
Yohimbine, an α2-Adrenoceptor Antagonist, Suppresses PDGF-BB-Stimulated Vascular Smooth Muscle Cell Proliferation by Downregulating the PLCγ1 Signaling Pathway

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of YOH on PDGF-BB-Induced Proliferation and Cell Cycle Progression in MOVAS-1 Cells
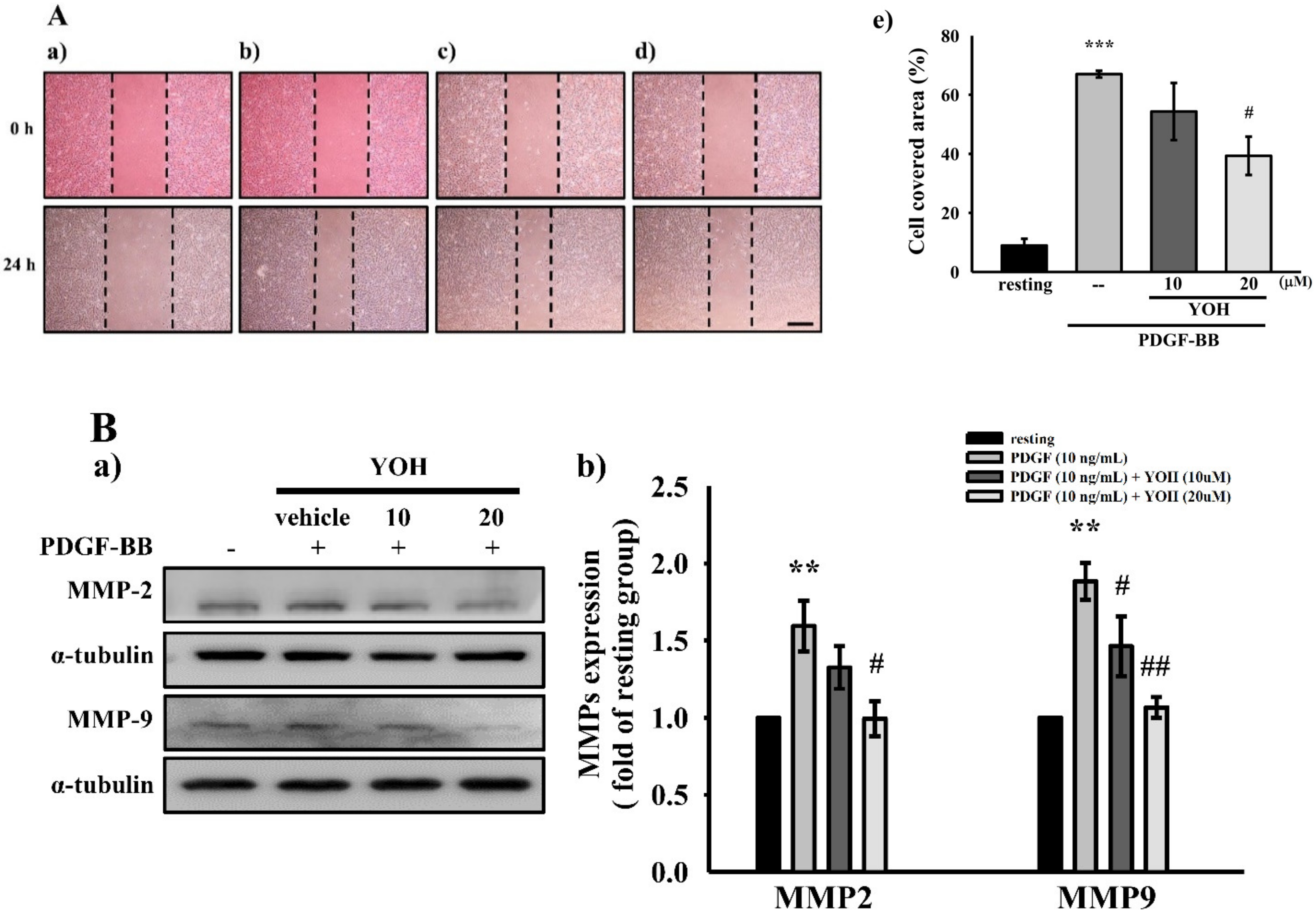
2.2. YOH Inhibits PDGF-BB-Stimulated Migration as Well as MMP-2 and MMP-9 Expression in MOVAS-1 Cells
2.3. YOH Modulates the Expression of Cell-Cycle-Associated Proteins in VSMCs Treated with PDGF-BB
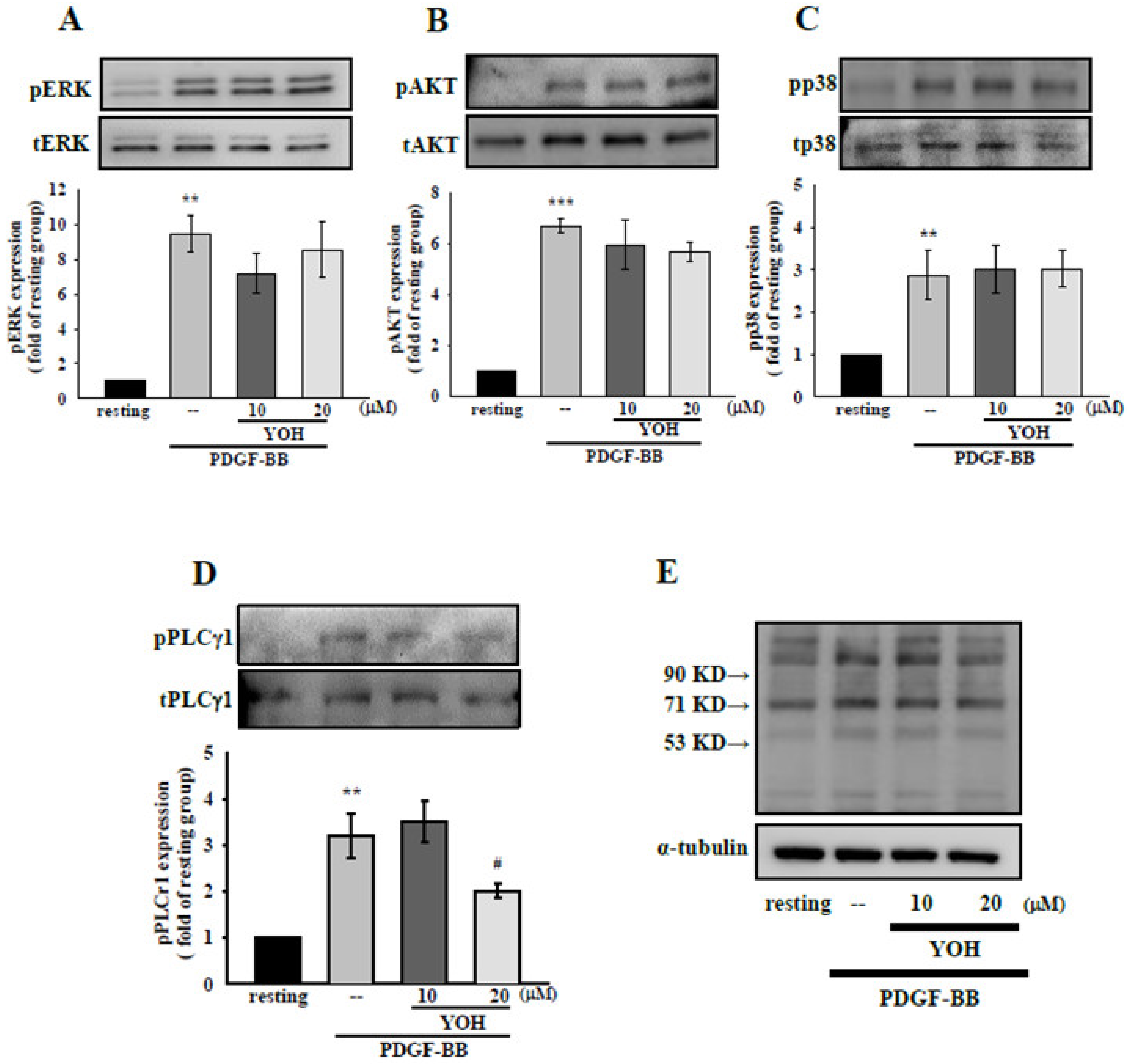
2.4. Mechanisms Underlying the Inhibitory Effects of YOH on PLCγ1 Phosphorylation in PDGF-BB-Treated MOVAS-1 Cells
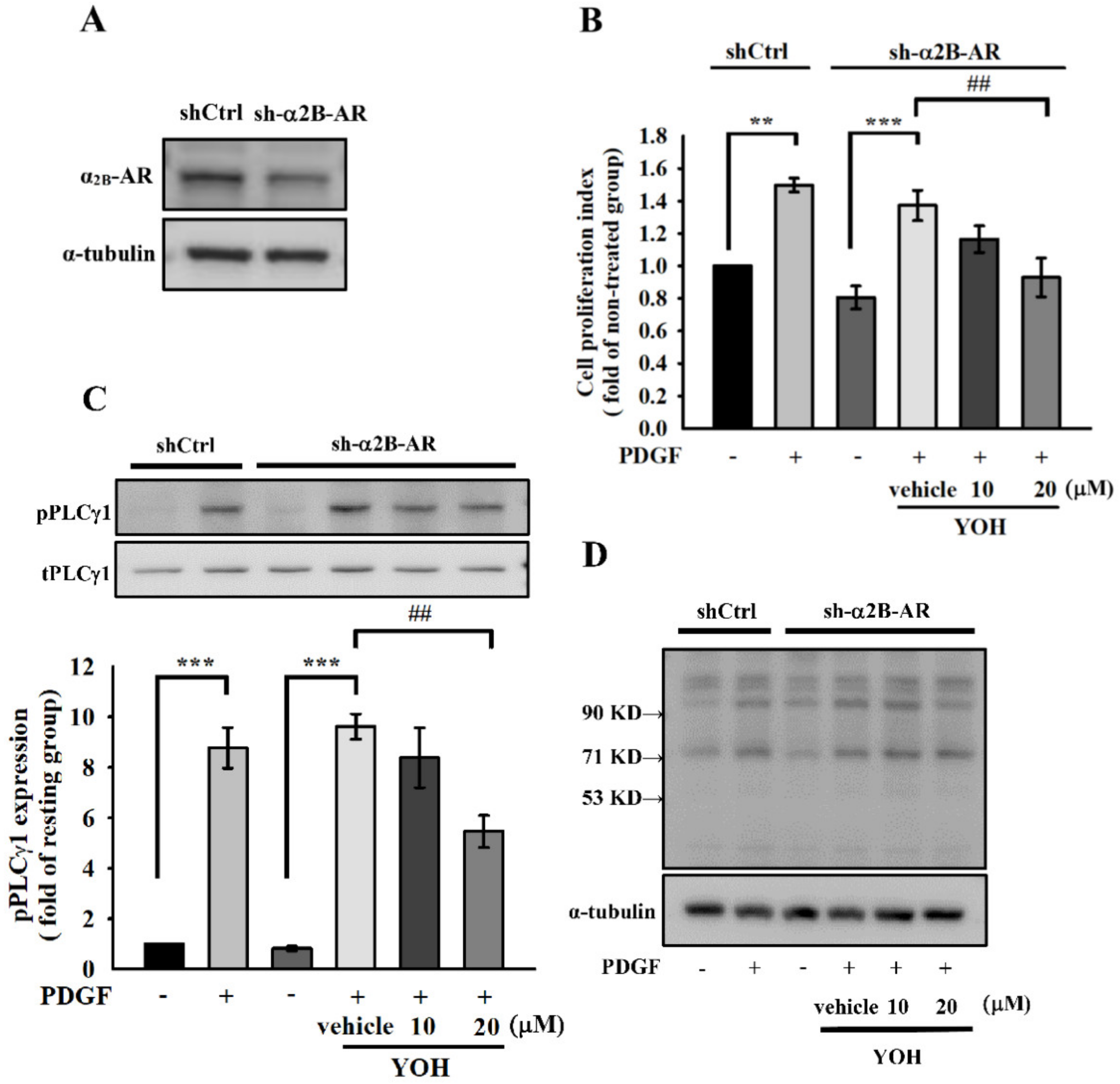
2.5. Downregulation of Cell Proliferation and PLCγ1 Phosphorylation by YOH Is Independent of Its Known α2-Antagonism in MOVAS-1 Cells Stimulated by PDGF-BB
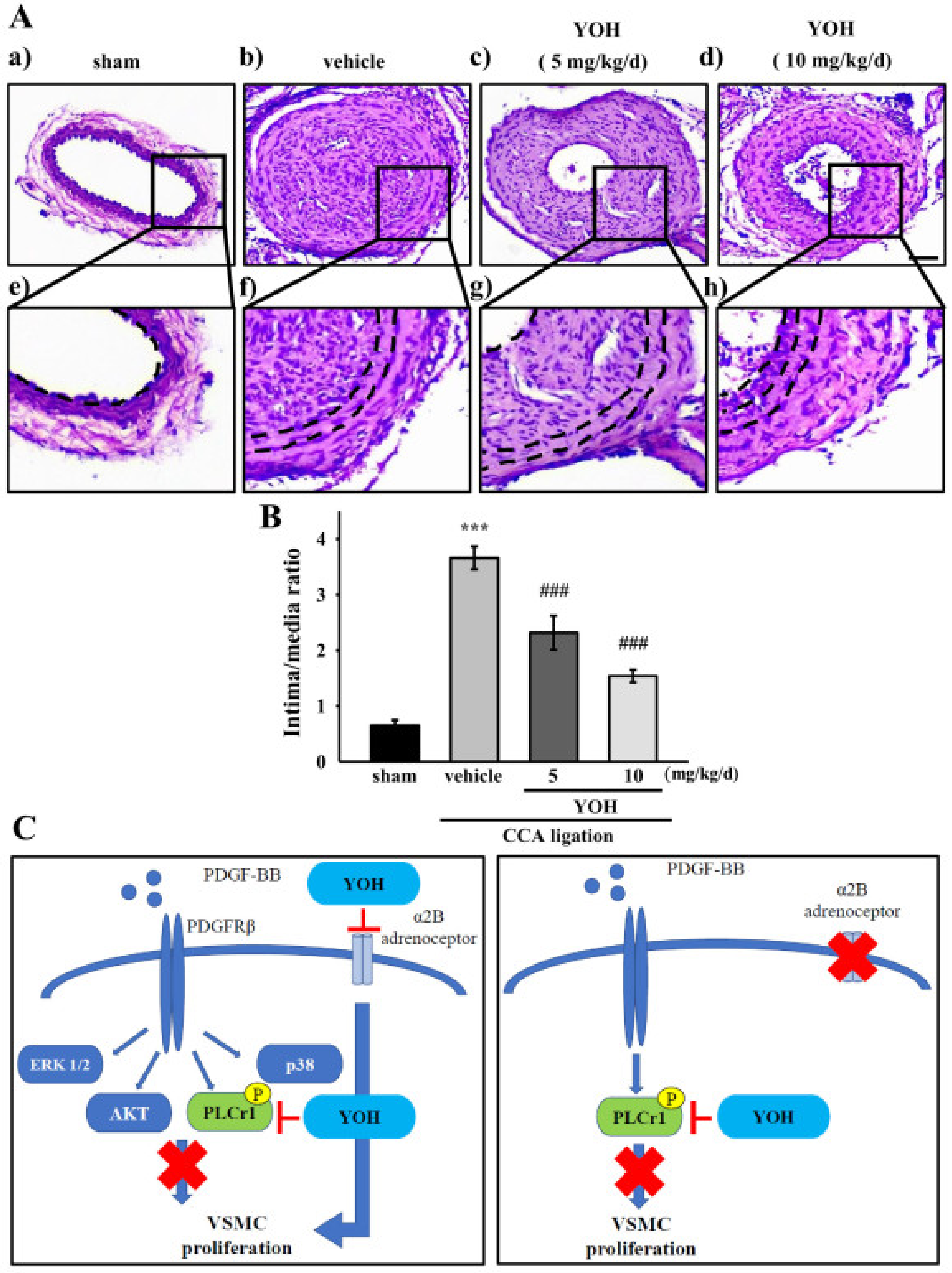
2.6. YOH Suppresses Neointimal Hyperplasia in a Mouse Model of CCA Ligation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. VSMC Cell Culture and Lentivirus Infection
4.3. Proliferation Assay (MTT Assay)
4.4. Wound Healing Assay
4.5. Cell Cycle Analysis
4.6. Western Blotting
4.7. Mouse Model of Common Carotid Artery Ligation Model
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hsieh, C.Y.; Liu, C.L.; Hsu, M.J.; Jayakumar, T.; Chou, D.S.; Wang, Y.H.; Hsiao, G.; Sheu, J.R. Inhibition of vascular smooth muscle cell proliferation by the vitamin E derivative pentamethylhydroxychromane in an in vitro and in vivo study: Pivotal role of hydroxyl radical-mediated PLCgamma1 and JAK2 phosphorylation. Free Radic. Biol. Med. 2010, 49, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, M.J.; Li, B.H.; Chen, L.; Pi, Y.; Yin, Y.W.; Long, C.Y.; Wang, X.; Sun, M.J.; Chen, X.; et al. PPARγ Inhibits VSMC Proliferation and Migration via Attenuating Oxidative Stress through Upregulating UCP2. PLoS ONE 2016, 11, e0154720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, P.; Jian, W.; Xu, X.; Kou, W.; Yu, Q.; Hu, H.; Li, D.; Wang, W.; Feinberg, M.W.; Zhuang, J.; et al. NLRC5 inhibits neointima formation following vascular injury and directly interacts with PPARγ. Nat. Commun. 2019, 10, 2882. [Google Scholar] [CrossRef] [PubMed]
- Jawien, A.; Bowen-Pope, D.F.; Lindner, V.; Schwartz, S.M.; Clowes, A.W. Platelet-derived growth factor promotes smooth muscle migration and intimal thickening in a rat model of balloon angioplasty. J. Clin. Investig. 1992, 89, 507–511. [Google Scholar] [CrossRef]
- Dzau, V.J.; Braun-Dullaeus, R.C.; Sedding, D.G. Vascular proliferation and atherosclerosis: New perspectives and therapeutic strategies. Nat. Med. 2002, 8, 1249–1256. [Google Scholar] [CrossRef]
- Lee, S.J.; Won, S.Y.; Park, S.L.; Song, J.H.; Noh, D.H.; Kim, H.M.; Yin, C.S.; Kim, W.J.; Moon, S.K. Rosa hybrida extract suppresses vascular smooth muscle cell responses by the targeting of signaling pathways, cell cycle regulation and matrix metalloproteinase-9 expression. Int. J. Mol. Med. 2016, 37, 1119–1126. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.H.; Westermark, B. Mechanism of action and in vivo role of platelet-derived growth factor. Physiol. Rev. 1999, 79, 1283–1316. [Google Scholar] [CrossRef]
- Uzui, H.; Lee, J.D.; Shimizu, H.; Tsutani, H.; Ueda, T. The role of protein-tyrosine phosphorylation and gelatinase production in the migration and proliferation of smooth muscle cells. Atherosclerosis 2000, 149, 51–59. [Google Scholar] [CrossRef]
- Jiang, D.; Zhuang, J.; Peng, W.; Lu, Y.; Liu, H.; Zhao, Q.; Chi, C.; Li, X.; Zhu, G.; Xu, X.; et al. Phospholipase Cγ1 Mediates Intima Formation Through Akt-Notch1 Signaling Independent of the Phospholipase Activity. J. Am. Heart Assoc. 2017, 6, e005537. [Google Scholar] [CrossRef]
- Caglayan, E.; Vantler, M.; Leppänen, O.; Gerhardt, F.; Mustafov, L.; Ten Freyhaus, H.; Kappert, K.; Odenthal, M.; Zimmermann, W.H.; Tallquist, M.D.; et al. Disruption of platelet-derived growth factor-dependent phosphatidylinositol 3-kinase and phospholipase Cγ 1 activity abolishes vascular smooth muscle cell proliferation and migration and attenuates neointima formation in vivo. J. Am. Coll. Cardiol. 2011, 57, 2527–2538. [Google Scholar] [CrossRef] [Green Version]
- Porst, H.; Burnett, A.; Brock, G.; Ghanem, H.; Giuliano, F.; Glina, S.; Hellstrom, W.; Martin-Morales, A.; Salonia, A.; Sharlip, I. SOP conservative (medical and mechanical) treatment of erectile dysfunction. J. Sex Med. 2013, 10, 130–171. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Shang, B.; Wang, X.; Chen, Q. PEEK tube-based online solid-phase microextraction-high-performance liquid chromatography for the determination of yohimbine in rat plasma and its application in pharmacokinetics study. Biomed. Chromatogr. 2017, 31, 3866. [Google Scholar] [CrossRef] [PubMed]
- Neha; Ansari, M.M.; Khan, H.A. Yohimbine hydrochloride ameliorates collagen type-II-induced arthritis targeting oxidative stress and inflammatory cytokines in Wistar rats. Environ. Toxicol. 2017, 32, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Ou, F.; Huang, Y.; Sun, J.; Su, K.; He, Y.; Zeng, R.; Tang, D.; Liao, G. Yohimbine Ameliorates Temporomandibular Joint Chondrocyte Inflammation with Suppression of NF-κB Pathway. Inflammation 2021, 44, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Le Dorze, C.; Gisquet-Verrier, P. Sensitivity to trauma-associated cues is restricted to vulnerable traumatized rats and reinstated after extinction by yohimbine. Behav. Brain Res. 2016, 313, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, S.; Tobassum, S.; Farooq, U.; Malik, H.; İdris, M. Effects of naloxone and yohimbine in polycystic ovary syndrome: A rabbit model study. Turk. J. Med. Sci. 2016, 46, 1265–1270. [Google Scholar] [CrossRef]
- Shimokawa, T.; Yoneda, K.; Yamagata, M.; Hayashi, K.; Tomita, S. Yohimbine ameliorates lipopolysaccharide-induced acute kidney injury in rats. Eur. J. Pharmacol. 2020, 871, 172917. [Google Scholar] [CrossRef]
- Xia, M.; Tong, J.H.; Zhou, Z.Q.; Duan, M.L.; Xu, J.G.; Zeng, H.J.; Wang, S.H. Tramadol inhibits proliferation, migration and invasion via a2-adrenoceptor signaling in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 157–165. [Google Scholar]
- Shen, S.G.; Zhang, D.; Hu, H.T.; Li, J.H.; Wang, Z.; Ma, Q.Y. Effects of alpha-adrenoreceptor antagonists on apoptosis and proliferation of pancreatic cancer cells in vitro. World J. Gastroenterol. 2008, 14, 2358–2363. [Google Scholar] [CrossRef]
- Huhtinen, A.; Scheinin, M. Expression and characterization of the human alpha 2B-adrenoceptor in a vascular smooth muscle cell line. Eur. J. Pharmacol. 2008, 587, 48–56. [Google Scholar] [CrossRef]
- Handy, D.E.; Johns, C.; Bresnahan, M.R.; Tavares, A.; Bursztyn, M.; Gavras, H. Expression of alpha2-adrenergic receptors in normal and atherosclerotic rabbit aorta. Hypertension 1998, 32, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, R.E.; Desai, K.; Hein, L.; Stevens, M.E.; Chruscinski, A.; Bernstein, D.; Barsh, G.S.; Kobilka, B.K. Cardiovascular regulation in mice lacking alpha2-adrenergic receptor subtypes b and c. Science 1996, 273, 803–805. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.D.; Trendelenburg, A.U.; MacMillan, L.; Bernstein, D.; Limbird, L.; Starke, K.; Kobilka, B.K.; Hein, L. Abnormal regulation of the sympathetic nervous system in alpha2A-adrenergic receptor knockout mice. Mol. Pharmacol. 1999, 56, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Chotani, M.A.; Mitra, S.; Eid, A.H.; Han, S.A.; Flavahan, N.A. Distinct cAMP signaling pathways differentially regulate alpha2C-adrenoceptor expression: Role in serum induction in human arteriolar smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H69–H76. [Google Scholar] [CrossRef] [Green Version]
- Huhtinen, A.; Hongisto, V.; Laiho, A.; Löyttyniemi, E.; Pijnenburg, D.; Scheinin, M. Gene expression profiles and signaling mechanisms in α(2B)-adrenoceptor-evoked proliferation of vascular smooth muscle cells. BMC Syst. Biol. 2017, 11, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Song, I.S.; Jeong, Y.J.; Park, J.H.; Shim, S.; Jang, S.W. Chebulinic acid inhibits smooth muscle cell migration by suppressing PDGF-Rβ phosphorylation and inhibiting matrix metalloproteinase-2 expression. Sci. Rep. 2017, 7, 11797. [Google Scholar] [CrossRef]
- Belo, V.A.; Guimarães, D.A.; Castro, M.M. Matrix Metalloproteinase 2 as a Potential Mediator of Vascular Smooth Muscle Cell Migration and Chronic Vascular Remodeling in Hypertension. J. Vasc. Res. 2015, 52, 221–231. [Google Scholar] [CrossRef]
- Johnson, C.; Galis, Z.S. Matrix metalloproteinase-2 and -9 differentially regulate smooth muscle cell migration and cell-mediated collagen organization. Arter. Thromb. Vasc. Biol. 2004, 24, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.S.; Wang, M.J.; Jayakumar, T.; Chou, D.S.; Ko, C.Y.; Hsu, M.J.; Hsieh, C.Y. Antiproliferative Activity of Hinokitiol, a Tropolone Derivative, Is Mediated via the Inductions of p-JNK and p-PLCg1 Signaling in PDGF-BB-Stimulated Vascular Smooth Muscle Cells. Molecules 2015, 20, 8198–8212. [Google Scholar] [CrossRef] [Green Version]
- Philipp, M.; Brede, M.; Hein, L. Physiological significance of alpha(2)-adrenergic receptor subtype diversity: One receptor is not enough. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R287–R295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, A.W.; Tulis, D.A. Experimental Rat and Mouse Carotid Artery Surgery: Injury & Remodeling Studies. ISRN Minim. Invasive Surg. 2013, 2013, 167407. [Google Scholar] [PubMed]
- Dudek, M.; Knutelska, J.; Bednarski, M.; Nowiński, L.; Zygmunt, M.; Mordyl, B.; Głuch-Lutwin, M.; Kazek, G.; Sapa, J.; Pytka, K. A Comparison of the Anorectic Effect and Safety of the Alpha2-Adrenoceptor Ligands Guanfacine and Yohimbine in Rats with Diet-Induced Obesity. PLoS ONE 2015, 10, e0141327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenway, F.L.; Bray, G.A.; Heber, D. Topical fat reduction. Obes. Res. 1995, 3, 561s–568s. [Google Scholar] [CrossRef] [PubMed]
- Worssam, M.D.; Jørgensen, H.F. Mechanisms of vascular smooth muscle cell investment and phenotypic diversification in vascular diseases. Biochem. Soc. Trans. 2021, 49, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Lacolley, P.; Regnault, V.; Nicoletti, A.; Li, Z.; Michel, J.B. The vascular smooth muscle cell in arterial pathology: A cell that can take on multiple roles. Cardiovasc. Res. 2012, 95, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular Smooth Muscle Cells in Atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef]
- Newby, A.C. Matrix metalloproteinases regulate migration, proliferation, and death of vascular smooth muscle cells by degrading matrix and non-matrix substrates. Cardiovasc. Res. 2006, 69, 614–624. [Google Scholar] [CrossRef]
- Karki, R.; Ho, O.M.; Kim, D.W. Magnolol attenuates neointima formation by inducing cell cycle arrest via inhibition of ERK1/2 and NF-kappaB activation in vascular smooth muscle cells. Biochim. Biophys. Acta 2013, 1830, 2619–2628. [Google Scholar] [CrossRef]
- Liu, H.; Li, Z.; Huo, S.; Wei, Q.; Ge, L. Induction of G0/G1 phase arrest and apoptosis by CRISPR/Cas9-mediated knockout of CDK2 in A375 melanocytes. Mol. Clin. Oncol. 2020, 12, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Liang, C.J.; Leu, Y.L.; Chen, Y.L.; Wang, S.H. Viscolin Inhibits In Vitro Smooth Muscle Cell Proliferation and Migration and Neointimal Hyperplasia In Vivo. PLoS ONE 2016, 11, e0168092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Q.; Li, Y.L.; Li, X.T.; Lv, J.Y.; Gao, Y.; Li, W.N.; Gong, Q.H.; Yang, D.L. Osthole Alleviates Neointimal Hyperplasia in Balloon-Induced Arterial Wall Injury by Suppressing Vascular Smooth Muscle Cell Proliferation and Downregulating Cyclin D1/CDK4 and Cyclin E1/CDK2 Expression. Front. Physiol. 2020, 11, 514494. [Google Scholar] [CrossRef] [PubMed]
- Tanner, F.C.; Boehm, M.; Akyürek, L.M.; San, H.; Yang, Z.Y.; Tashiro, J.; Nabel, G.J.; Nabel, E.G. Differential effects of the cyclin-dependent kinase inhibitors p27(Kip1), p21(Cip1), and p16(Ink4) on vascular smooth muscle cell proliferation. Circulation 2000, 101, 2022–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polyak, K.; Kato, J.Y.; Solomon, M.J.; Sherr, C.J.; Massague, J.; Roberts, J.M.; Koff, A. p27Kip1, a cyclin-Cdk inhibitor, links transforming growth factor-beta and contact inhibition to cell cycle arrest. Genes Dev. 1994, 8, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, R.; Mei, X.; Wang, Y.C.; Cui, X.B.; Zhang, G.; Li, W.; Chen, S.Y. LncRNA GAS5 regulates vascular smooth muscle cell cycle arrest and apoptosis via p53 pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2516–2525. [Google Scholar] [CrossRef] [PubMed]
- Engeland, K. Cell cycle arrest through indirect transcriptional repression by p53: I have a DREAM. Cell Death Differ. 2018, 25, 114–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.K.; Kim, J.W.; Zilberstein, A.; Margolis, B.; Kim, J.G.; Schlessinger, J.; Rhee, S.G. PDGF stimulation of inositol phospholipid hydrolysis requires PLC-gamma 1 phosphorylation on tyrosine residues 783 and 1254. Cell 1991, 65, 435–441. [Google Scholar] [CrossRef]
- Heldin, C.H.; Ostman, A.; Rönnstrand, L. Signal transduction via platelet-derived growth factor receptors. Biochim. Biophys. Acta 1998, 1378, F79–F113. [Google Scholar] [CrossRef]
- Talaia, C.; Morato, M.; Quintas, C.; Gonçalves, J.; Queiroz, G. Functional crosstalk of prejunctional receptors on the modulation of noradrenaline release in mesenteric vessels: A differential study of artery and vein. Eur. J. Pharmacol. 2011, 652, 33–39. [Google Scholar] [CrossRef]
- Fortuño, A.; Rodríguez, A.; Gómez-Ambrosi, J.; Muñiz, P.; Salvador, J.; Díez, J.; Frühbeck, G. Leptin inhibits angiotensin II-induced intracellular calcium increase and vasoconstriction in the rat aorta. Endocrinology 2002, 143, 3555–3560. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-W.; Hsieh, C.-Y.; Yang, C.-H.; Tsai, J.-H.; Huang, S.-Y.; Sheu, J.-R. Yohimbine, an α2-Adrenoceptor Antagonist, Suppresses PDGF-BB-Stimulated Vascular Smooth Muscle Cell Proliferation by Downregulating the PLCγ1 Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 8049. https://doi.org/10.3390/ijms23148049
Chiu C-W, Hsieh C-Y, Yang C-H, Tsai J-H, Huang S-Y, Sheu J-R. Yohimbine, an α2-Adrenoceptor Antagonist, Suppresses PDGF-BB-Stimulated Vascular Smooth Muscle Cell Proliferation by Downregulating the PLCγ1 Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(14):8049. https://doi.org/10.3390/ijms23148049
Chicago/Turabian StyleChiu, Chih-Wei, Cheng-Ying Hsieh, Chih-Hao Yang, Jie-Heng Tsai, Shih-Yi Huang, and Joen-Rong Sheu. 2022. "Yohimbine, an α2-Adrenoceptor Antagonist, Suppresses PDGF-BB-Stimulated Vascular Smooth Muscle Cell Proliferation by Downregulating the PLCγ1 Signaling Pathway" International Journal of Molecular Sciences 23, no. 14: 8049. https://doi.org/10.3390/ijms23148049
APA StyleChiu, C. -W., Hsieh, C. -Y., Yang, C. -H., Tsai, J. -H., Huang, S. -Y., & Sheu, J. -R. (2022). Yohimbine, an α2-Adrenoceptor Antagonist, Suppresses PDGF-BB-Stimulated Vascular Smooth Muscle Cell Proliferation by Downregulating the PLCγ1 Signaling Pathway. International Journal of Molecular Sciences, 23(14), 8049. https://doi.org/10.3390/ijms23148049