1. Introduction
Understanding hepatocyte concentrations of substrates is a recent translational topic. In the past, great achievements in the understanding of substrate distribution in the liver have been primarily made through the measurement of plasma concentrations. Liver concentrations were not available. Therefore, when conducting pharmacokinetic studies, the classical approach assumed that unbound concentrations approximate unbound plasma concentrations. With the emerging knowledge of hepatocyte membrane transporters, it became clear that, depending on the relative hepatocyte influx and efflux clearances of substrates, unbound hepatocyte concentrations can exceed, equal, or be lower than unbound plasma concentrations. Disconnection between hepatocyte and plasma concentrations of various endogenous or exogenous substrates becomes even more unpredictable when the expression and functions of membrane transporters are altered during liver diseases [
1,
2]. With advances in imaging, the quantification of liver concentrations is now possible following the injection of hepatobiliary substrates [
3]. Using pharmacokinetic modelling, liver imaging can also predict the function of influx and efflux transporters of hepatocytes [
4].
Most patients with liver metastases from colorectal cancer receive chemotherapy before liver resection. This presurgical chemotherapy may induce liver injuries such as steatosis, nodular regenerative hyperplasia, steatohepatitis, and sinusoidal obstruction syndrome (SOS) [
5]. This chemotherapy-associated liver injury increases the number of complications following hepatectomy. Post-chemotherapy SOS is characterised by the loss of sinusoidal wall integrity and subsequent sinusoidal congestive obstruction. SOS regresses only months after the end of chemotherapy [
6]. The syndrome has been associated with more than 20 drugs, including conventional immunosuppressive and chemotherapeutic agents such as oxaliplatin.
Chemotherapy in rodents does not induce SOS, but the syndrome is observed after monocrotaline (MCT) gavage [
7]. MCT is a pyrrolizidine alkaloid metabolised into the toxic compound monocrotaline pyrrole by cytochrome P450 enzymes present in sinusoidal endothelial cells, inducing sinusoidal injury. In this model, we previously showed that sinusoidal obstruction persists 18 days after MCT gavage in 25% of rat livers [
8]. Following a 70% hepatectomy, liver regeneration was significantly impaired, and this impaired regeneration was associated with hepatocellular injury. The biliary excretion rates of the imaging substrate gadobenate dimeglumine (BOPTA) was significantly decreased.
BOPTA is a hepatobiliary substrate able to characterise human liver function and focal lesions when injected during magnetic resonance imaging (MRI) (
Figure 1D) [
9,
10]. This characterisation relies on BOPTA transport across hepatocyte membrane proteins. We previously showed in cultured oocytes that BOPTA is a substrate of the rat organic anion-transporting polypeptides (Oatps) (
Figure 1A) [
11]. BOPTA is not excreted in the bile canaliculi of rats lacking the multidrug-resistance-associated protein 2 (Mrp2) [
12]. We also showed that BOPTA is excreted back into sinusoids, but we did not identify the responsible membrane transporters. Mrp3/4 transporters might be involved as published with another imaging substrate, gadoxetate dimeglumine [
13]. In the present study, we characterised how the alterations of transporter functions in injured hepatocytes explain the decreased biliary excretion of BOPTA associated with SOS.
3. Discussion
This study evidences a dysfunction of efflux transporters associated with SOS. Other diseases such as liver inflammation, fibrosis, and cirrhosis were previously associated with transporter dysfunction [
1,
2]. The function of OATPs and MRP2 were altered in human nonalcoholic steatohepatitis, leading to a decreased influx clearance of
99mTc[Mebrofenin] [
15]. The biliary clearance of
99mTc[Mebrofenin] was also decreased. The authors anticipated that the pharmacokinetics of drugs transported by OATP1B1/1B3 and MRP2 are likely to have similar abnormalities. SOS induced by pyrrolizidine alkaloids is also associated with biologic markers of cholestasis. High concentrations of total bilirubin were measured following the administration of the pyrrolizidine alkaloid senecionine in rats [
16]. Conjugated bilirubin and BOPTA are both transported across hepatocytes by Oatps, Mrp2, and Mrp3/4 transporters [
17]. Hyperbilirubinemia is present in patients with SOS [
18]. In accordance with our results, hepatic Mrp3 mRNA expression was 13 times higher in senecionine-treated than in normal rats [
16]. In contrast, Oatps mRNA expression was significantly decreased in senecionine-treated rats.
As previously published [
8], BOPTA bile excretion is altered in livers 18 days after the induction of SOS. Before BOPTA perfusion, bile flow rates in Nl and MCT livers were not significantly different, but BOPTA-induced bile flow increases (choleresis) and BOPTA biliary excretion were importantly reduced in MCT livers during the perfusion period. Similar results were published in mice injected with senecionine [
19]. In this study, intravital two-photon imaging showed a delayed biliary excretion rate of fluorescent bile salt analogues. In the present study, we characterised how the alterations of transporter functions in injured hepatocytes explain the decreased biliary excretion of BOPTA associated with SOS.
Besides BOPTA, we perfused the extracellular substrate DTPA to estimate BOPTA extracellular concentrations. BOPTA distribution into the extracellular space was similar in both groups. This result confirmed the absence of fibrosis, inflammation, or hepatocyte necrosis observed in biopsies collected from MCT livers 18 days after SOS induction [
8]. Following extracellular distribution, BOPTA enters hepatocytes by Oatps located on the basolateral membrane [
11]. The function of Oatps was not modified by MCT according to the similar influx clearances (CL
in) measured in both groups. However, a case report found a decreased expression of OATP1B3 in a patient with SOS [
20]. In the study, the reduced expression was associated with a decreased liver accumulation of another hepatobiliary MRI substrate Gadoxetate. The identical CL
in was not associated with a similar early accumulation within hepatocytes (MCT livers had a lower L
1 slope than Nl livers). One explanation might be that BOPTA sinusoidal distribution is altered in MCT livers, impairing early BOPTA uptake. Moreover, CL
in was measured at the end of the perfusion period when hepatocyte concentrations were maximal.
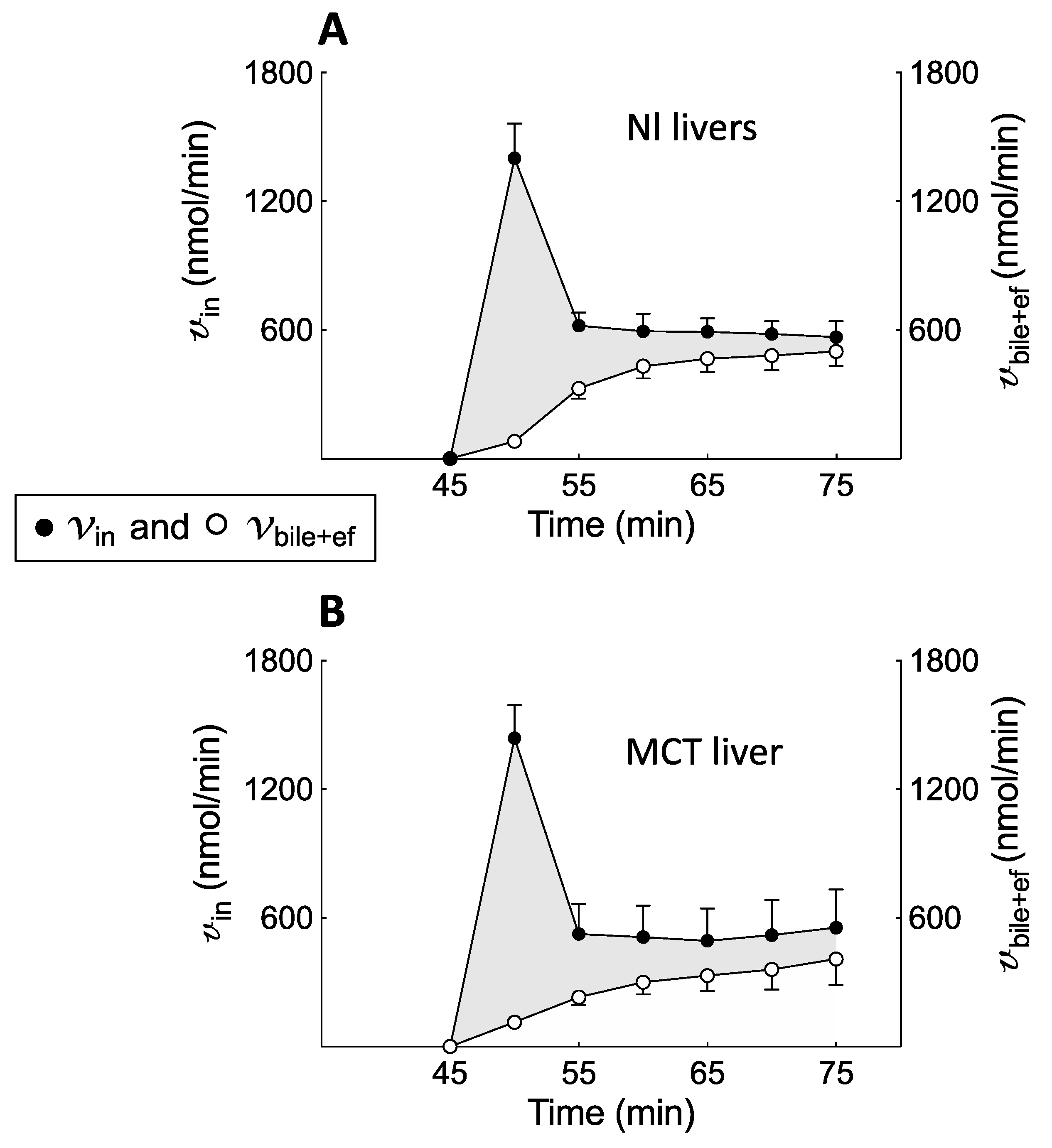
BOPTA biliary clearance (CL
bile) and BOPTA bile concentrations were much lower in MCT livers, and the choleretic effect of BOPTA was nearly abolished. Mrp2 function is linked to that of aquaporin 8 on cholesterol-enriched domains in the canalicular membrane [
21]. However, the mechanism of this link was never investigated. BOPTA efflux clearance back to sinusoids (CL
ef) was higher in MCT livers and compensated for the low biliary excretion. The total efflux clearance (CL
bile+ef) was similar in both groups. Thus, Mrp2 function is highly impaired, while that of Mrp3/4 is proportionally increased in MCT livers. Such compensation of CL
ef was also observed in livers with steatosis [
22]. Similar findings were published with bilirubin [
17], morphine [
23], and endogenous substrates such as coproporphyrins [
24]. Because influx (CL
in) and efflux (CL
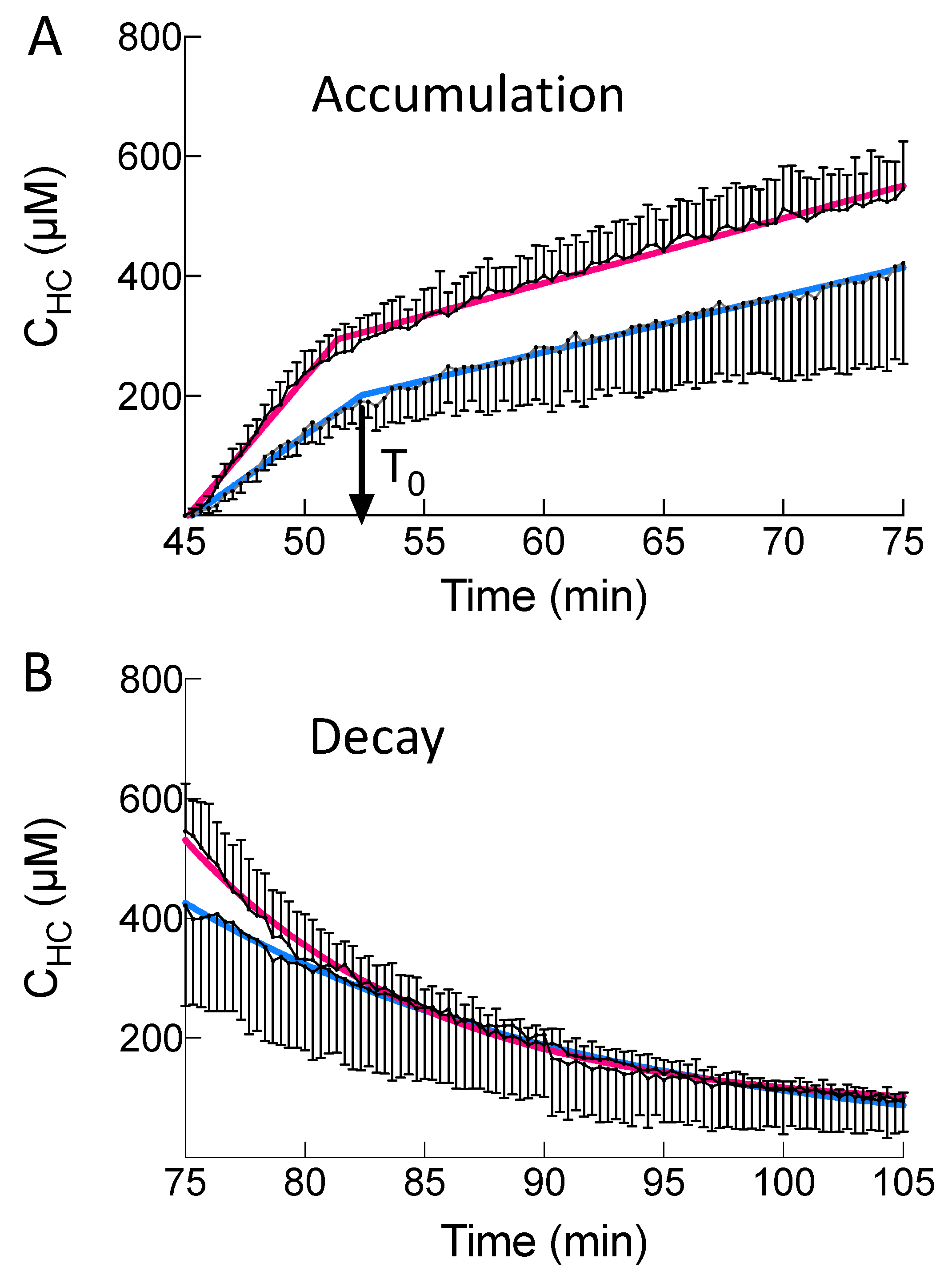
bile+ef) clearances were similar in Nl and MCT livers, the BOPTA concentration profile in hepatocytes was not significantly different during the perfusion and rinse periods. This cooperation between Mrp2 and Mrp3/4 functions seems useful to maintain normal hepatocyte concentrations. The mechanism that triggers such cooperation was never investigated. In the absence of such cooperation, cholestasis would increase hepatocyte concentrations when Oatp function was maintained.
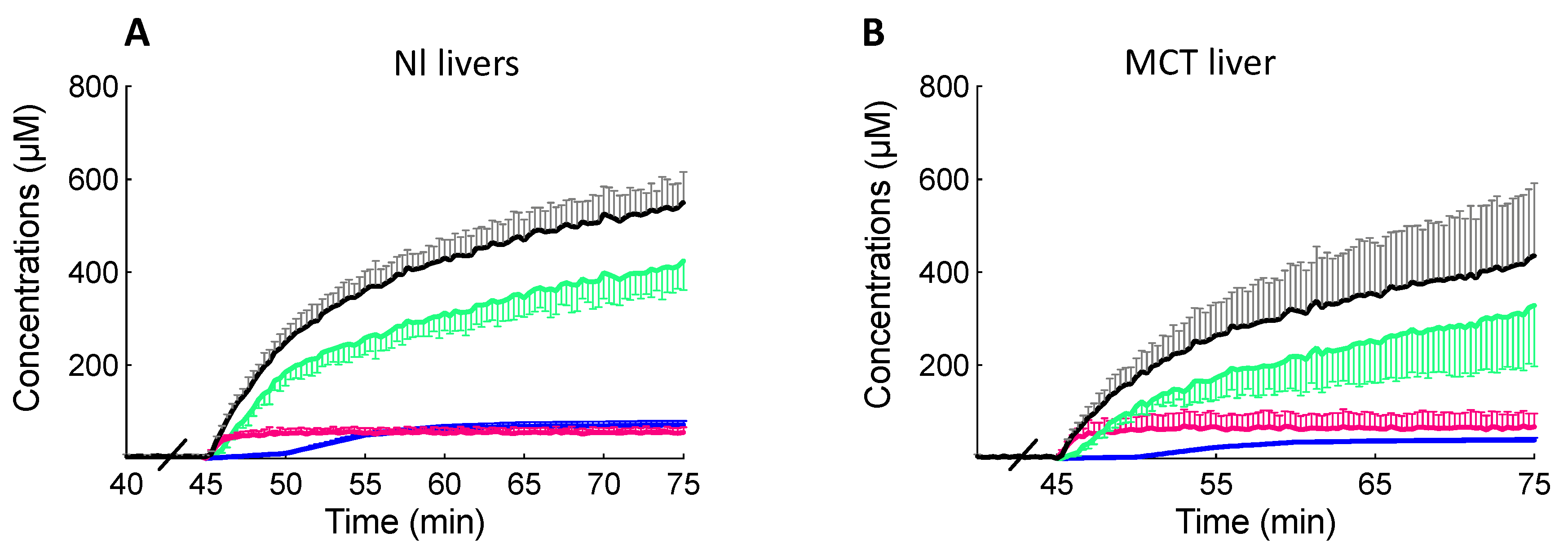
However, the high BOPTA efflux clearance back to the sinusoids altered BOPTA elimination from MCT livers. Concentrations in the hepatic veins (Cout) were higher in MCT than in Nl livers because these concentrations include BOPTA concentrations that did not enter the livers plus BOPTA concentrations that had entered the liver but returned to the sinusoids. Liver extraction ratio, sinusoidal removal rates (v), and hepatic clearance (CLH) were lower in MCT livers. By extrapolating this finding to the in vivo situation, plasma concentrations would remain higher in MCT than in Nl rats. According to this finding, all substrates including drugs transported by Oatps, Mrp2, and Mrp3/4 would have a decreased plasma concentration decay.
The isolated and perfused rat liver is a convenient model because the experimental conditions are well controlled and simplified. The experimental protocol was designed to measure various parameters during the accumulation and rinse periods. Livers are perfused only through the portal vein with a constant 200 µM concentration of BOPTA. There is no recirculation of solutions, which are discarded after the first pass through the sinusoids. This approach is useful to measure the liver extraction ratio (Cin − Cout)/Cin. To simplify the protocol, we did not add proteins into the KHB solution, and BOPTA was free to enter the hepatocytes.
The experimental model has limitations. In humans, SOS is associated with more than 20 chemotherapeutic drugs which have no effect on rat livers. However, the rodent syndrome can be induced by pyrrolizidine alkaloids [
7,
25]. Extrapolation of the results to other species and humans must be cautious because the expression of membrane transporters can differ. We did not measure the expression of transporters but quantified the concomitant functions of three transporters and their resulting effects on hepatocyte concentrations. The experimental model which quantifies all compartmental concentrations, transfer rates, and clearances associated with MCT administration is unique. We improved it over the past decades. The model is adequate for quantifying the changes of BOPTA hepatocyte concentrations associated with various flow rates and drug–drug interactions. Other limitations are linked to the assumptions needed to obtain several pharmacokinetic parameters. BOPTA extracellular concentrations cannot be measured because the substrate enters the hepatocytes within 2 min. To assess these concentrations, we used another MRI substrate, DTPA, which has a similar chemical formula and is distributed only in the extracellular spaces of the liver. Moreover, we assumed that concentrations inside the bile canaliculi were similar to those measured in the common bile duct, although solute export from the cholangiocytes and water transport along the ductules and ducts may modify the primary bile in the canaliculi. This issue was never investigated and has been recently debated by the authors [
26]. Another limitation is the ratio between the liver compartments and the liver. The volume ratio of bile canaliculi and liver was previously estimated by Blouin et al. [
27] at 0.43% on normal biopsies in rats. In the same article, the volume ratio of hepatocytes and liver was 78%. Because we previously published that fibrosis, inflammation and hepatocyte necrosis disappeared 18 days after MCT administration, we used the same ratios in Nl and MCT livers. CL
bile was similar during the accumulation and rinse periods (data not shown), and we assumed that it was also true for CL
ef during both periods. Then, we were able to estimate C
ef or BOPTA concentrations that returned into the sinusoids by (C
HC × CL
ef)/Q
H 4. Materials and Methods
4.1. Induction of the Sinusoidal Obstruction Syndrome (SOS)
Male Sprague–Dawley rats (175–250 g, n = 4) had an intragastric administration of MCT (160 mg/kg, Sigma, Buchs, Switzerland) while 5 additional normal (Nl) rats had an intragastric administration of saline solution [
7,
28]. MCT livers were isolated and perfused 18 days after gavage. The protocol was carried out in accordance with the Swiss Guidelines for the Care and Use of Laboratory Animals and was approved by the local animal welfare committee and the veterinary office in Geneva, Switzerland (N° 1006.3384.2).
4.2. Isolated and Perfused Rat Livers
Before liver isolation, normal (Nl) rats and rats that received MCT were anesthetised with pentobarbital (50 mg kg
−1, ip). In these two groups, we published preliminary data on the transport of BOPTA across the hepatocyte membranes [
8]. In the present study, we returned to the raw data to thoroughly analyse new pharmacokinetic parameters as recently published [
29]. Rat livers were isolated, leaving the liver in the carcass. The abdominal cavity was opened, and the portal vein cannulated. The hepatic artery was not perfused. The abdominal vena cava was transected, and an oxygenated Krebs–Henseleit bicarbonate (KHB) solution was pumped into the portal vein, the solution being discarded after liver distribution via vena cava transection. The flow rate was slowly increased over 1 min up to 30 mL/min. In a second step, the chest was opened, and a cannula was inserted through the right atrium to collect solutions flowing from the hepatic veins. Finally, the abdominal inferior vena cava was ligated, allowing solutions perfused by the portal vein to be eliminated by the hepatic veins.
The perfusion system included a reservoir, a pump, a heating circulator, a bubble trap, a filter, and an oxygenator. Solutions of perfusion were equilibrated with a mixture of 95% O
2 and 5% CO
2. Livers were continuously perfused with fresh solutions using a nonrecirculating system. The common bile duct was cannulated with a PE
10 catheter, and bile samples were collected every 5 min to measure bile flow rates (Q
bile, µL/min per liver) and BOPTA concentrations (C
bile, µM). BOPTA-increased bile flow rates were measured by the area under the curve of Q
bile increases over time (AUCQ
bile, µL). Samples were collected from hepatic veins every 5 min to measure BOPTA concentrations (C
out, µM). C
out during the BOPTA perfusion period were concentrations that did not enter into the hepatocytes plus concentrations that entered into the hepatocytes and returned to the sinusoids (
Figure 1A). During the rinse period, C
out were concentrations leaving the hepatocytes because no BOPTA was perfused. To estimate concentrations leaving the hepatocytes during the perfusion period, we used the equation (C
HC • CL
ef)/Q
H (see later sections).
A gamma counter was placed over the liver to detect BOPTA concentrations in a single region of interest that must be representative of the entire liver. Concentrations in the region of interest averaged out the concentrations of numerous hepatocytes, knowing that Oatps are mainly expressed in perivenous hepatocytes where BOPTA is likely to enter.
4.3. Perfusion of DTPA and BOPTA
Rat livers were perfused with gadopentetate dimeglumine (DTPA; Magnevist
®; Bayer imaging) and gadobenate dimeglumine (BOPTA, MultiHance
®; Bracco Imaging). DTPA distributes only within the sinusoids and interstitium, while BOPTA distributes into the extracellular space, hepatocytes, and bile canaliculi. DTPA and BOPTA labelled with
153Gd were obtained by adding
153GdCl
3 (1 MBq/mL) to the commercially available (0.5 M) solutions of DTPA and BOPTA. Then, [
153Gd]DTPA and [
153Gd]BOPTA were diluted in the KHB solution to obtain a 200 µM concentration. Livers were successively perfused with 200 µM [
153Gd]DTPA (10 min), KHB solution (35 min), 200 µM [
153Gd]BOPTA (30 min, accumulation or perfusion period), and KHB solution (30 min, decay or elimination period) (
Figure 1B). The protocol lasted 105 min for each group. BOPTA extracellular concentrations cannot be measured because the substrate enters within 2 min into the hepatocytes. To assess these concentrations, we used another MRI substrate DTPA which has a similar chemical formula and distributes only in the extracellular space of the livers. BOPTA and DTPA are widely used in human imaging to characterise BOPTA extracellular and hepatocyte accumulation.
4.4. BOPTA Concentrations in Liver Compartments
To quantify BOPTA concentrations in the liver compartments, a gamma counter that collects count rates every 20 s was placed 1 cm above the right liver lobe. The counter measured the radioactivity in a region of interest that was identical in each rat liver. To transform count rates into BOPTA concentrations, the total liver radioactivity was measured by an activimeter at the end of each experiment and was related to the last count rates. Radioactivity was corrected for decay.
The gamma counter delineated a region of interest in the liver lobe from which all count rates originating from the extracellular space, hepatocytes, and bile canaliculi were divided by the liver weight to obtain liver concentrations (C
liver, µM). Concentrations in the extracellular space were measured during DTPA pre-perfusion (C
EC, µM). C
EC was constant during the 10 min perfusion. We assumed that concentrations inside the bile canaliculi were similar to those measured in the common bile duct (C
bile, µM), although solute export from the cholangiocytes and water transport along the ductules and ducts might have modified the primary bile in the canaliculi. This issue was recently debated by the authors [
26]. The volume ratio of the bile canaliculi and liver was previously estimated by Blouin et al. [
27] at 0.43%, and BOPTA concentrations in the bile canaliculi detected by the counter (C
BC) were 0.0043 × C
bile. Hepatocyte concentrations (C
HC78%) detected by the counter were C
liver − C
EC − 0.0043 C
bile. Blouin et al. [
27] previously determined that the volume ratio of hepatocytes in livers without fibrosis or inflammation was 78%. In situ BOPTA hepatocyte concentrations (C
HC100%) were calculated by (100/78) × C
HC78%.
4.5. BOPTA Transfer Rates and Clearances between Compartments
BOPTA removal rates from sinusoids during the perfusion period (v, nmol/min) were measured by QH × (Cin − Cout), where QH is the constant liver flow rate (30 mL/min), Cin (µM) is the constant portal concentration, and Cout (µM) is the concentration measured in the hepatic veins. The unbound fraction in the solutions was 1 because no protein was added into the solutions. Hepatic clearance (CLH, mL/min) was the ratio of v and Cin during the last min of perfusion. The BOPTA extraction ratio (ER) was (Cin − Cout)/Cin. The BOPTA biliary excretion rate (vbile, nmol/min) was Cbile × Qbile, where Cbile (µM) was the concentration in the common bile duct and Qbile was the bile flow rate (µL/min per liver). Clearance from the hepatocytes to the bile canaliculi (CLbile, mlHC/min) was the slope of linear regression between vbile (Y-axis) and the hepatocyte concentrations (CHC, X-axis), which was measured during the entire protocol. This clearance was expressed in ml of hepatocytes (mlHC) per min.
During the rinse period, BOPTA concentrations leaving the hepatocytes (Cef, µM) into the sinusoids were measured by Cout because no BOPTA was perfused in the portal vein. Basolateral efflux from the hepatocytes into the sinusoids (vef, nmol/min) was Cef × QH, and basolateral clearance (CLef, mlHC/min) was the slope of linear regression between vef (Y-axis) and CHC (X-axis). With the assumption that CLef was similar during both perfusion and rinse periods, we estimated Cef during the accumulation period by (CHC × CLef)/QH. The hepatocyte influx rate vin (nmol/min) was [Cin − (Cout − Cef )] × QH or v + vef. In other words, BOPTA entry into the hepatocytes includes BOPTA eliminated into the bile canaliculi and back into the sinusoids. Hepatocyte influx clearance CLin was vin/Cin. CLin was measured during the last min of perfusion and is expressed in ml of Krebs–Henseleit bicarbonate solution per min (mlKBH/min)
4.6. Accumulation Profile of Hepatocyte Concentrations
During the perfusion period, BOPTA hepatocyte accumulation was best described by a segmental linear regression obtained from GraphPad Prism version 8, GraphPad Software, La Jolla, CA, USA [
14]. This function defines a first line L
1 for time below T
0 and a second line L
2 for time higher than T
0. T
0 is the time when BOPTA efflux from the hepatocytes decreases BOPTA accumulation into the hepatocyte. No constraint was applied to fit the data.
4.7. Decay Profile of Hepatocyte Concentrations
During the decay period, the data were best described by a one-phase decay (GraphPad Prism version 8, GraphPad Software, La Jolla, CA, USA) [
30]. No constraint was applied to fit the data. The model was defined by a rate constant of elimination (k
el,HC, min
−1). Knowing CL
bile+ef and k
el,HC, we calculated the apparent hepatocyte volumes (V
HC, mL) as CL
bile+ef/k
el,HC.
4.8. Statistics
Data are means ± SD. Parameters obtained in Nl and MCT livers were compared with a Mann–Whitney test (GraphPad Prism version 8, GraphPad Software, La Jolla, CA, USA). To compare over time the effect of treatment (Nl vs. MCT) on additional parameters, we used a two-way ANOVA with Sidak multiple comparison tests. p < 0.05 was considered statistically significant.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}