
Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations

 , , ,
, , ,
Abstract
:1. Introduction
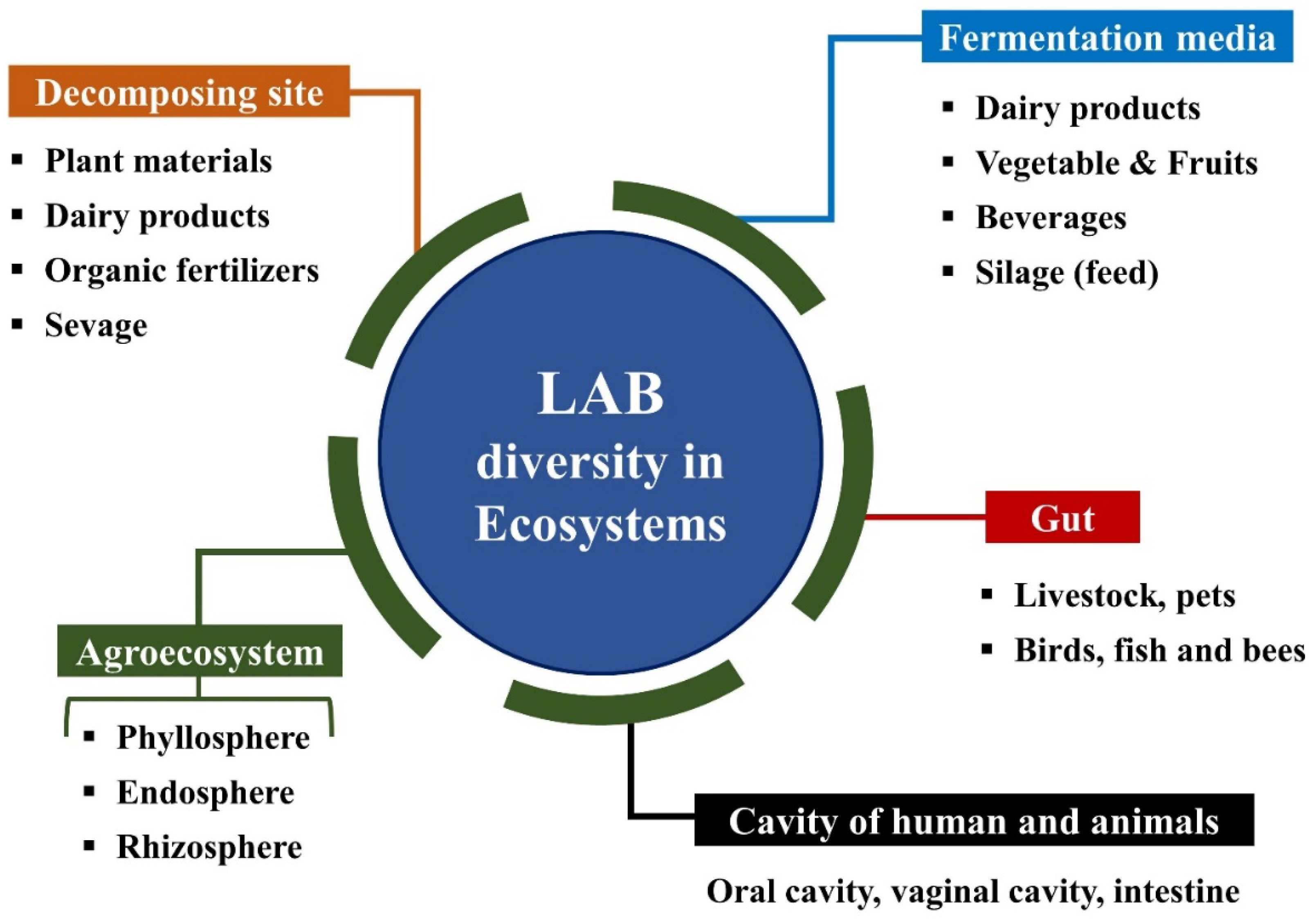
2. Lactic Acid Bacteria (LAB)
3. Biocontrol Agents of LAB
4. Antibacterial Activity of LAB
5. Antifungal Activity of LAB

{kind=link}
{kind=link}
{kind=link}
| Strains | Source | Active Compound | Active Spectrum | References |
|---|---|---|---|---|
| Antibacterial | ||||
| L. plantarum | Cucumber pickle | Organic acids | Pseudomonas campestris | [67] |
| LAB strain | Tomato rhizosphere | None | Ralstonia solanacearum, Xanthomonas campestris pv. vesicatoria, Pectobacterium carotovorum subsp. carotovorum | [65,66] |
| LAB strain | Unknown | None | Xanthomonas campestris pv. vesicatoria | [65] |
| L. lactis | Curd | Glycolipid biosurfactants | E. coli | [63] |
| Antifungal | ||||
| Lactobacillus species | Type culture | 3-Phenyllactic acid | P. expansum, A. flavus | [13] |
| L. acidophilus | Chicken intestine | Organic acid | Fusarium sp., Alternaria alternate | [36,77] |
| L. amylovorus | Gluten-free sourdough | Fatty acid, LA, salicyclic acid | P. paneum, Cladosporium sp., Rhizopus oryzae, Endomyces fibuliger, Aspergillus sp., Fusarium culmorum | [36,78,79] |
| L. brevis | Brewing barley | Organic acid, proteinaceous | A. flavus, F. culmorum, Trichophyton tonsurans, Eurotium repens, Penicillium sp. | [79,80,81]. |
| L casei | Dairy products | None | Trichophyton tonsurans, Penicillium sp. | [80,82] |
| L. coryniformis | Silage, flower, sourdough | PLA, proteinaceous | Aspergillus sp., Fusarium, Rhodotorula sp., Talaromyces flavus, Kluyveromyces sp. | [77,79] |
| L. fermentum | Fermented food and dairy products | Proteinaceous, PLA | A. niger, Fusarium graminearum, A. oryzae, A. niger, Fusarium sp. | [83,84] |
| L. harbinensis | Type strain | Fatty acids | Mucor racemosus | [85] |
| L. lactis | Wheat semolina | None | P. expansum | [82] |
| L. mesenteroides | Raw milk | LA, succinic acid, fatty acids | Penicillium species | [86] |
| L. plantarum | Plant materials, food grains, fermented soybean, raw milk | Fatty acids, LA, cyclic dipeptide, phenyllactic acid, peptides, succinic acid | Broad spectrum | [53,72,77,86,87,88,89,90,91] |
| L. paracasei | Dairy products, raw milk | Proteinaceous, LA, succinic acid, fatty acids | Fusarium sp. | [86,92] |
| L. pentosus | Fruit and fermented food | PLA | A. oryzae, A. niger, Fusarium sp. | [86] |
| Pediococcus pentosaceus | None | Proteinaceous, cyclic acids | Penicillium sp., Aspergillus sp., Fusarium sp., Rhizopus stolonifer, Sclerotium oryzae, Rhizoctonia solani, Botrytis cinerea, Sclerotinia minor, Rhodotorula sp. | [10,17,77,84] |
| L. reuteri | Murine gut, porcine | None | F. graminearum, A. niger, Fusarium sp. | [80,83] |
| L. sakei | Leaves, dandelions, flour | Peptide, PLA | A. fumigatus, Fusarium species | [77] |
| L. salivarius | Chicken intestine | Peptide, PLA | A. nidulans, F. sporotrichioies | [77] |
| Weissella cibaria | Food grains, fruits, and vegetables | Organic acids, proteinaceous | Fusarium culmorum, Penicillium sp., Aspergillus sp., Rhodotorula sp., Endomyces fibuliger | [10,18,93,94] |
| W. confuse | Food grains | Organic acids, proteinaceous | Penicillium sp., Aspergillus nidulans, Rhodotorula sp., Endomyces fibuliger | [10,70] |
| W. paramesenteroides | Fermented wax gourd | Organic acids | Penicillium sp., Fusarium graminearum, Rhizopus stolonifer, Sclerotium oryzae, Rhizoctonia solani, Botrytis cinerea, Sclerotinia minor | [17,93] |
6. Biopesticides and Insecticides of LAB
7. Biostimulants of LAB
8. Biofertilizer of LAB
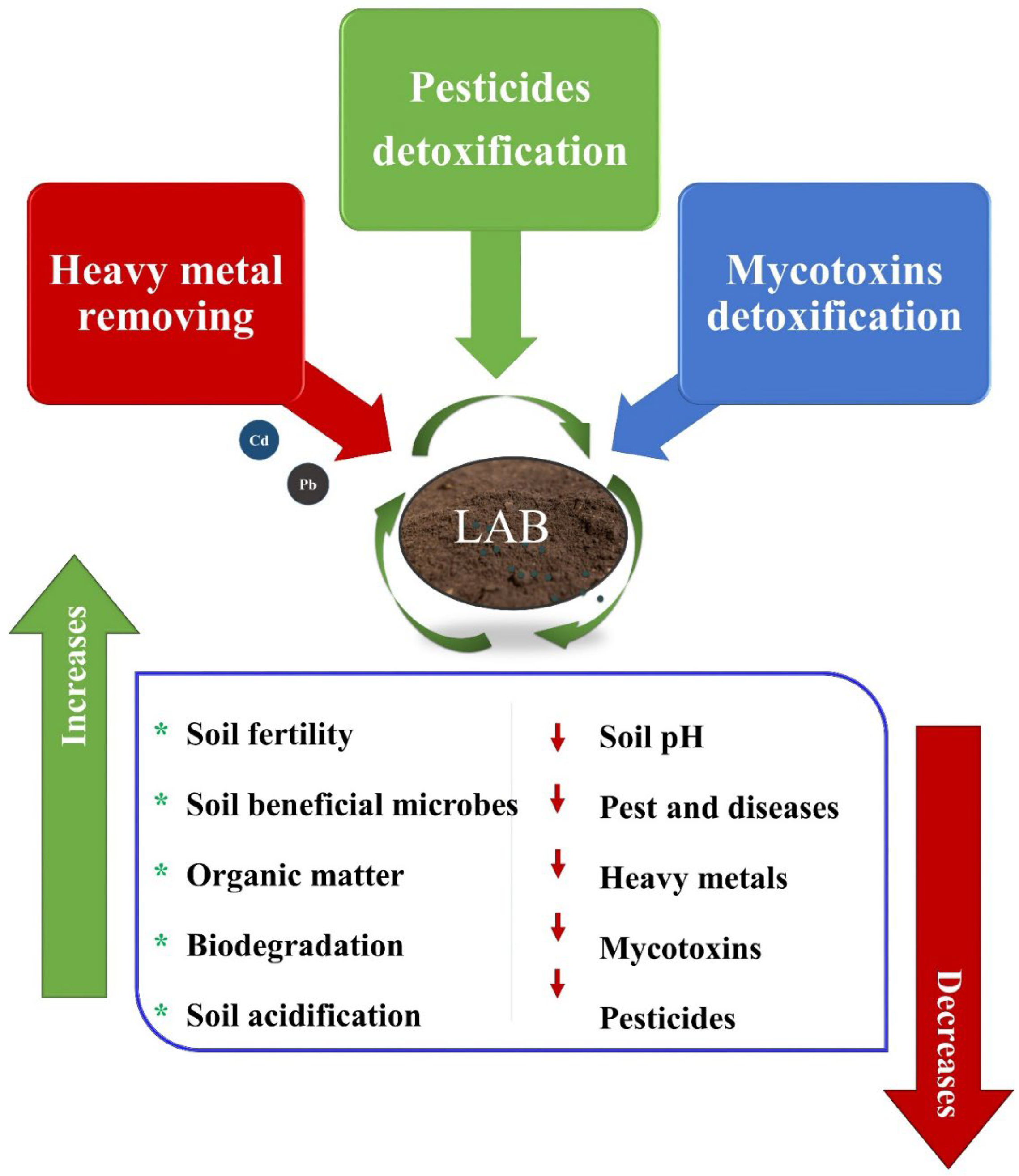
9. Soil Bioremediation of Lactic Acid Bacteria
10. Modern Technology and Metabolic Engineering of LAB
11. Limitations and Future Prospects of LAB
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Available online: http://www.fao.org/faostat/en/#country (accessed on 20 June 2020).
- Avis, T.J.; Gravel, V.R.; Antoun, H.; Tweddell, R.J. Multifaceted beneficial effects of rhizosphere microorganisms on plant health and productivity. Soil Biol. Biochem. 2008, 40, 1733–1740. [Google Scholar] [CrossRef]
- Lamont, J.R.; Wilkins, O.; Bywater-Ekegärd, M.; Smith, D.L. From yogurt to yield: Potential applications of lactic acid bacteria in plant production. Soil Biol. Biochem. 2017, 111, 1–9. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Perez-Muñoz, M.E.; Leulier, F.; Ganzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic Acid Bacteria as Antifungal and Anti-Mycotoxigenic Agents: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Yan, X.; Du, G.; Guo, Q.; Shi, Y.; Chang, J.; Wang, X.; Yuan, Y.; Yue, T. Recent developments in antifungal lactic acid bacteria: Application, screening methods, separation, purification of antifungal compounds and antifungal mechanisms. Crit. Rev. Food Sci. Nutr. 2021, 15, 1–15. [Google Scholar] [CrossRef]
- Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. Exploration of the Microbial Biodiversity Associated with North Apulian Sourdoughs and the Effect of the Increasing Number of Inoculated Lactic Acid Bacteria Strains on the Biocontrol against Fungal Spoilage. Fermentation 2019, 5, 97. [Google Scholar] [CrossRef] [Green Version]
- Gajbhiye, M.H.; Kapadnis, B.P. Antifungal-Activity-Producing Lactic Acid Bacteria as Biocontrol Agents in Plants. Biocontrol Sci. Technol. 2016, 26, 1451–1470. [Google Scholar] [CrossRef] [Green Version]
- Rouse, S.; Harnett, D.; Vaughan, A.; Sinderen, D. Lactic acid bacteria with potential to eliminate fungal spoilage in foods. J. Appl. Microbiol. 2008, 104, 915–923. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Siddiqui, A.J.; Hamadou, W.S.; Surti, M.; Awadelkareem, A.M.; Ashraf, S.A.; Alreshidi, M.; Snoussi, M.; Rizvi, S.M.D.; Bardakci, F.; et al. Inhibition of bacterial adhesion and antibiofilm activities of a glycolipid biosurfactant from Lactobacillus rhamnosus with its physicochemical and functional properties. Antibiotics 2021, 17, 1546. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Zavaleta, O.; Lopez-Malo, A.; Hernandez-Mendoza, A.; Garcia, H.S. Antifungal Activity of Lactobacilli and Its Relationship with 3-Phenyllactic Acid Production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [PubMed]
- Hashemi, S.M.B.; Jafarpour, D. Bioactive Edible Film Based on Konjac Glucomannan and Probiotic Lactobacillus plantarum Strains: Physicochemical Properties and Shelf Life of Fresh-Cut Kiwis. J. Food Sci. 2021, 86, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, K.; Samayanpaulraj, V.; Narayanadoss, V.; Uthandakalaipandian, R. Isolation of Lactic Acid Bacteria from Intestine of Freshwater Fishes and Elucidation of Probiotic Potential for Aquaculture Application. Probiotics Antimicrob. Proteins. 2021, 13, 1598–1610. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteriaas starter cultures: An update in their metabolism and genetics. Aims Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef]
- Sathe, S.; Nawani, N.; Dhakephalkar, P.; Kapadnis, B. Antifungal lactic acid bacteria with potential to prolong shelf-life of fresh vegetables. J. Appl. Microbiol. 2007, 103, 2622–2628. [Google Scholar] [CrossRef]
- Trias, R.; Bañeras, L.; Montesinos, E.; Badosa, E. Lactic acid bacteria from fresh fruit and vegetables as biocontrol agents of phytopathogenic bacteria and fungi. Int. Microbiol. 2008, 11, 231–236. [Google Scholar]
- Djadouni, F.; Kihal, M. Antimicrobial activity of lactic acid bacteria and the spectrum of their biopeptides against spoiling germs in foods. Braz. Arch. Biol. Technol. 2012, 55, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Pang, H.; Zhang, H.; Cai, Y. Biodiversity of lactic acid bacteria. In Lactic Acid Bacteria; Zhang, Y., Cai, Y., Eds.; Springer Science & Business Media: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Khalid, K. An overview of lactic acid bacteria. Int. J. Biosci. 2011, 1, 1–13. [Google Scholar]
- Hidalgo, D.; Corona, F.; Martín-Marroquin, J. Manure biostabilization by effective microorganisms as a way to improve its agronomic value. Biomass Convers. Bioref. 2022. [Google Scholar] [CrossRef]
- Han, H.; Ogata, Y.; Yamamoto, Y.; Nagao, S.; Nishino, N. Identification of lactic acid bacteria in the rumen and feces of dairy cows fed total mixed ration silage to assess the survival of silage bacteria in the gut. J. Dairy Sci. 2014, 97, 5754–5762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, L.C.; McFeeters, R.F.; Daeschel, M.A.; Fleming, H.P. A differential medium for the enumeration of homofermentative and heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 1987, 53, 1382–1384. [Google Scholar] [CrossRef] [Green Version]
- Minervini, F.; Celano, G.; Lattanzi, A.; Tedone, L.; De Mastro, G.; Gobbetti, M.; De Angelis, M. Lactic acid bacteria in durum wheat flour are endophytic components of the plant during its entire life cycle. Appl. Environ. Microbiol. 2015, 81, 6736–6748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekundayo, F.O. Isolation and identification of lactic acid bacteria from rhizosphere soils of three fruit trees, fish and ogi. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 991–998. [Google Scholar]
- Chen, Y.S.; Yanagida, F.; Shinohara, T. Isolation and identification of lactic acid bacteria from soil using an enrichment procedure. Lett. Appl. Microbiol. 2005, 40, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L. Organic acid in the rhizosphere—A critical review. Plant Soil. 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Fhoula, I.; Najjari, A.; Turki, Y.; Jaballah, S.; Boudabous, A.; Ouzari, H. Diversity and antimicrobial properties of lactic acid bacteria isolated from rhizosphere of olive trees and desert truffles of Tunisia. Biomed. Res. Int. 2013, 2013, 405708. [Google Scholar] [CrossRef] [Green Version]
- Varsha, K.K.; Nampoothiri, K.M. Appraisal of lactic acid bacteria as protective cultures. Food Control 2016, 69, 61–64. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajslova, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited “FAO estimate” of 25. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Wagacha, J.M.; Muthomi, J.W. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol. 2008, 124, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Paradhipta, D.H.V.; Joo, Y.H.; Lee, H.J.; Lee, S.S.; Noh, H.T.; Choi, J.S.; Kim, J.; Min, H.G.; Kim, S.C. Effects of Inoculants Producing Antifungal and Carboxylesterase Activities on Corn Silage and Its Shelf Life against Mold Contamination at Feed-Out Phase. Microorganisms 2021, 9, 558. [Google Scholar] [CrossRef] [PubMed]
- Hamed, H.A.; Moustafa, Y.A.; Abdel-Aziz, S.M. In vivo efficacy of lactic acid bacteria in biological control against Fusarium oxysporum for protection of tomato plant. Life Sci. 2011, 8, 462–468. [Google Scholar]
- Oliveira, P.M.; Zannini, E.; Arendt, E.K. Cereal fungal infection, mycotoxins, and lactic acid bacteria mediated bioprotection: From crop farming to cereal products. Food Microbiol. 2014, 37, 78–95. [Google Scholar] [CrossRef] [PubMed]
- Laury-Shaw, A.; Gragg, S.E.; Echeverry, A.; Brashears, M.M. Survival of Escherichia coli O157: H7 after application of lactic acid bacteria. J. Sci. Food Agric. 2019, 99, 1548–1553. [Google Scholar] [CrossRef]
- Prusky, D.; Kobiler, I.; Akerman, M.; Miyara, I. Effect of acidic solutions and acidic prochloraz on the control of postharvest decay caused by Alternaria alternata in mango and persimmon fruit. Postharvest Biol. Technol. 2006, 42, 134–141. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.; Chen, C.; Sun, Z.; Zhou, Y.; Shen, F.; Zhang, H.; Dai, Y. Genome shuffling of Lactobacillus plantarum for improving antifungal activity. Food Control 2013, 32, 341–347. [Google Scholar] [CrossRef]
- Crowley, S.; Mahony, J.; van Sinderen, D. Comparative analysis of two antifungal Lactobacillus plantarum isolates and their application as bioprotectants in refrigerated foods. J. Appl. Microbiol. 2012, 113, 1417–1427. [Google Scholar] [CrossRef]
- Gupta, R.; Srivastava, S. Antifungal effect of antimicrobial peptides (AMPs LR14) derived from Lactobacillus plantarum strain LR/14 and their applications in prevention of grain spoilage. Food Microbiol. 2014, 42, 1–7. [Google Scholar] [CrossRef]
- Ghosh, R.; Barman, S.; Mukhopadhyay, A.; Mandal, N.C. Biological control of fruit-rot of jackfruit by rhizobacteria and food grade lactic acid bacteria. Biol. Control. 2015, 83, 29–36. [Google Scholar] [CrossRef]
- Matei, G.M.; Matei, S.; Matei, A.; Cornea, C.P.; Draghici, E.M.; Jerca, I.O. Bioprotection of fresh food productsagainst blue mold using lactic acid bacteria with antifungal properties. Rom. Biotechnol. Lett. 2016, 21, 11201–11208. [Google Scholar]
- Lynch, K.M.; Zannini, E.; Guo, J.; Axel, C.; Arendt, E.K.; Kildea, S.; Coffey, A. Control of Zymoseptoria tritici cause of septoria tritici blotch of wheat using antifungal Lactobacillus strains. J. Appl. Microbiol. 2016, 121, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, N.; Cai, P.; Wu, D.; Pan, Y.; Curtis, J.M.; Ganzle, M.G. High-speed counter-current chromatography (HSCCC) purification of antifungal hydroxy unsaturated fatty acids from plant-seed oil and Lactobacillus cultures. J. Agric. Food Chem. 2017, 65, 11229–11236. [Google Scholar] [CrossRef] [PubMed]
- Kharazian, Z.A.; Jouzani, G.S.; Aghdasi, M.; Khorvash, M.; Zamani, M.; Mohammadzadeh, H. Biocontrol potential of Lactobacillus strains isolated from corn silages against some plant pathogenic fungi. Biol. Control 2017, 110, 33–43. [Google Scholar] [CrossRef]
- Juodeikiene, G.; Bartkiene, E.; Cernauskas, D.; Cizeikiene, D.; Zadeike, D.; Lele, V.; Bartkevics, V. Antifungal activity of lactic acid bacteria and their application for Fusarium mycotoxin reduction in malting wheat grains. LWT 2018, 89, 307–314. [Google Scholar] [CrossRef]
- Ma, J.; Hong, Y.; Deng, L.; Yi, L.; Zeng, K. Screening and characterization of lactic acid bacteria with antifungal activity against Penicillium digitatum on citrus. Biol. Control 2019, 138, 104044. [Google Scholar] [CrossRef]
- Khodaei, D.; Hamidi-Esfahani, Z. Influence of bioactive edible coatings loaded with Lactobacillus plantarum on physicochemical properties of fresh strawberries. Postharvest Biol. Technol. 2019, 156, 110944. [Google Scholar] [CrossRef]
- Omedi, J.O.; Huang, W.; Zheng, J. Effect of sourdough lactic acid bacteria fermentation on phenolic acid release and antifungal activity in pitaya fruit substrate. Food Sci. Technol. 2019, 111, 309–317. [Google Scholar] [CrossRef]
- Luz, C.; D’Opazo, V.; Quiles, J.M.; Romano, R.; Manes, J.; Meca, G. Biopreservation of tomatoes using fermented media by lactic acid bacteria. LWT 2020, 130, 109618. [Google Scholar] [CrossRef]
- Li, Z.; Wang, L.; Xie, B.; Hu, S.; Zheng, Y.; Jin, P. Effects of exogenous calcium and calcium chelant on cold tolerance of postharvest loquat fruit. Sci. Hortic. 2020, 269, 109391. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Algboory, H.L.; Kadum, H.; Mohammed, N.K.; Saari, N.; Hassan, Z.; Hussin, A.S.M. Antifungal activity determination for the peptides generated by Lactobacillus plantarum TE10 against Aspergillus flavus in maize seeds. Food Control 2020, 109, 106898. [Google Scholar] [CrossRef]
- De Simone, N.; Capozzi, V.; de Chiara, M.L.V.; Amodio, M.L.; Brahimi, S.; Colelli, G.; Drider, D.; Spano, G.; Russo, P. Screening of Lactic Acid Bacteria for the Bio-Control of Botrytis cinerea and the Potential of Lactiplantibacillus plantarum for Eco-Friendly Preservation of Fresh-Cut Kiwifruit. Microorganisms 2021, 9, 773. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.B.; Nes, I.F. Ribosomally synthesized antibacterial peptides in Gram positive bacteria. Curr. Drug Targets 2002, 3, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, P. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Montville, T.J.; Chen, Y. Mechanistic action of pediocin and nisin: Recent progress and unresolved questions. Appl. Microbiol. Biotechnol. 1998, 50, 511–519. [Google Scholar] [CrossRef]
- Rooney, W.M.; Grinter, R.W.; Correia, A.; Parkhill, J.; Walker, D.C.; Milner, J.J. Engineering bacteriocin-mediated resistance against the plant pathogen Pseudomonas syringae. Plant Biotechnol. J. 2020, 18, 1296–1306. [Google Scholar] [CrossRef] [Green Version]
- Quadriya, H.; Ali, S.A.M.; Parameshwar, J.; Manasa, M.; Khan, M.Y.; Hameeda, B. Microbes Living Together: Exploiting the Art for Making Biosurfactants and Biofilms. In Implication of Quorum Sensing System in Biofilm Formation and Virulence; Springer: Berlin/Heidelberg, Germany, 2018; pp. 161–177. [Google Scholar]
- Rodrigues, L.; van der Mei, H.C.; Teixeira, J.; Oliveira, R. Biosurfactant from Lactococcus lactis 53 inhibits microbial adhesion on silicone rubber. Appl. Microbiol. Biotechnol. 2004, 66, 306–311. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumari, P.; Mani, K. Structural characterization of a novel xylolipid biosurfactant from Lactococcus lactis and analysis of antibacterial activity against multi-drug resistant pathogens. Bioresour. Technol. 2010, 101, 8851–8854. [Google Scholar] [CrossRef]
- Ahn, K.B.; Baik, J.E.; Park, O.J.; Yun, C.H.; Han, S.H. Lactobacillus plantarum lipoteichoic acid inhibits biofilm formation of Streptococcus mutans. PLoS ONE 2018, 13, e0192694. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, A.; Kim, E.C.; Lim, C.K.; Cho, S.Y.; Hur, J.H.; Park, D.H. Biological control of soft rot on Chinese cabbage using beneficial bacterial agents in greenhouse and field. Korean J. Pestic. Sci. 2009, 13, 325–331. [Google Scholar]
- Shrestha, A.; Choi, K.U.; Lim, C.K.; Hur, J.H.; Cho, S.Y. Antagonistic effect of Lactobacillus sp. Strain KLF01 against plant pathogenic bacteria Ralstonia solanacearum. J. Pestic. Sci. 2009, 13, 45–53. [Google Scholar]
- Visser, R.; Holzapfel, W.H.; Bezuidenhout, J.J.; Kotze, J.M. Antagonism of lactic acid bacteria against phytopathogenic bacteria. Appl. Environ. Microbiol. 1986, 52, 552–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baffoni, L.; Gaggia, F.; Dalanaj, N.; Prodi, A.; Nipoti, P.; Pisi, A.; Biavati, B.; Di Gioia, D. Microbial inoculants for the biocontrol of Fusarium spp. in durum wheat. BMC Microbiol. 2015, 15, 242. [Google Scholar] [CrossRef]
- Tsitsigiannis, D.I.; Dimakopoulou, M.; Antoniou, P.P.; Tjamos, E.C. Biological control strategies of mycotoxigenic fungi and associated mycotoxins in Mediterranean basin crops. Phytopathol. Mediterr. 2012, 51, 158–174. [Google Scholar]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef]
- Lipinska, L.; Klewicki, R.; Sojka, M.; Bonikowski, R.; Zyzelewiz, D.; Kolodziejczyk, K.; Klrwicka, E. Antifungal Activity of Lactobacillus pentosus ŁOCK 0979 in the Presence of Polyols and Galactosyl-Polyols. Probiotics Antimicro. Proteins 2018, 10, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Sjogren, J.; Magnusson, J.; Broberg, A.; Schnürer, J.; Kenne, L. Antifungal 3-hydroxy fatty acids from Lactobacillus plantarum MiLAB 14. Appl. Environ. Microbiol. 2003, 69, 7554–7557. [Google Scholar] [CrossRef] [Green Version]
- Lappa, I.K.; Mparampouti, S.; Lanza, B.; Panagou, E.Z. Control of Aspergillus carbonarius in grape berries by Lactobacillus plantarum: A phenotypic and gene transcription study. Int. J. Food Microbiol. 2018, 275, 56–65. [Google Scholar] [CrossRef]
- Pohl, C.H.; Kock, J.L.F.; Thibane, V.S. Antifungal free fatty acids: A review. In Science against Microbial Pathogens: Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2011; Volume 1, pp. 61–71. [Google Scholar]
- Dopazo, V.; Luz, C.; Quiles, J.M.; Calpe, J.; Romano, R.; Manes, J.; Meca, G. Potential application of lactic acid bacteria in the biopreservation of red grape from mycotoxigenic fungi. J. Sci. Food Agric. 2022, 102, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Marin, A.; Plotto, A.; Atares, L.; Chiralt, A. Lactic acid bacteria incorporated into edible coatings to control fungal growth and maintain postharvest quality of grapes. HortScience 2019, 54, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, J.; Strom, K.; Roos, S.; Sjogren, J.; Schnürer, J. Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol. Lett. 2003, 219, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Ryan, L.A.M.; Zannini, E.; Dal Bello, F.B.; Pawlowska, A.; Koehler, P.; Arendt, E.K. Lactobacillus amylovorus DSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food Microbiol. 2011, 146, 276–283. [Google Scholar] [CrossRef] [PubMed]
- De Muynck, C.; Leroy, A.I.J.; De Maeseneire, S.; Arnaut, F.; Soetaert, W.; Vandamme, E.J. Potential of selected lactic acid bacteria to produce food compatible antifungal metabolites. Microbiol. Res. 2004, 159, 339–346. [Google Scholar] [CrossRef]
- Guo, J.; Mauch, A.; Galle, S.; Murphy, P.; Arendt, E.; Coffey, A. Inhibition of growth of Trichophyton tonsurans by Lactobacillus reuteri. J. Appl. Microbiol. 2011, 111, 474–483. [Google Scholar] [CrossRef]
- Falguni, P.; Shilpa, V.; Mann, B. Production of proteinaceous antifungal substances from Lactobacillus brevis NCDC 02. Int. J. Dairy Technol. 2010, 63, 70–76. [Google Scholar] [CrossRef]
- Florianowicz, T. Antifungal activity of some microorganisms against Penicillium expansum. Eur. Food Res. Technol. 2001, 212, 282–286. [Google Scholar] [CrossRef]
- Gerez, C.; Torres, M.J.; Font de Valdez, G.; Rollan, G. Control of spoilage fungi by lactic acid bacteria. Biol. Control 2012, 64, 231–237. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Hassan, Z.; Sadon, S.K.; Zulkifli, N.A.; Azfar, A. Effect of pH and heat treatment on antifungal activity of Lactobacillus fermentum Te007, Lactobacillus pentosus G004 and Pediococcus pentosaceus Te010. Innov. Rom. Food Biotechnol. 2011, 8, 41–53. [Google Scholar]
- Salas, M.L.; Mounier, J.; Maillard, M.B.; Valence, F.; Coton, E.; Thierry, A. Identification and quantification of natural compounds produced by antifungal bioprotective cultures in dairy products. Food Chem. 2019, 301, 125260. [Google Scholar] [CrossRef]
- Ouiddir, M.; Bettache, G.; Salas, M.L.; Pawtowski, A.; Donot, C.; Brahimi, S.; Mabrouk, K.; Coton, E.; Mounier, J. Selection of Algerian lactic acid bacteria for use as antifungal bioprotective cultures and application in dairy and bakery products. Food Microbiol. 2019, 82, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsettti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21 B. Appl. Environ. Microbiol. 2000, 6, 4084–4090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strom, K.; Sjogren, J.; Broberg, A.; Schnürer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides cyclo(L-Phe-L-Pro) and cyclo(L-Phe-trans-4-OH-L-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [Green Version]
- Sangmanee, P.; Hongpattarakere, T. Inhibitory of multiple antifungal components produced by Lactobacillus plantarum K35 on growth, aflatoxin production and ultrastructure alterations of Aspergillus flavus and Aspergillus parasiticus. Food Control 2014, 40, 224–233. [Google Scholar] [CrossRef]
- Dal Bello, F.; Clarke, C.I.; Ryan, L.A.M.; Ulmer, H.; Schober, T.J.; Strom, K.; van Sinderen, D.; Schnurer, J.; Arendt, E.K. Improvement of the quality and shelf life of wheat bread by fermentation with the antifungal strain Lactobacillus plantarum FST 1.7. J. Cereal Sci. 2007, 45, 309–318. [Google Scholar] [CrossRef]
- Yang, E.J.; Chang, H.C. Purification of a new antifungal compound produced by Lactobacillus plantarum AF1 isolated from kimchi. Int. J. Food Microbiol. 2010, 139, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Franco, T.; Garcia, S.; Hirooka, E.; Ono, Y.; dos Santos, J. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J. Appl. Microbiol. 2011, 111, 739–748. [Google Scholar] [CrossRef]
- Lan, W.; Chen, Y.; Wu, H.; Yanagida, F. Bio-protective potential of lactic acid bacteria isolated from fermented wax gourd. Folia Microbiol. 2012, 57, 99–105. [Google Scholar] [CrossRef]
- Valerio, F.; Favilla, M.; De Bellis, P.; Sisto, A.; de Candia, S.; Lavermicocca, P. Antifungal activity of strains of lactic acid bacteria isolated from a semolina ecosystem against Penicillium roqueforti, Aspergillus niger and Endomyces fibuliger contaminating bakery products. Syst. Appl. Microbiol. 2009, 32, 438–448. [Google Scholar] [CrossRef]
- Skendzic, S.; Zovko, M.; Zivković, I.P.; Lesic, V.; Lemic, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Karami-Mohajeri, S.; Ahmadipour, A.; Rahimi, H.R.; Abdollahi, M. Adverse effects of organophosphorus pesticides on the liver: A brief summary of four decades of research. Arh. Hig. Rada. Toksikol. 2017, 68, 261–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiu, L. Microbial Biopesticides in Agroecosystems. Agronomy 2018, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; Park, J.H.; Choi, I.S.; Wi, S.G.; Ha, S.; Chun, H.H.; Hwang, I.M.; Chang, J.Y.; Choi, H.J.; Kim, J.C.; et al. Effective approach to organic acid production from agricultural kimchi cabbage waste and its potential application. PLoS ONE 2018, 13, e0207801. [Google Scholar] [CrossRef] [PubMed]
- Alawamleh, A.; Durovic, G.; Maddalena, G.; Guzzon, R.; Ganassi, S.; Hashmi, M.M.; Wäckers, F.; Anfora, G.; Cristofaro, A.D. Selection of Lactic Acid Bacteria Species and Strains for Efficient Trapping of Drosophila suzukii. Insects 2021, 12, 153. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahin, A.; Sonomoto, K. Nisin tolerance of DnaK-overexpressing Lactococcus lactis strains at 40 °C. Am. J. Biochem. Mol. Biol. 2012, 2, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Takei, T.; Yoshida, M.; Hatate, Y.; Shiomori, K.; Kiyoyama, S. Lactic acid bacteria-enclosing poly(epsilon-caprolactone) microcapsules as soil bioamendment. J. Biosci. Bioeng. 2008, 106, 268–272. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Kavallieratos, N.G.; Benelli, G.; Losic, D.; Rani, P.U.; Desneux, N. Nanoparticles for pest control: Current status and future perspectives. J. Pest Sci. 2018, 91, 1–15. [Google Scholar] [CrossRef]
- Chen, H.D.; Yada, R.N. Nanotechnologies in agriculture: New tools for sustainable development. Trends Food Sci. Technol. 2011, 22, 585–594. [Google Scholar] [CrossRef]
- Dikbas, N.; Ucar, S.; Tozlu, G.; Ozer, T.O.; Kotan, R. Bacterial Chitinase Biochemical Properties, Immobilization on Zinc Oxide (ZnO) Nanoparticle and its Effect on Sitophilus zeamais as a Potential Insecticide. World J. Microbiol. Biotechnol. 2021, 37, 173. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Klimova, S.Y.; Cherdyntseva, T.A.; Netrusov, A.I. Microbial producers of plant growth stimulators and their practical use: A review. Appl. Biochem. Microbiol. 2006, 42, 117–126. [Google Scholar] [CrossRef]
- Amprayna, K.; Supawonga, V.; Kengkwasingha, P.; Getmalab, A. Plant Growth Promoting Traits of Lactic Acid Bacterium Isolated from Rice Rhizosphere and Its Effect on Rice Growth. In Proceedings of the 5th Burapha University International Conference STP-029-10, Pattaya, Thailand, 28–29 July 2016. [Google Scholar]
- Lynch, J.M. Origin, Nature and Biological Activity of Aliphatic Substances and Growth Hormones Found in Soil. In Soil Organic Matter and Biological Activity. Developments in Plant and Soil Sciences; Vaughan, D., Malcolm, R.E., Eds.; Springer: Dordrecht, The Netherlands, 1985; Volume 16. [Google Scholar]
- Somers, E.; Amake, A.; Croonenborghs, A.; Overbeek, L.S.; Vanderleyden, J. Lactic acid bacterial in organic agricultural soil. In Proceedings of the Rhizosphere 2, Montpellier, France, 26–31 August 2007. [Google Scholar]
- Higa, T.; Kinjo, S. Effect of lactic acid fermentation bacteria on plant growth and soil humus formation. In Proceedings of the First International Conference on Kyusei Nature Farming, Khon Kaen, Thailand, 17–21 October 1989; Parr, J.F., Hornick, S.B., Whitman, C.E., Eds.; US Department of Agriculture: Washington, DC, USA, 1991; pp. 140–147. [Google Scholar]
- Rusch, H.P. Bodenfruchtbarkeit; Karl F. Haug Verlag: Heidelberg, Germany, 1964. [Google Scholar]
- Kang, S.M.; Radhakrishnan, R.; You, Y.H.; Khan, A.L.; Park, J.M.; Lee, S.M.; Lee, I.J. Cucumber performance is improved by inoculation with plant growth-promoting microorganisms. Acta Agric. Scand. B soil Plant Sci. 2015, 65, 36–44. [Google Scholar] [CrossRef]
- Lutz, M.P.; Michel, V.; Martinez, C.; Camps, C. Lactic acid bacteria as biocontrol agents of soil-borne pathogens biological control of fungal and bacterial plant pathogens. Biol. Control Fungal Bact. Plant Pathog. 2012, 78, 285–288. [Google Scholar]
- Shrestha, A.; Kim, B.S.; Park, D.H. Biological control of bacterial spot disease and plant growth-promoting effects of lactic acid bacteria on pepper. Biocontrol Sci. Technol. 2014, 24, 763–779. [Google Scholar] [CrossRef]
- Strafella, S.; Simpson, D.J.; Yaghoubi Khanghahi, M.; De Angelis, M.; Ganzle, M.; Minervini, F.; Crecchio, C. Comparative genomics and in vitro plant growth promotion and biocontrol traits of lactic acid bacteria from the wheat rhizosphere. Microorganisms 2021, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, A.; Okada, S.; Hols, P.; Satoh, E. Metabolic engineering of Lactobacillus plantarum for succinic acid production through activation of the reductive branch of the tricarboxylic acid cycle. Enzym. Microb. Technol. 2013, 53, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Limanska, N.; Ivanytsia, T.; Basiul, O.; Krylova, K.; Biscola, V.; Chobert, J.M.; Ivanytsia, V.; Haertle, T. Effect of Lactobacillus plantarum on germination and growth of tomato seedlings. Acta Physiol. Plant. 2013, 35, 1587–1595. [Google Scholar] [CrossRef]
- Rzhevskaya, V.S.; Oturina, I.P.; Oturina, L.M. Teplitskaya Study of the biological characteristics of the lactic acid bacteria strains. Серuя Бuoлoгuя Xuмuя 2014, 27, 145–160. [Google Scholar]
- Yarullina, D.R.; Asafova, E.V.; Kartunova, J.E.; Ziyatdinova, G.K.; Ilinskaya, O.N. Probiotics for plants: NO-producing lactobacilli protect plants from drought. Appl. Biochem. Microbiol. 2014, 50, 166–168. [Google Scholar] [CrossRef]
- Phoboo, S.; Sarkar, D.; Bhowmik, P.C.; Jha, P.K.; Shetty, K. Improving salinity resilience in Swertia chirayita clonal line with Lactobacillus plantarum. Can. J. Plant Sci. 2016, 96, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Mohite, B. Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nut. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Giassi, V.; Kiritani, C.; Kupper, K.C. Bacteria as growth-promoting agents for citrus rootstocks. Microbiol. Res. 2016, 190, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bi, L.; Liao, Y.; Lu, D.; Zhang, H.; Liao, X.; Liang, J.B.; Wu, Y. Influence and characteristics of Bacillus stearothermophilus in ammonia reduction during layer manure composting. Ecotoxicol. Environ. Saf. 2019, 180, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Blais, A. Lactic Acid and Bacillaceae Fertilizer and Method of Producing Same. Canadian Patent No. CA2598539A1, 31 August 2006. Available online: https://patents.google.com/patent/CA2598539A1/un (accessed on 16 June 2022).
- 2020 Industry Report: Mushroom, 2021, Market Intelligence Team. Available online: https://www.fortunebusinessinsights.com (accessed on 16 June 2022).
- Raman, J.; Lee, S.K.; Im, J.H.; Oh, M.J.; Oh, Y.L.; Jang, K.Y. Current prospects of mushroom production and industrial growth in India. J. Mushrooms 2018, 16, 239–249. [Google Scholar]
- HaqMaher, M.J.; Smyth, S.; Dodd, V.A.; McCabe, T.; Magette, W.L.; Duggan, J.; Hennerty, M.J. Managing Spent Mushroom Compost; Teagasc: Dublin, Ireland, 2000; pp. 111–121. [Google Scholar]
- Kwiatkowski, C.A.; Harasim, E. The Effect of Fertilization with Spent Mushroom Substrate and Traditional Methods of Fertilization of Common Thyme (Thymus vulgaris L.) on Yield Quality and Antioxidant Properties of Herbal Material. Agronomy 2021, 11, 329. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, Y.H.; Kim, Y.I.; Ahmadi, F.; Oh, Y.K.; Park, J.M.; Kwak, W.S. Effect of microbial inoculant or molasses on fermentative quality and aerobic stability of sawdust-based spent mushroom substrate. Bioresour. Technol. 2016, 216, 188–195. [Google Scholar] [CrossRef]
- Chuang, W.Y.; Liu, C.L.; Tsai, C.F.; Lin, W.C.; Chang, S.C.; Shih, H.; Shy, Y.M.; Lee, T.T. Evaluation of Waste Mushroom Compost as a Feed Supplement and Its Effects on the Fat Metabolism and Antioxidant Capacity of Broilers. Animals 2020, 10, 445. [Google Scholar] [CrossRef] [Green Version]
- Cacace, C.; Rizzello, C.G.; Brunetti, G.; Verni, M.; Cocozza, C. Reuse of Wasted Bread as Soil Amendment: Bioprocessing, Effects on Alkaline Soil and Escarole (Cichorium endivia) Production. Foods 2022, 11, 189. [Google Scholar] [CrossRef]
- Cocozza, C.; Ercolani, G.L. Siderophore production and associated characteristics in rhizosphere and non-rhizosphere fluorescent pseudomonads. Ann. Microbiol. 1997, 47, 17–28. [Google Scholar]
- Sposito, G. The Chemistry of Soil; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Mao, B.; Yin, R.; Li, X.; Cui, S.; Zhang, H.; Zhao, J.; Chen, W. Comparative Genomic Analysis of Lactiplantibacillus plantarum Isolated from Different Niches. Genes 2021, 12, 241. [Google Scholar] [CrossRef]
- Yan, Y.H.; Zhang, F.; Chai, Z.Y.; Liu, M.; Battino, M.; Meng, X.H. Mixed fermentation of blueberry pomace with L. rhamnosus GG and L. plantarum-1: Enhance the active ingredient, antioxidant activity and health promoting benefits. Food Chem. Toxicol. 2019, 131, 110541. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Salminen, S.; Tahvonen, R. Rapid removal of lead and cadmium from water by specific lactic acid bacteria. Int. J. Food Microbiol. 2007, 114, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Haskard, C.; El-Nezami, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Surface binding of aflatoxin B1 by lactic acid bacteria. Appl. Environ. Microbiol. 2001, 67, 3086–3091. [Google Scholar] [CrossRef] [Green Version]
- Kromah, V.; Zhang, G. Aqueous Adsorption of Heavy Metals on Metal Sulfide Nanomaterials: Synthesis and Application. Water 2021, 13, 1843. [Google Scholar] [CrossRef]
- Ameen, F.A.; Hamdan, A.M.; El-Naggar, M.Y. Assessment of the heavy metal bioremediation efficiency of the novel marine lactic acid bacterium, Lactobacillus plantarum MF042018. Sci. Rep. 2020, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Kirillova, A.V.; Danilushkina, A.A.; Irisov, D.S.; Bruslik, N.L.; Fakhrullin, R.F.; Zakharov, Y.A.; Bukhmin, V.S.; Yarullina, D.R. Assessment of Resistance and Bioremediation Ability of Lactobacillus Strains to Lead and Cadmium. Int. J. Microbiol. 2017, 2017, 9869145. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, H. Biosorption of Heavy Metals by Lactic Acid Bacteria for Detoxification. Methods Mol. Biol. 2019, 1887, 145–157. [Google Scholar]
- Bo, L.Y.; Zhang, Y.H.; Zhao, X.H. Degradation kinetics of seven organophosphorus pesticides in milk during yoghurt processing. J. Serb. Chem. Soc. 2011, 76, 353–362. [Google Scholar] [CrossRef]
- Islam, M.A.; Math, R.K.; Cho, K.M.; Lim, W.J.; Hong, S.Y.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Yun, H.D. Organophosphorus hydrolase (OpdB) of Lactobacillus brevis WCP902 from kimchi is able to degrade organophosphorus pesticides. J. Agri. Food Chem. 2010, 58, 5380–5386. [Google Scholar] [CrossRef]
- Zhou, X.W.; Zhao, X.H. Susceptibility of nine organophosphorus pesticides in skimmed milk towards inoculated lactic acid bacteria and yogurt starters. J. Sci. Food Agric. 2015, 95, 260–266. [Google Scholar] [CrossRef]
- Papagianni, M. Metabolic engineering of lactic acid bacteria for the production of industrially important compounds. Comput. Struct. Biotechnol. J. 2012, 29, e201210003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Barrangou, R. Applications of CRISPR-Cas systems in lactic acid bacteria. FEMS Microbiol. Rev. 2020, 44, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Auras, R.; Lim, L.T.; Selke, S.E.M.; Tsuji, H. Poly (Lactic Acid): Synthesis, Structures, Properties, Processing, and Applications; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Soundharrajan, I.; Park, H.S.; Rengasamy, S.; Sivanesan, R.; Choi, K.C. Application and Future Prospective of Lactic Acid Bacteria as Natural Additives for Silage Production—A Review. Appl. Sci. 2021, 11, 8127. [Google Scholar] [CrossRef]
- Alonso, S.; Castrol, M.C.; Berdascol, M.; de la Banda, I.G.; Moreno-Ventas, X.; de Rojas, A.H. Isolation and partial characterization of lactic acid bacteria from the gut microbiota of marine fishes for potential application as probiotics in aquaculture. Probiotics Antimicrob. Proteins 2019, 11, 569–579. [Google Scholar] [CrossRef]



| Strain Name (LAB) | Pathogens | Food Crops | References |
|---|---|---|---|
| LAB | Alternaria alternata | Post-harvest decay | [38] |
| Lactobacillus plantarum CUK-501 | Aspergillus flavu, Fusarium graminearum, Rhizopus stolonifer, B. cinerea | Cucumber | [17] |
| LAB | Bacteria and fungi | Vegetables and fruits | [18] |
| L. plantarum IMAU10014, | Penicillium digitatum | Citrus japonica (kumquat), | [39] |
| Pediococcus pentosaceous | P. expansum | Pyrus (pear), Vitis vinifera (grape), Prunus (plum) | [40] |
| L. plantarum LR/14 | A. niger, R. stolonifer, Mucor racemosus, P. chrysogenum | Wheat seeds | [41] |
| LAB | Fusarium | Cereal-based products | [36] |
| Lactococcus lactis subsp. lactis | Rhizopus stolonifer | Artocarpus heterophyllus (jackfruit) | [42] |
| Lactic acid bacteria 43, LCM5 | Penicillium expansum | Malus domestica (apple) | [43] |
| LAB | Zymoseptoria tritici | Wheat | [44] |
| L. plantarum | Filamentous fungi and yeast | - | [45] |
| Lactobacilli | F. verticillioides | Ensiled corns | [46] |
| LAB | Fusarium malting | Wheat grains | [47] |
| L. sucicola, P. acidilactici | P. digitatum | Citrus | [48] |
| L. plantarum | - | Fragaria x ananassa (strawberry) | [49] |
| L. plantarum, L. pentosus, P. pentosaceus | A. niger, Cladosporium sphaerospermum, P. chrysogenum | Pitaya (cactus fruit) | [50] |
| L. plantarum TR7 | P. expansum | Solanum lycopersicum (tomato) | [51] |
| LAB | Blackening | Banana | [52] |
| L. plantarum TE10 | Aspergillus flavus | Fresh maize seeds | [53] |
| L. plantarum | Botrytis cinerea | Horticultural crops | [54] |
| Strains | Source | Crops | Effects | Mechanisms | References |
|---|---|---|---|---|---|
| L. plantarum | EM-4, type strain, grape must | Radish, tomato | Increased yield, shoot branching, shoot and root growth | None | [35,109] |
| L. plantarum | Grape must, oyster mushroom | Tomato | Increased germination, increased shoot and root growth | Bacteriogenic metabolites | [116] |
| L. plantarum | Commercial phytostimulant | Cucumber | Increased germination and seedling growth | None | [117] |
| L. plantarum | Dairy products | Tomato | Increasing germination rate and root growth | Bacteriogenic metabolites | [116] |
| L. plantarum | Human probiotic | Wheat | Osmotic stress alleviation | None | [118] |
| L. plantarum | PGPR Corp. (Korea) | Cucumber | Increased growth, nutrient uptake, and amino acid content | Increased nutrient availability via succinic acid and LA | [111] |
| L. plantarum | Unknown | Swertia chirayita | Salt stress tolerant | Stress response | [119] |
| L. acidophilus | Dairy products | Tomato | Increased shoot branching, shoot and root growth | None | [35] |
| Lactobacillus sp. | Dairy products | Tomato | Increased shoot branching, shoot and root growth | None | [35] |
| LAB | Unknown | Pepper | Biocontrol and biostimulant property | IAA and siderophores | [112] |
| L. acidophilus | Wheat rhizosphere | Wheat | Increased plant length and chlorophyll content | IAA | [120] |
| L. casei | Commercial phytostimulant | Cucumber | Increased germination rate | None | [117] |
| LAB strain KLF01 | Tomato rhizosphere | Pepper | Increased root and shoot length, root fresh weight, and chlorophyll content | IAA, phosphate solubilization | [113] |
| LAB strain KLCO2, KPD03 | Unknown | Pepper | Increased root and shoot length, root fresh weight and chlorophyll content | IAA, phosphate solubilization | [113] |
| LAB strain BL06 | Sugarcane ferment | Citrus seedling | Increased height, stem diameter, root and shoot weight | Phosphate solubilization, nitrogen fixation | [121] |
| LAB | None | None | PGP properties | IAA and mineral solubilization | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raman, J.; Kim, J.-S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.-J.; Kim, S.-J. Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. Int. J. Mol. Sci. 2022, 23, 7784. https://doi.org/10.3390/ijms23147784
Raman J, Kim J-S, Choi KR, Eun H, Yang D, Ko Y-J, Kim S-J. Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. International Journal of Molecular Sciences. 2022; 23(14):7784. https://doi.org/10.3390/ijms23147784
Chicago/Turabian StyleRaman, Jegadeesh, Jeong-Seon Kim, Kyeong Rok Choi, Hyunmin Eun, Dongsoo Yang, Young-Joon Ko, and Soo-Jin Kim. 2022. "Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations" International Journal of Molecular Sciences 23, no. 14: 7784. https://doi.org/10.3390/ijms23147784
APA StyleRaman, J., Kim, J. -S., Choi, K. R., Eun, H., Yang, D., Ko, Y. -J., & Kim, S. -J. (2022). Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. International Journal of Molecular Sciences, 23(14), 7784. https://doi.org/10.3390/ijms23147784