M.C.; Fraietta, J.A. An introduction to chimeric antigen receptor (CAR) T-cell immunotherapy for human cancer. Am. J. Hematol. 2019, 94, S3–S9. [Google Scholar] [CrossRef] [Green Version]
- Pauza, C.D.; Liou, M.-L.; Lahusen, T.; Xiao, L.; Lapidus, R.G.; Cairo, C.; Li, H. Gamma delta T cell therapy for cancer: It is good to be local. Front. Immunol. 2018, 9, 1305. [Google Scholar] [CrossRef]
- Capsomidis, A.; Benthall, G.; Van Acker, H.H.; Fisher, J.; Kramer, A.M.; Abeln, Z.; Majani, Y.; Gileadi, T.; Wallace, R.; Gustafsson, K. Chimeric antigen receptor-engineered human gamma delta T cells: Enhanced cytotoxicity with retention of cross presentation. Mol. Ther. 2018, 26, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Prasad, V. Tisagenlecleucel—the first approved CAR-T-cell therapy: Implications for payers and policy makers. Nat. Rev. Clin. Oncol. 2018, 15, 11–12. [Google Scholar] [CrossRef]
- Majzner, R.G.; Rietberg, S.P.; Sotillo, E.; Dong, R.; Vachharajani, V.T.; Labanieh, L.; Myklebust, J.H.; Kadapakkam, M.; Weber, E.W.; Tousley, A.M. Tuning the antigen density requirement for CAR T-cell activity. Cancer Discov. 2020, 10, 702–723. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Kuramitsu, S.; Posey, A.D., Jr.; June, C.H. Expanding the therapeutic window for CAR T cell therapy in solid tumors: The knowns and unknowns of CAR T cell biology. Front. Immunol. 2018, 9, 2486. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greenshpan, Y.; Sharabi, O.; Yegodayev, K.M.; Novoplansky, O.; Elkabets, M.; Gazit, R.; Porgador, A. The Contribution of the Minimal Promoter Element to the Activity of Synthetic Promoters Mediating CAR Expression in the Tumor Microenvironment. Int. J. Mol. Sci. 2022, 23, 7431. https://doi.org/10.3390/ijms23137431
Greenshpan Y, Sharabi O, Yegodayev KM, Novoplansky O, Elkabets M, Gazit R, Porgador A. The Contribution of the Minimal Promoter Element to the Activity of Synthetic Promoters Mediating CAR Expression in the Tumor Microenvironment. International Journal of Molecular Sciences. 2022; 23(13):7431. https://doi.org/10.3390/ijms23137431
Chicago/Turabian StyleGreenshpan, Yariv, Omri Sharabi, Ksenia M. Yegodayev, Ofra Novoplansky, Moshe Elkabets, Roi Gazit, and Angel Porgador. 2022. "The Contribution of the Minimal Promoter Element to the Activity of Synthetic Promoters Mediating CAR Expression in the Tumor Microenvironment" International Journal of Molecular Sciences 23, no. 13: 7431. https://doi.org/10.3390/ijms23137431
APA StyleGreenshpan, Y., Sharabi, O., Yegodayev, K. M., Novoplansky, O., Elkabets, M., Gazit, R., & Porgador, A. (2022). The Contribution of the Minimal Promoter Element to the Activity of Synthetic Promoters Mediating CAR Expression in the Tumor Microenvironment. International Journal of Molecular Sciences, 23(13), 7431. https://doi.org/10.3390/ijms23137431