
PEDF Deletion Induces Senescence and Defects in Phagocytosis in the RPE



Abstract
:1. Introduction
2. Results
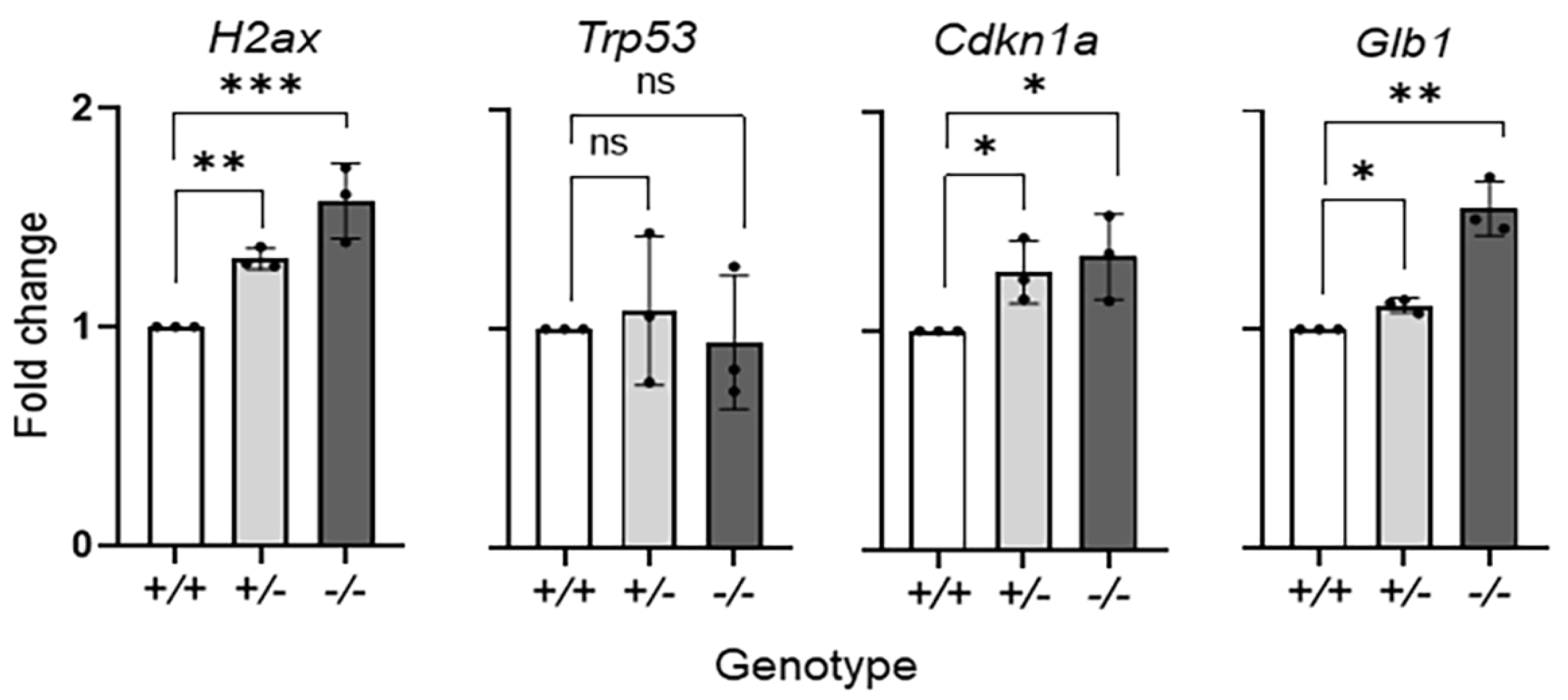
2.1. Serpinf1 Deletion Increased Senescence-Associated Gene Expression
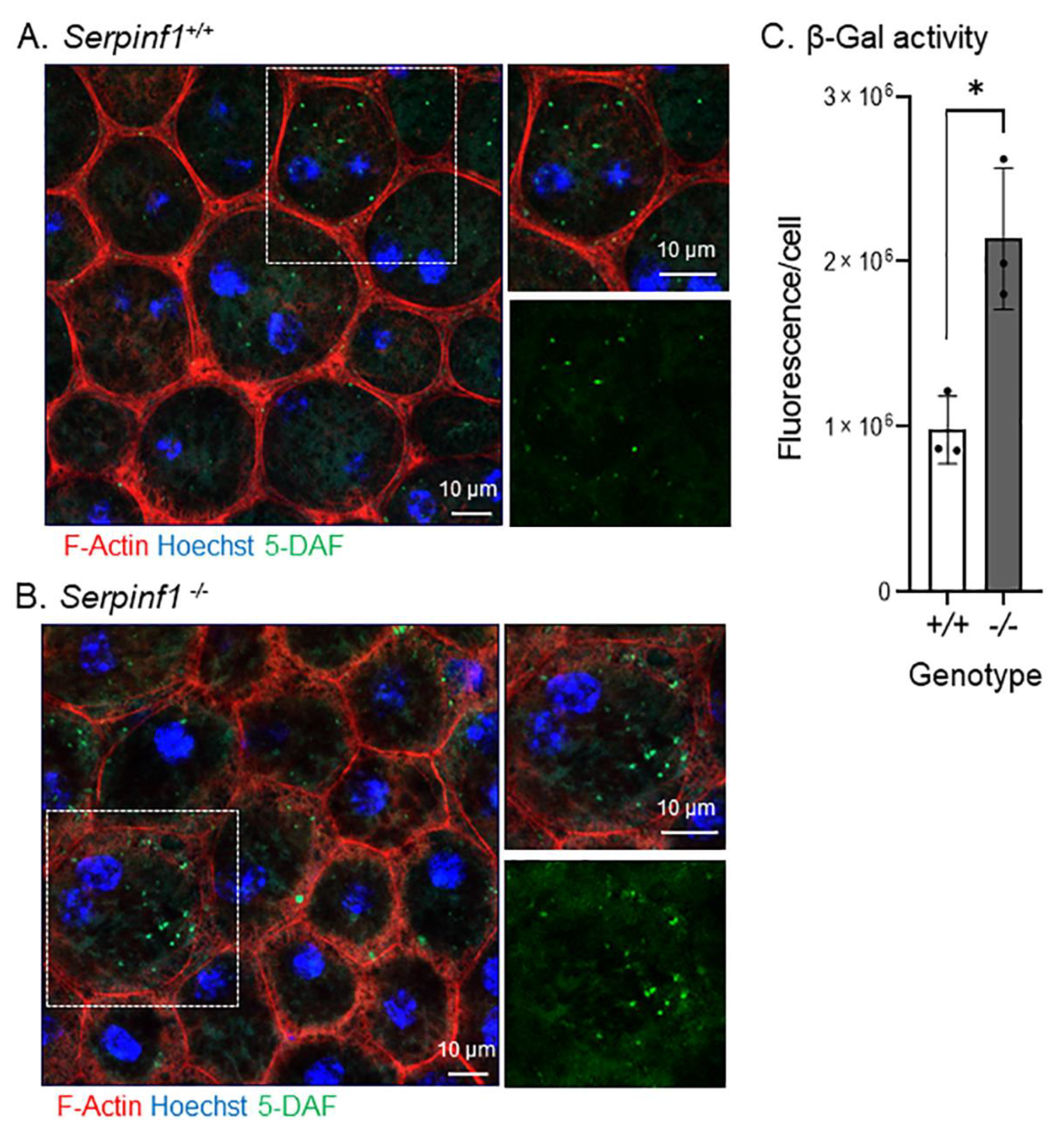
2.2. Serpinf1 Deletion Increased Β-Galactosidase Activity in the RPE
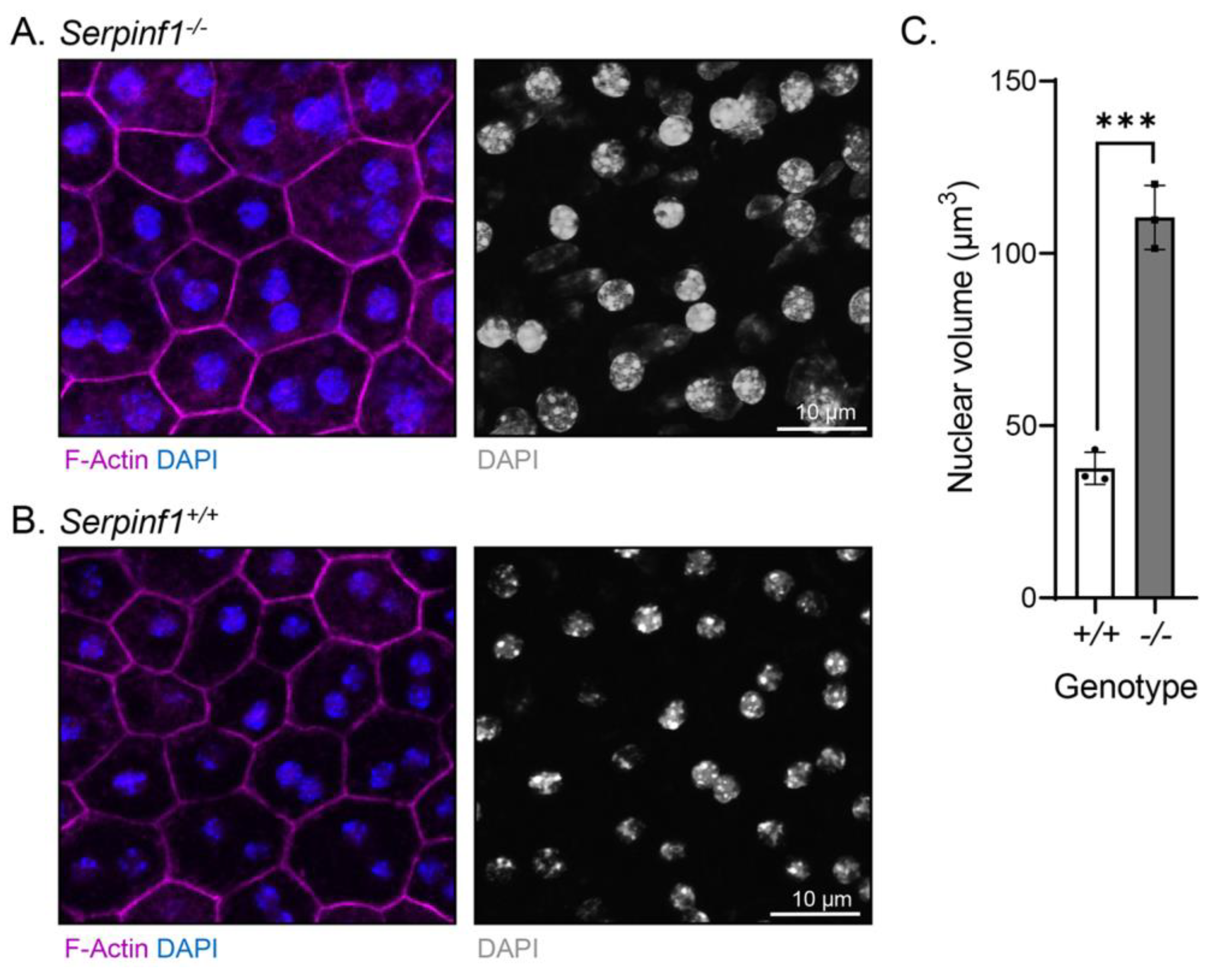
2.3. Serpinf1 Deletion Increased the Size of Nuclei of the RPE Cells
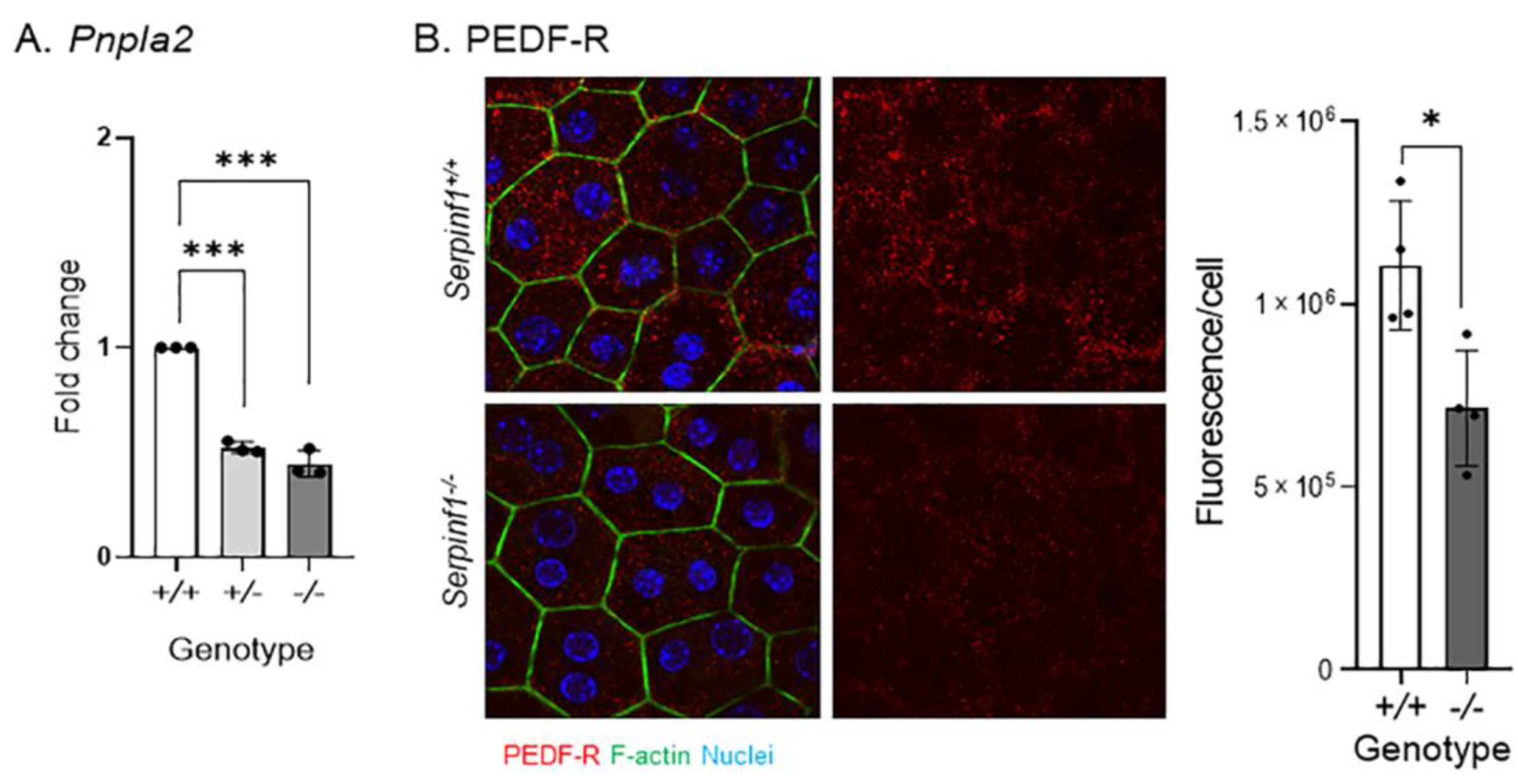
2.4. Serpinf1 Ablation Downregulated Pnpla2 in the RPE
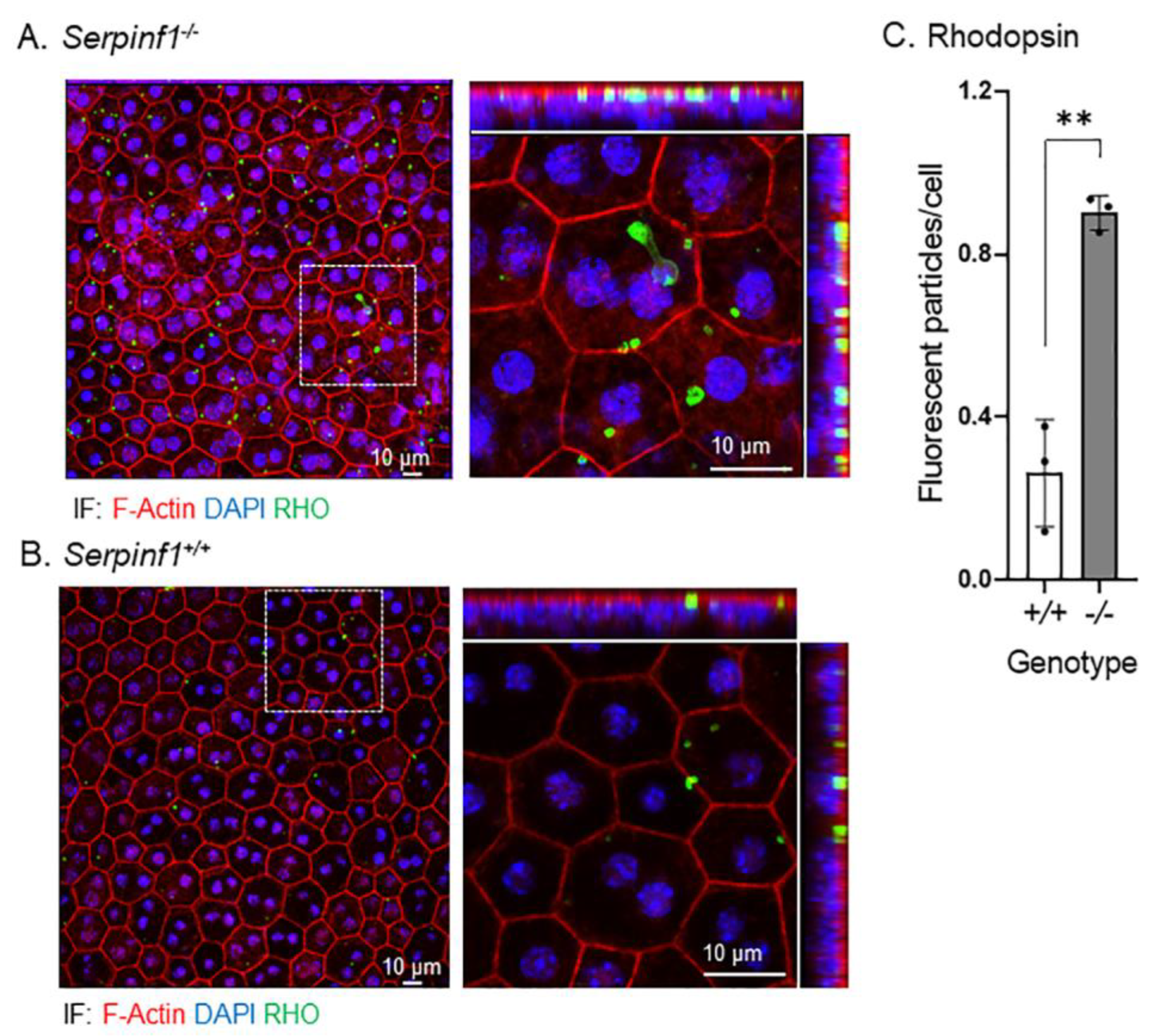
2.5. Rhodopsin and Lipid Accumulated in the Serpinf1-Depleted RPE
3. Discussion
4. Materials and Methods
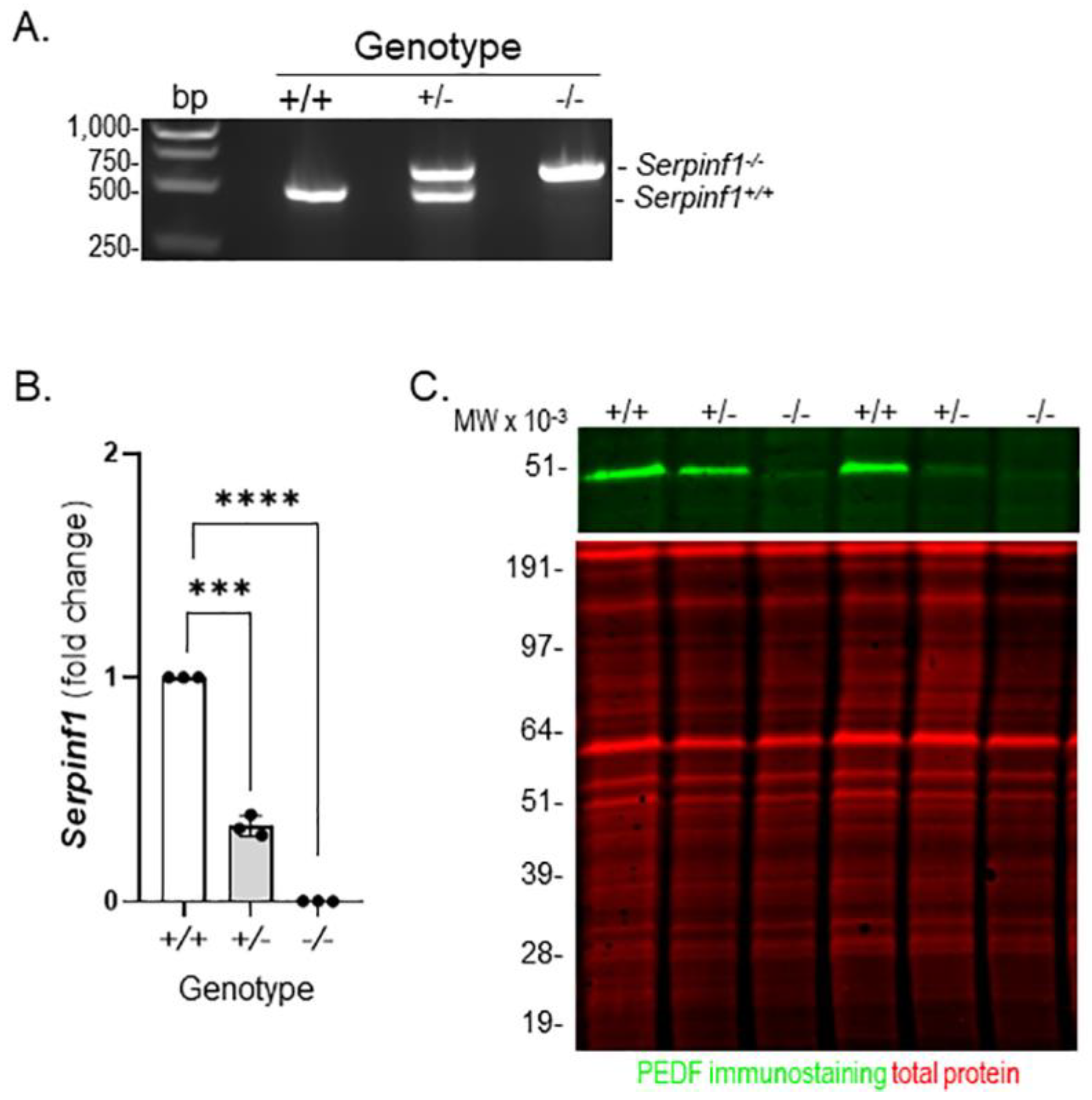
4.1. Animals
4.2. RPE/Choroid Flat Mounts
4.3. Immunofluorescence and Fluorescent Confocal Microscopy
4.4. Fluorescent Imaging Quantification
4.5. Transcript Levels Determination by Quantitative PCR (qPCR)
4.6. Western Blot
4.7. β-Galactosidase Activity Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becerra, S.P.; Fariss, R.N.; Wu, Y.Q.; Montuenga, L.M.; Wong, P.; Pfeffer, B.A. Pigment epithelium-derived factor in the monkey retinal pigment epithelium and interphotoreceptor matrix: Apical secretion and distribution. Exp. Eye Res. 2004, 78, 223–234. [Google Scholar] [CrossRef]
- Zhu, D.; Deng, X.; Spee, C.; Sonoda, S.; Hsieh, C.-L.; Barron, E.; Pera, M.; Hinton, D.R. Polarized Secretion of PEDF from Human Embryonic Stem Cell–Derived RPE Promotes Retinal Progenitor Cell Survival. Investig. Opthalmol. Vis. Sci. 2011, 52, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Liu, J.; Pu, J.; Milne, G.; Chen, M.; Xu, H.; Shipley, A.; Forrester, J.V.; McCaig, C.D.; Lois, N. Polarized retinal pigment epithelium generates electrical signals that diminish with age and regulate retinal pathology. J. Cell. Mol. Med. 2018, 22, 5552–5564. [Google Scholar] [CrossRef]
- Van Kirk, C.A.; VanGuilder, H.D.; Young, M.; Farley, J.A.; Sonntag, W.E.; Freeman, W.M. Age-related alterations in retinal neurovascular and inflammatory transcripts. Mol. Vis. 2011, 17, 1261–1274. [Google Scholar]
- Kolomeyer, A.M.; Sugino, I.K.; Zarbin, M.A. Characterization of conditioned media collected from cultured adult versus fetal retinal pigment epithelial cells. Investig. Opthalmol. Vis. Sci. 2011, 52, 5973–5986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinle, J.J.; Sharma, S.; Chin, V.C. Normal aging involves altered expression of growth factors in the rat choroid. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Subramanian, P.; Shen, D.; Tuo, J.; Becerra, S.P.; Chan, C.C. Pigment epithelium-derived factor reduces apoptosis and pro-inflammatory cytokine gene expression in a murine model of focal retinal degeneration. ASN Neuro 2013, 5, e00126. [Google Scholar] [CrossRef] [Green Version]
- Francis, M.K.; Appel, S.; Meyer, C.; Balin, S.J.; Balin, A.K.; Cristofalo, V.J. Loss of EPC-1/PEDF expression during skin aging in vivo. J. Investig. Dermatol. 2004, 122, 1096–1105. [Google Scholar] [CrossRef] [Green Version]
- Pignolo, R.J.; Cristofalo, V.J.; Rotenberg, M.O. Senescent WI-38 cells fail to express EPC-1, a gene induced in young cells upon entry into the G0 state. J. Biol. Chem. 1993, 268, 8949–8957. [Google Scholar] [CrossRef]
- Steele, F.R.; Chader, G.J.; Johnson, L.V.; Tombran-Tink, J. Pigment epithelium-derived factor: Neurotrophic activity and identification as a member of the serine protease inhibitor gene family. Proc. Natl. Acad. Sci. USA 1993, 90, 1526–1530. [Google Scholar] [CrossRef] [Green Version]
- Polato, F.; Becerra, S.P. Pigment Epithelium-Derived Factor, a Protective Factor for Photoreceptors in Vivo. Adv. Exp. Med. Biol. 2016, 854, 699–706. [Google Scholar] [PubMed]
- Mori, K.; Duh, E.; Gehlbach, P.; Ando, A.; Takahashi, K.; Pearlman, J.; Mori, K.; Yang, H.S.; Zack, D.; Ettyreddy, D.; et al. Pigment epithelium-derived factor inhibits retinal and choroidal neovascularization. J. Cell. Physiol. 2001, 188, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Dawson, D.W.; Volpert, O.V.; Gillis, P.; Crawford, S.E.; Xu, H.-J.; Benedict, W.; Bouck, N.P. Pigment epithelium-derived factor: A potent inhibitor of angiogenesis. Science 1999, 285, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Bouck, N. PEDF: Anti-angiogenic guardian of ocular function. Trends Mol. Med. 2002, 8, 330–334. [Google Scholar] [CrossRef]
- Craword, S.E.; Fitchev, P.; Veliceasa, D.; Volpert, O.V. The many facets of PEDF in drug discovery and disease: A diamond in the rough or split personality disorder? Expert Opin. Drug Discov. 2013, 8, 769–792. [Google Scholar] [CrossRef]
- Dixit, S.; Polato, F.; Samardzija, M.; Abu-Asab, M.; Grimm, C.; Crawford, S.E.; Becerra, S.P. PEDF deficiency increases the susceptibility of rd10 mice to retinal degeneration. Exp. Eye Res. 2020, 198, 108121. [Google Scholar] [CrossRef]
- Subramanian, P.; Locatelli-Hoops, S.; Kenealey, J.; DesJardin, J.; Notari, L.; Becerra, S.P. Pigment epithelium-derived factor (PEDF) prevents retinal cell death via PEDF Receptor (PEDF-R): Identification of a functional ligand binding site. J. Biol. Chem. 2013, 288, 23928–23942. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, A.S.; Duffy, J.; Wang, M.C. Lipid metabolism and lipid signals in aging and longevity. Dev. Cell 2021, 56, 1394–1407. [Google Scholar] [CrossRef]
- Johnson, A.A.; Stolzing, A. The role of lipid metabolism in aging, lifespan regulation, and age-related disease. Aging Cell 2019, 18, e13048. [Google Scholar] [CrossRef] [Green Version]
- Flor, A.C.; Wolfgeher, D.; Wu, D.; Kron, S.J. A signature of enhanced lipid metabolism, lipid peroxidation and aldehyde stress in therapy-induced senescence. Cell Death Discov. 2017, 3, 17075. [Google Scholar] [CrossRef] [Green Version]
- Griveau, A.; Wiel, C.; Le Calvé, B.; Ziegler, D.; Djebali, S.; Warnier, M.; Martin, N.; Marvel, J.; Vindrieux, D.; Bergo, M.O.; et al. Targeting the phospholipase A2 receptor ameliorates premature aging phenotypes. Aging Cell 2018, 17, e12835. [Google Scholar] [CrossRef] [PubMed]
- Jové, M.; Naudí, A.; Aledo, J.C.; Cabré, R.; Ayala, V.; Portero-Otin, M.; Barja, G.; Pamplona, R. Plasma long-chain free fatty acids predict mammalian longevity. Sci. Rep. 2013, 3, 3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobson, R.W.; Nabholz, B.; Galtier, N. An evolutionary genome scan for longevity-related natural selection in mammals. Mol. Biol. Evol. 2009, 27, 840–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, D.; Blier, P.U. The extreme longevity of Arctica islandica is associated with increased peroxidation resistance in mitochondrial membranes. Aging Cell 2012, 11, 845–855. [Google Scholar] [CrossRef]
- Borchman, D.; Stimmelmayr, R.; George, J.C. Whales, lifespan, phospholipids, and cataracts. J. Lipid Res. 2017, 58, 2289–2298. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, T.W.; Buffenstein, R.; Hulbert, A. Membrane phospholipid composition may contribute to exceptional longevity of the naked mole-rat (Heterocephalus glaber): A comparative study using shotgun lipidomics. Exp. Gerontol. 2007, 42, 1053–1062. [Google Scholar] [CrossRef]
- Huang, X.; Withers, B.R.; Dickson, R.C. Sphingolipids and lifespan regulation. Biochim. Biophys. Acta 2014, 1841, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Webster, M.; Witkin, K.L.; Cohen-Fix, O. Sizing up the nucleus: Nuclear shape, size and nuclear-envelope assembly. J. Cell Sci. 2009, 122, 1477–1486. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Fix, O. Cell biology: How does the nucleus get its membrane? Curr. Biol. 2021, 31, R1077–R1079. [Google Scholar] [CrossRef]
- Inana, G.; Murat, C.; An, W.; Yao, X.; Harris, I.R.; Cao, J. RPE phagocytic function declines in age-related macular degeneration and is rescued by human umbilical tissue derived cells. J. Transl. Med. 2018, 16, 63. [Google Scholar] [CrossRef] [Green Version]
- Yako, T.; Otsu, W.; Nakamura, S.; Shimazawa, M.; Hara, H. Lipid Droplet Accumulation Promotes RPE Dysfunction. Int. J. Mol. Sci. 2022, 23, 1790. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.L.; Robison, W. Age-related changes in the retinal pigment epithelium of pigmented rats. Exp. Eye Res. 1984, 38, 137–151. [Google Scholar] [CrossRef]
- Bullock, J.; Polato, F.; Abu-Asab, M.; Bernardo-Colón, A.; Aflaki, E.; Agbaga, M.-P.; Becerra, S.P. Degradation of Photoreceptor Outer Segments by the Retinal Pigment Epithelium Requires Pigment Epithelium-Derived Factor Receptor (PEDF-R). Investig. Opthalmol. Vis. Sci. 2021, 62, 30. [Google Scholar] [CrossRef] [PubMed]
- Denk, H.; Pabst, D.; Abuja, P.M.; Reihs, R.; Tessaro, B.; Zatloukal, K.; Lackner, C. Senescence markers in focal nodular hy-perplasia of the liver: Pathogenic considerations on the basis of immunohistochemical results. Mod. Pathol. 2022, 35, 87–95. [Google Scholar] [CrossRef]
- Chaum, E.; Winborn, C.S.; Bhattacharya, S. Genomic regulation of senescence and innate immunity signaling in the retinal pigment epithelium. Mamm. Genome 2015, 26, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.L.; Fuchshofer, R.; Kook, D.; Kampik, A.; Bloemendal, H.; Welge-Lussen, U. Subtoxic oxidative stress induces senescence in retinal pigment epithelial cells via TGF-beta release. Investig. Ophthalmol. Vis. Sci. 2009, 50, 926–935. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-associated beta-galactosidase is lysosomal beta-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated be-ta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Sakemura, R.; Kumagai, A.; Sumikawa, E.; Fujii, M.; Ayusawa, D. Nuclear Swelling Occurs during Premature Senescence Mediated by MAP Kinases in Normal Human Fibroblasts. Biosci. Biotechnol. Biochem. 2008, 72, 1122–1125. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.B.; Park, K.R.; Kim, S.Y.; Han, S.-Y. Induction of Nuclear Enlargement and Senescence by Sirtuin Inhibitors in Glioblastoma Cells. Immune Netw. 2016, 16, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Kwon, W.; Freeman, S.A. Phagocytosis by the Retinal Pigment Epithelium: Recognition, Resolution, Recycling. Front. Immunol. 2020, 11, 604205. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, F.; Safa, H.; Finnemann, S.C. Understanding photoreceptor outer segment phagocytosis: Use and utility of RPE cells in culture. Exp. Eye Res. 2014, 126, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusiñol, A.E.; Sinensky, M.S. Farnesylated lamins, progeroid syndromes and farnesyl transferase inhibitors. J. Cell Sci. 2006, 119 Pt 16, 3265–3272. [Google Scholar] [CrossRef] [Green Version]
- De Sandre-Giovannoli, A.; Bernard, R.; Cau, P.; Navarro, C.; Amiel, J.; Boccaccio, I.; Lyonnet, S.; Stewart, C.L.; Munnich, A.; Le Merrer, M.; et al. Lamin a truncation in Hutchinson-Gilford progeria. Science 2003, 300, 2055. [Google Scholar] [CrossRef]
- Siniossoglou, S. Lipins, lipids and nuclear envelope structure. Traffic 2009, 10, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Notari, L.; Baladron, V.; Aroca-Aguilar, J.D.; Balko, N.; Heredia, R.; Meyer, C.; Notario, P.M.; Saravanamuthu, S.; Nueda, M.-L.; Sanchez-Sanchez, F.; et al. Identification of a lipase-linked cell membrane receptor for pigment epithelium-derived factor. J. Biol. Chem. 2006, 281, 38022–38037. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, R.; Strauss, J.G.; Haemmerle, G.; Schoiswohl, G.; Birner-Gruenberger, R.; Riederer, M.; Lass, A.; Neuberger, G.; Eisenhaber, F.; Hermetter, A.; et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 2004, 306, 1383–1386. [Google Scholar] [CrossRef] [Green Version]
- Sathyanarayan, A.; Mashek, M.T.; Mashek, D.G. ATGL Promotes Autophagy/Lipophagy via SIRT1 to Control Hepatic Lipid Droplet Catabolism. Cell Rep. 2017, 19, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Blasiak, J.; Pawlowska, E.; Sobczuk, A.; Szczepanska, J.; Kaarniranta, K. The Aging Stress Response and Its Implication for AMD Pathogenesis. Int. J. Mol. Sci. 2020, 21, 8840. [Google Scholar] [CrossRef]
- Guo, L.; Chen, Y.; Li, H.; Yin, F.; Ge, M.; Hu, L.; Zi, M.; Qin, Z.; He, Y. Telomere length is maternally inherited and associated with lipid metabolism in Chinese population. Aging 2022, 14, 354–367. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | Forward Primer | Reverse Primer |
|---|---|---|---|
| Rplp0 | RPLP0 | 5′-CTTCATTGTGGGAGCAGACA-3′ | 5′-GTGAGGTCCTCCTTGGTGAA-3′ |
| Serpinf1 | PEDF | 5′-ACCGTGACCCAGAACTTGAC-3′ | 5′-CACGGGTTTGCCAGTAATCT-3′ |
| Pnpla2 | PEDF-R | 5′-ACAGTGTCCCCATTCTCAGG-3′ | 5′-TTGGTTCAGTAGGCCATTCC-3′ |
| H2ax | H2AX | 5′-GCCTCATACCAGTTGACCCTG-3′ | 5′-TAGAACTCTTGTCCACAGGCC-3′ |
| Trp53 | P53 | 5′-GACCCTGGCACCTACAATGAA-3′ | 5′-GGGGTGGATAAATGCAGACA-3′ |
| Cdkn1a | P21 | 5′-ATCTGCTGCTCTTTTCCCCC-3′ | 5′-CCCTAGACCCACAATGCAGG-3′ |
| Glb1 | GLB1 | 5′-GCACGGCATCTATAATGTCACC-3′ | 5′-AGCCGGTCCTCCCAGTAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebustini, I.T.; Crawford, S.E.; Becerra, S.P. PEDF Deletion Induces Senescence and Defects in Phagocytosis in the RPE. Int. J. Mol. Sci. 2022, 23, 7745. https://doi.org/10.3390/ijms23147745
Rebustini IT, Crawford SE, Becerra SP. PEDF Deletion Induces Senescence and Defects in Phagocytosis in the RPE. International Journal of Molecular Sciences. 2022; 23(14):7745. https://doi.org/10.3390/ijms23147745
Chicago/Turabian StyleRebustini, Ivan T., Susan E. Crawford, and S. Patricia Becerra. 2022. "PEDF Deletion Induces Senescence and Defects in Phagocytosis in the RPE" International Journal of Molecular Sciences 23, no. 14: 7745. https://doi.org/10.3390/ijms23147745
APA StyleRebustini, I. T., Crawford, S. E., & Becerra, S. P. (2022). PEDF Deletion Induces Senescence and Defects in Phagocytosis in the RPE. International Journal of Molecular Sciences, 23(14), 7745. https://doi.org/10.3390/ijms23147745