
Toxicity of Bacillus thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides besseyi


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
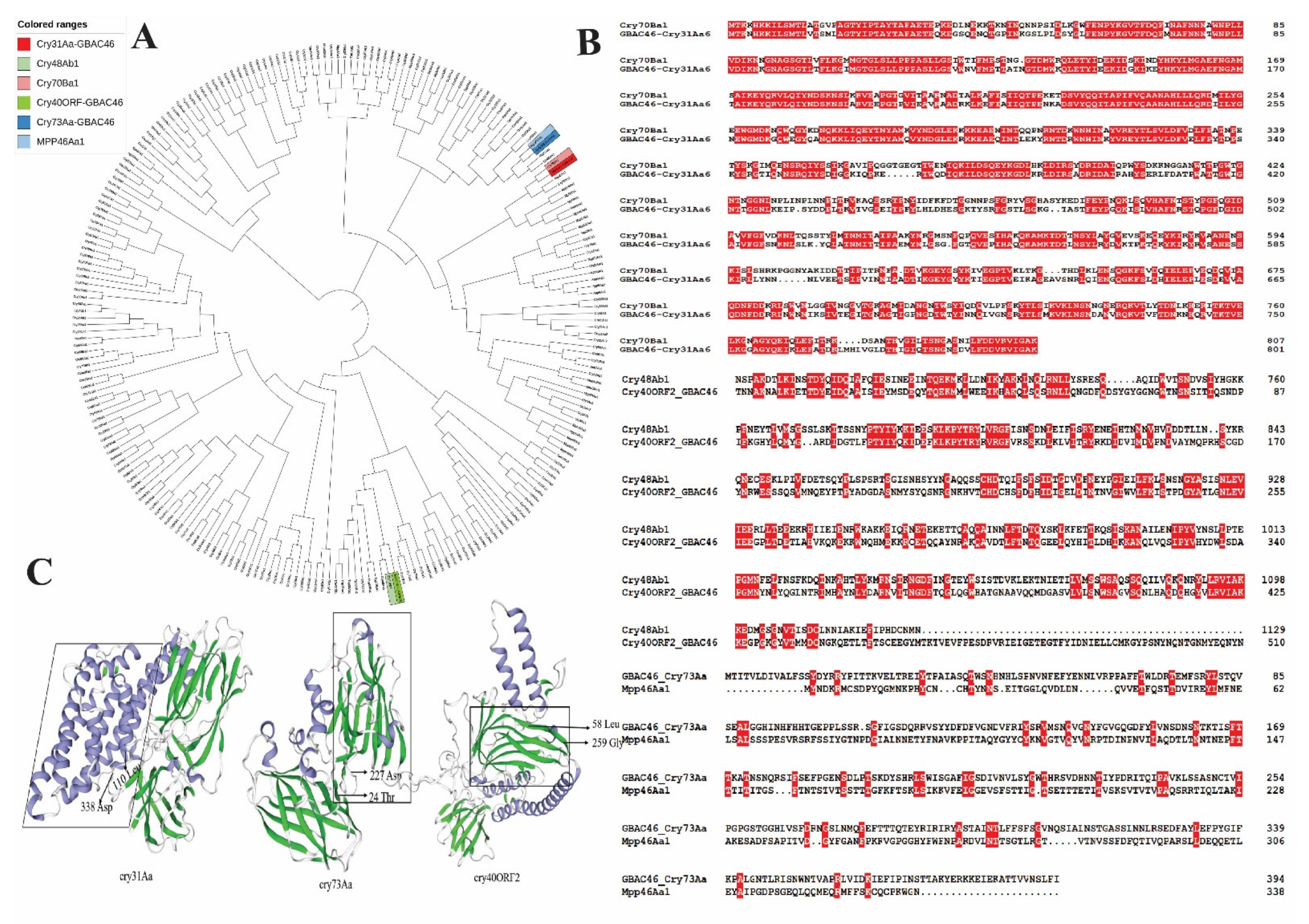
2.1. Sequence Analysis of Cry31Aa and Familiar Cry Protein
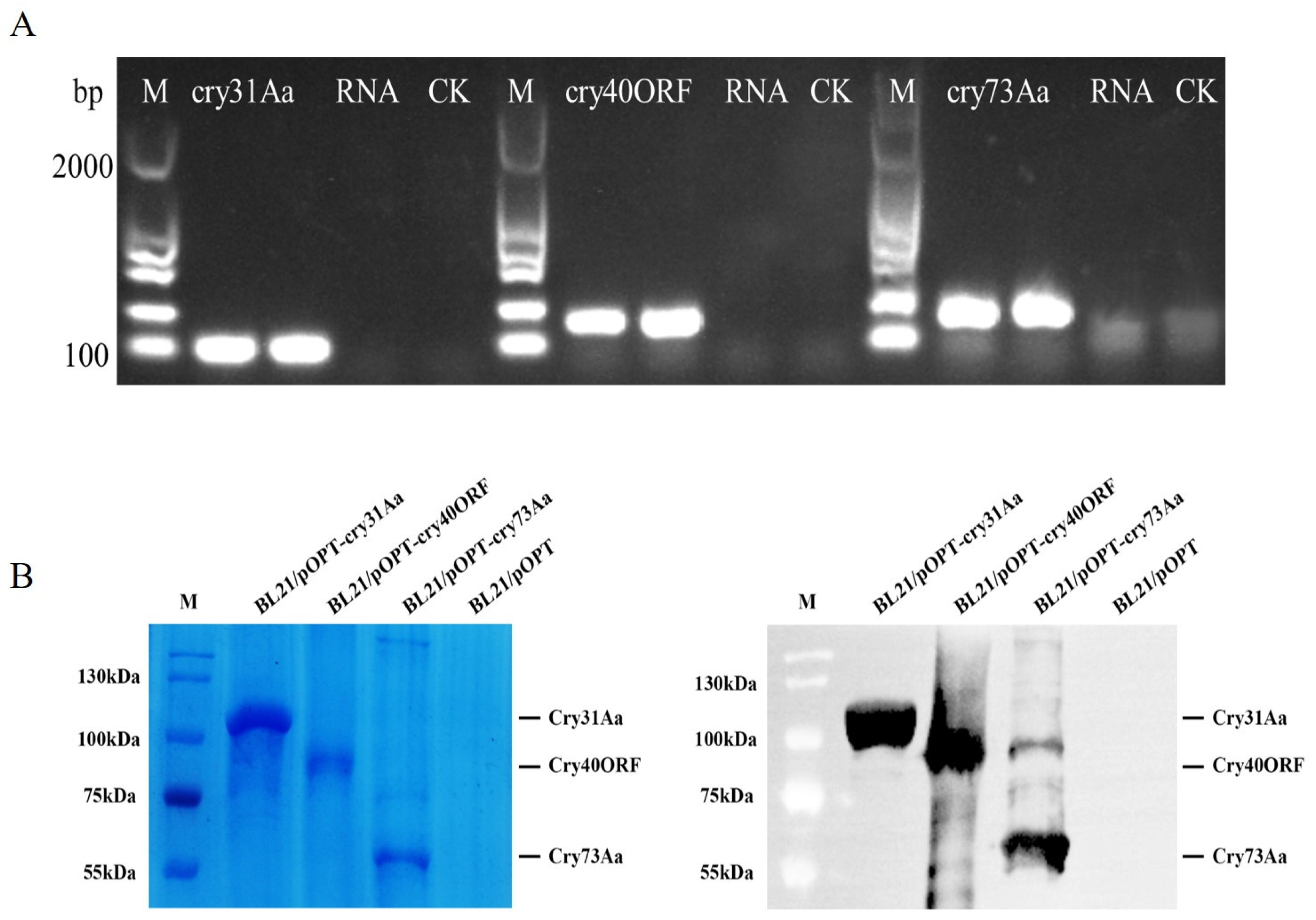
2.2. Expression, Immunoblot, and Purification of the Three Cry Proteins of GBAC46
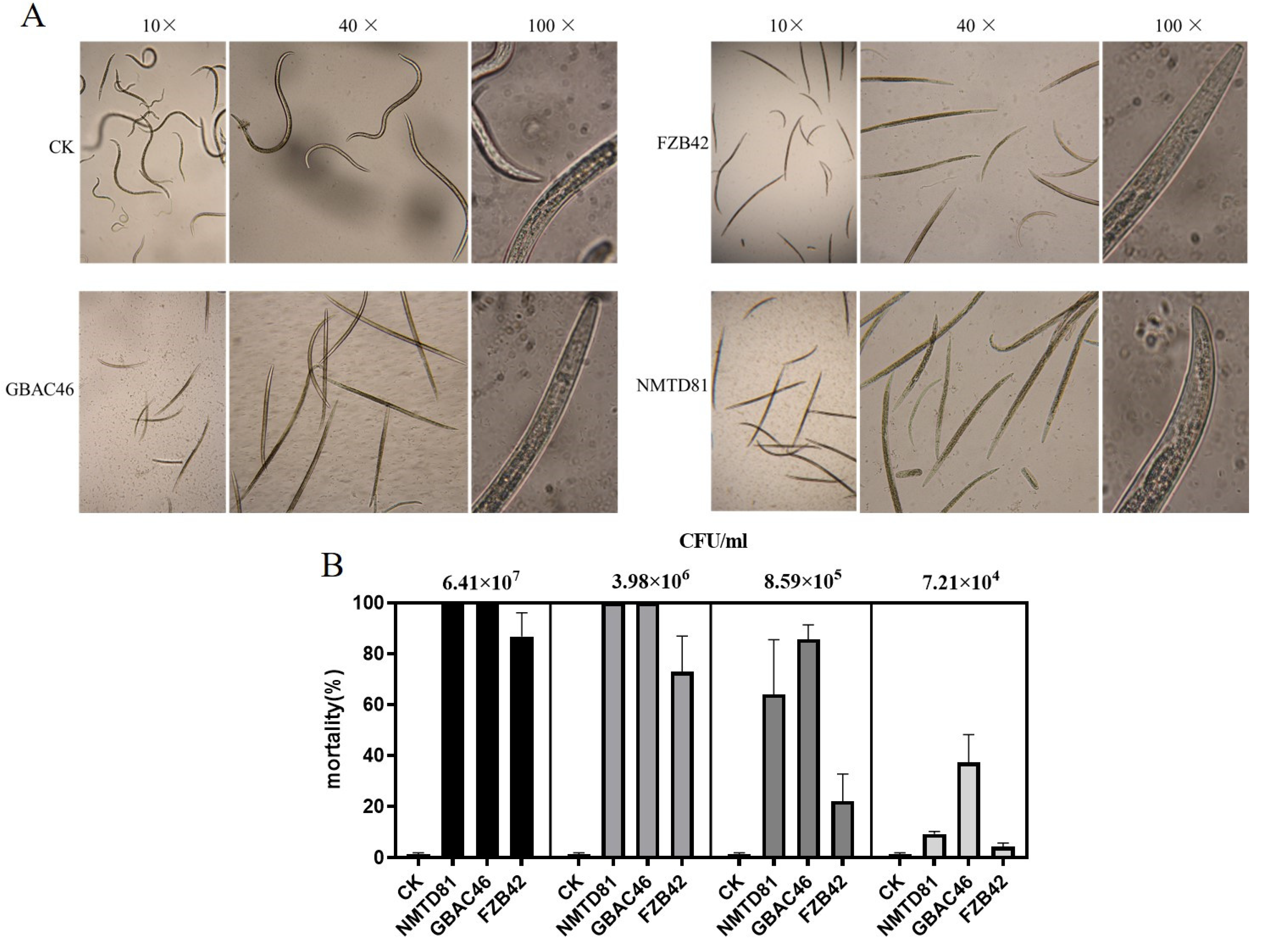
2.3. Virulence of Bacillus Thuringiensis Strains to A. besseyi
2.4. Morphological Observation and Pore-Formation of Bt Strains Via Staining
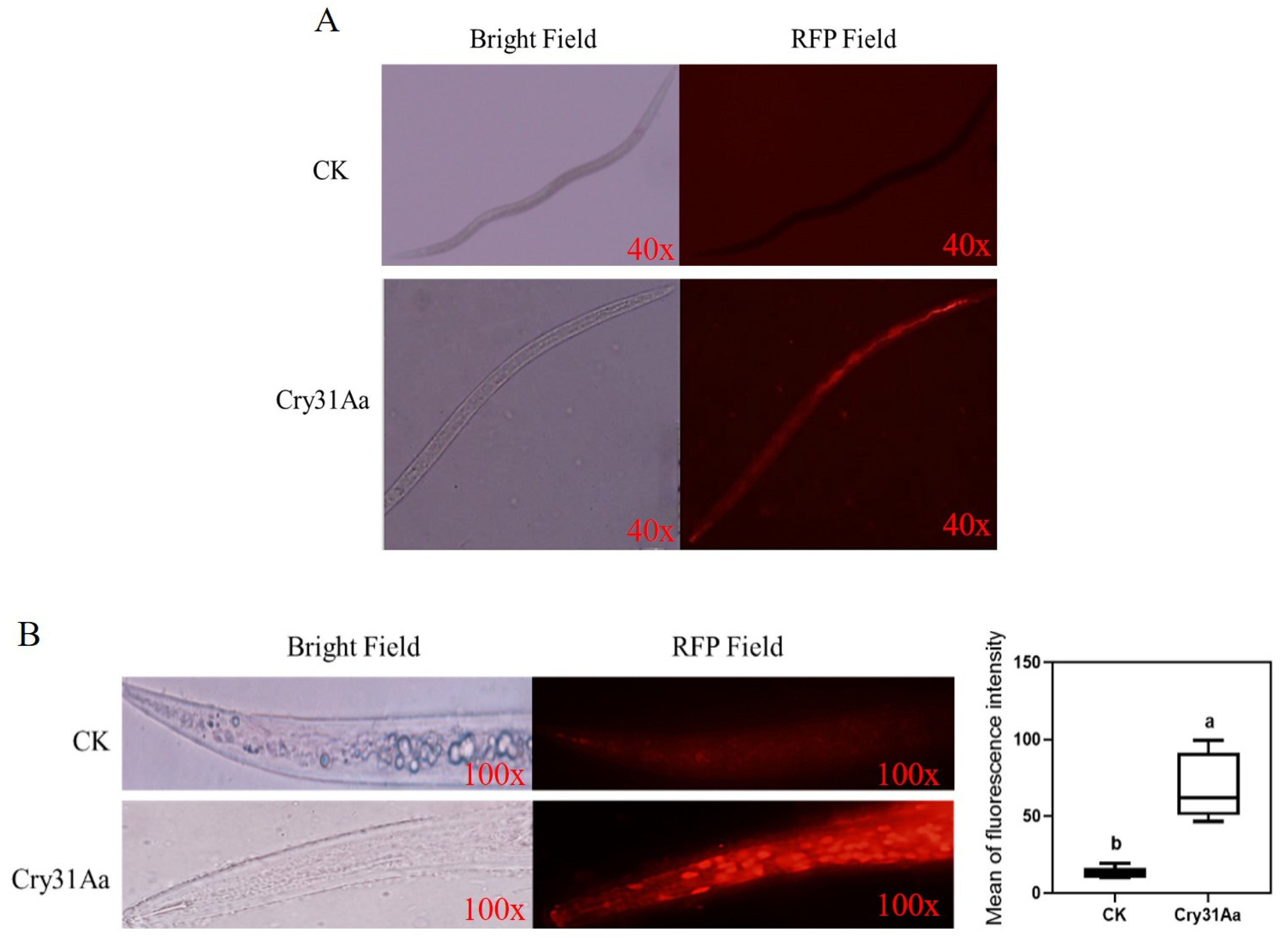
2.5. Toxicity of Cry Protein to A. besseyi
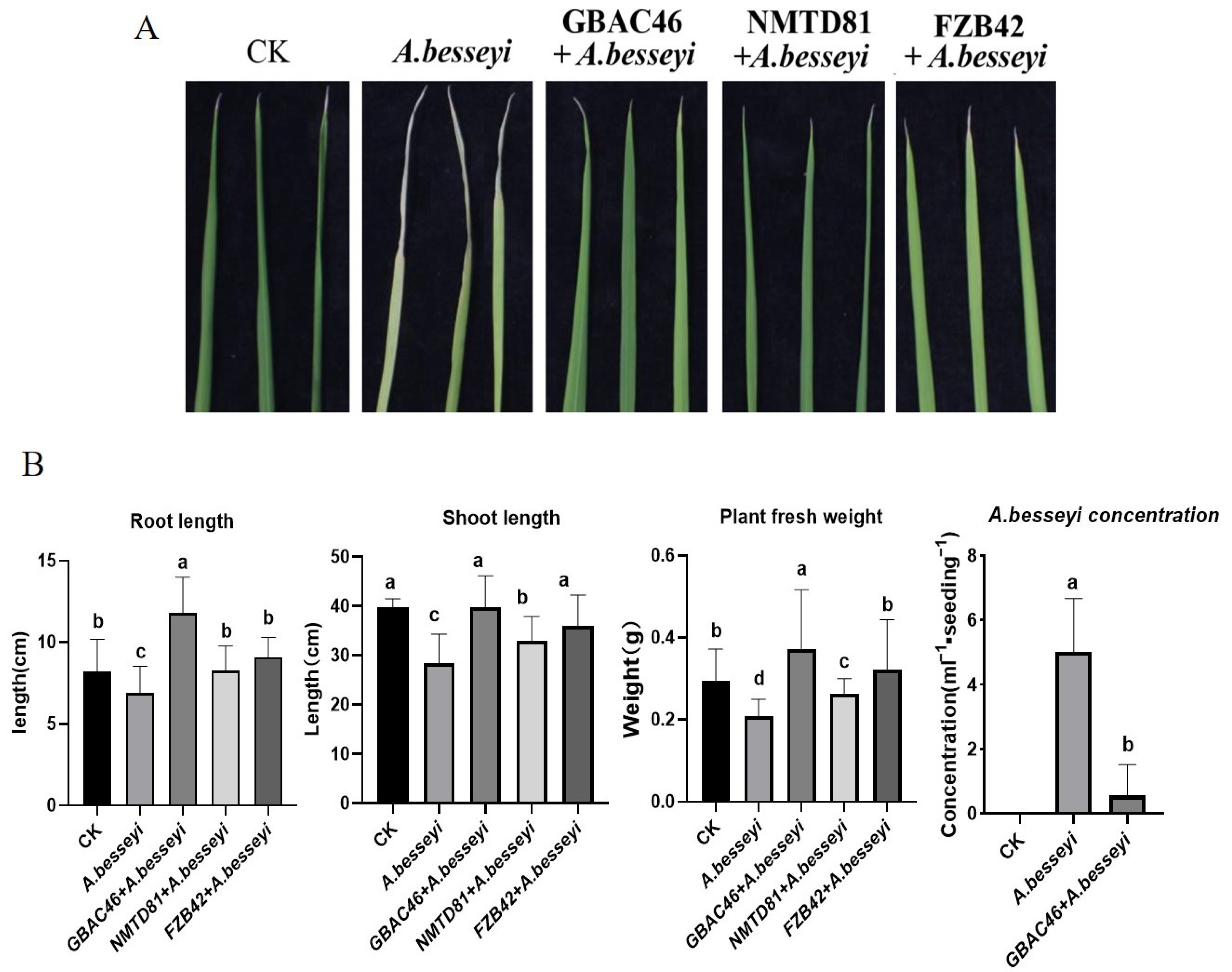
2.6. Plant Growth Promotion Traits of Bt Strains in a Pot Experiment
2.7. Hydrogen Peroxide (H2O2) and Malondialdehyde (MDA) Analysis in Rice
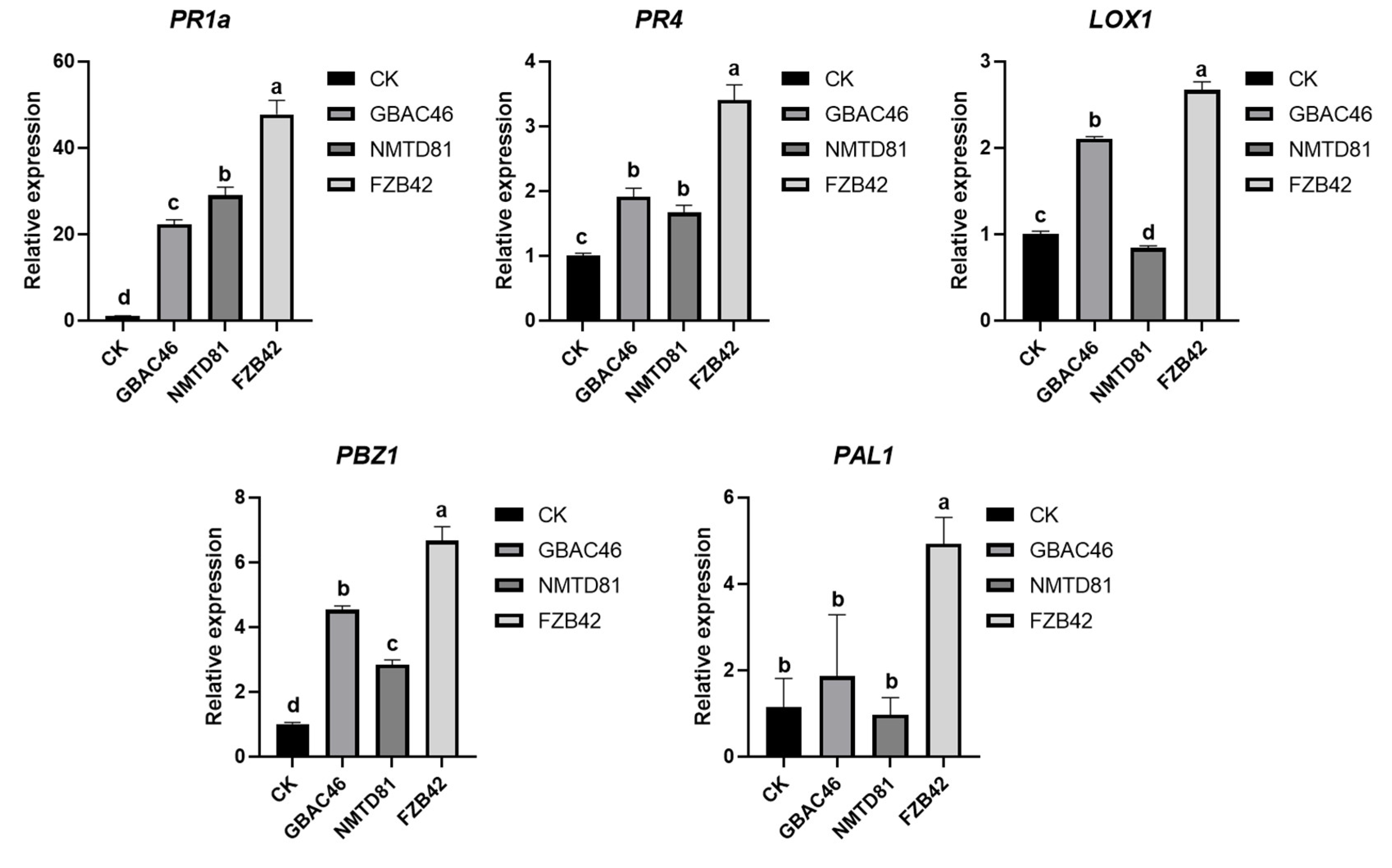
2.8. Relative Expression of Defense-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plasmids, Bacterial Strains, and Cultural Conditions
4.2. Nematode Inoculum
4.3. Bioinformatics Analysis
4.4. Expression of Cry31Aa Protein and Purification
4.5. Total RNA Extraction and cDNA Synthesis for RT-PCR
4.6. Nematode Bioassays
4.7. Determination of Sporulation Formation
4.8. Phenotypic Observation by Scanning Electron Microscopy
4.9. Endocytosis Assays with Rhodamine-Labeled Cry31Aa
4.10. Propidium Iodide Uptake Assays
4.11. In Vivo Experiments with Rice Plants
4.12. Estimation of MDA Level (Lipid Peroxidation) in Rice Plants
4.13. Determination of Hydrogen Peroxide
4.14. Defense-Related Genes’ Expression in Rice Plants
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, Q.; Zheng, H.; Rao, M.J.; Ali, M.; Hussain, A.; Saleem, M.H.; Nehela, Y.; Sohail, M.A.; Ahmed, A.M.; Kubar, K.A.; et al. Advances, limitations, and prospects of biosensing technology for detecting phytopathogenic bacteria. Chemosphere 2022, 296, 133773. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Cui, H.; Yang, X.; Xu, L.; Li, X.; Liu, R.; Yan, Z.; Li, X.; Zheng, W.; Zhao, Y.; et al. Nematicidal activity against Aphelenchoides besseyi and Ditylenchus destructor of three biflavonoids, isolated from roots of Stellera chamaejasme. J. Asia Pac. Entomol. 2018, 21, 1473–1478. [Google Scholar] [CrossRef]
- Cheng, C.; Gao, X.; Feng, B.; Sheen, J.; Ssheen, L.; He, P. Differential temperature operation of plant immune responses. Nat. Commun. 2013, 4, 2530. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Lin, X.; Poland, J.; Trick, H.; Leach, J.; Hulbert, S. A maize resistance gene functions against bacterial streak disease in rice. Proc. Natl. Acad. Sci. USA 2005, 102, 15383–15388. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Li, D.; Wang, Z.; Dong, A.; Liu, L.; Wang, B.; Chen, Q.; Liu, X. Transcriptomic analysis of the rice white tip nematode, Aphelenchoides besseyi (Nematoda: Aphelenchoididae). PLoS ONE 2014, 9, e91591. [Google Scholar] [CrossRef] [Green Version]
- Gu, Q.; Qiao, J.; Wang, R.; Lu, J.; Wang, Z.; Li, P.; Zhang, L.; Ali, Q.; Khan, A.R.; Gao, X.; et al. The Role of Pyoluteorin from Pseudomonas protegens Pf-5 in Suppressing the Growth and Pathogenicity of Pantoea ananatis on Maize. Int. J. Mol. Sci. 2022, 23, 6431. [Google Scholar] [CrossRef]
- Yu, C.; Liu, X.; Zhang, X.; Zhang, M.; Gu, Y.; Ali, Q.; Mohamed, M.; Xu, J.; Shi, J.; Gao, X. Mycosubtilin Produced by Bacillus subtilis ATCC6633 Inhibits Growth and Mycotoxin Biosynthesis of Fusarium graminearum and Fusarium verticillioides. Toxins 2021, 13, 791. [Google Scholar] [CrossRef]
- Jasmer, D.P.; Rosa, B.A.; Tyagi, R.; Mitreva, M. Rapid determination of nematode cell and organ susceptibility to toxic treatments. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 167–182. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Farzand, A.; Khan, A.R.; Ling, H.; Gao, X. Nematicidal volatiles from bacillus atrophaeus gbsc56 promote growth and stimulate induced systemic resistance in tomato against meloidogyne incognita. Int. J. Mol. Sci. 2021, 22, 5049. [Google Scholar] [CrossRef]
- Yu, Z.; Xiong, J.; Zhou, Q.; Luo, H.; Hu, S.; Xia, L.; Sun, M.; Li, L.; Yu, Z. The diverse nematicidal properties and biocontrol efficacy of Bacillus thuringiensis Cry6A against the root-knot nematode Meloidogyne hapla. J. Invertebr. Pathol. 2015, 125, 73–80. [Google Scholar] [CrossRef]
- Peng, D.; Wan, D.; Cheng, C.; Ye, X.; Sun, M. Nematode-specific cadherin CDH-8 acts as a receptor for Cry5B toxin in Caenorhabditis elegans. Appl. Microbiol. Biotechnol. 2018, 102, 3663–3673. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R. V Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [Green Version]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Lin, J.; Du, H.; Zhang, Y.; Bravo, A.; Soberón, M.; Sun, M.; Peng, D. Bacillus thuringiensis targets the host intestinal epithelial junctions for successful infection of Caenorhabditis elegans. Environ. Microbiol. 2019, 21, 1086–1098. [Google Scholar] [CrossRef]
- Rosas-García, N.M. Biopesticide production from Bacillus thuringiensis: An environmentally friendly alternative. Recent Pat. Biotechnol. 2009, 3, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Chandra, A.; Pandey, K.C. Bacillus thuringiensis (Bt) transgenic crop: An environment friendly insect-pest management strategy. J. Environ. Biol. 2008, 29, 641–653. [Google Scholar]
- Stewart, C.N., Jr.; Adang, M.J.; All, J.N.; Raymer, P.L.; Ramachandran, S.; Parrott, W.A. Insect control and dosage effects in transgenic canola containing a synthetic Bacillus thuringiensis cryIAc gene. Plant Physiol. 1996, 112, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Kumar, N.R.; Maniraj, R.; Lakshmikanth, R.; Rao, K.Y.S.; Muralimohan, N.; Arulprakash, T.; Karthik, K.; Shashibhushan, N.B.; Vinutha, T. Expression of Cry2Aa, a Bacillus thuringiensis insecticidal protein in transgenic pigeon pea confers resistance to gram pod borer, Helicoverpa armigera. Sci. Rep. 2018, 8, 8820. [Google Scholar] [CrossRef]
- Bříza, J.; Pavingerová, D.; Vlasák, J.; Niedermeierová, H. Norway spruce (Picea abies) genetic transformation with modified Cry3A gene of Bacillus thuringiensis. Acta Biochim. Pol. 2013, 60, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.; Haegeman, A.; Danchin, E.; Gaur, H.; Helder, J.; Jones, M.; Kikuchi, T.; Manzanilla-Lopez, R.; Palomares-Rius, J.; Wesemael, W. Top 10 plant-parasitic nematodes in molecular plant pathology. In Proceedings of the 6th International Congress of Nematology, Cape Town, South Africa, 4–9 May 2014. [Google Scholar]
- Ali, Q.; Ahmar, S.; Sohail, M.A.; Kamran, M.; Ali, M.; Saleem, M.H.; Rizwan, M.; Ahmed, A.M.; Mora-Poblete, F.; do Amaral Júnior, A.T.; et al. Research advances and applications of biosensing technology for the diagnosis of pathogens in sustainable agriculture. Environ. Sci. Pollut. Res. 2021, 28, 9002–9019. [Google Scholar] [CrossRef]
- Ali, M.; Ali, Q.; Sohail, M.A.; Ashraf, M.F.; Saleem, M.H.; Hussain, S.; Zhou, L. Diversity and Taxonomic Distribution of Endophytic Bacterial Community in the Rice Plant and Its Prospective. Int. J. Mol. Sci. 2021, 22, 10165. [Google Scholar] [CrossRef]
- Hu, Y.; Georghiou, S.B.; Kelleher, A.J.; Aroian, R.V. Bacillus thuringiensis Cry5B protein is highly efficacious as a single-dose therapy against an intestinal roundworm infection in mice. PLoS Negl. Trop. Dis. 2010, 4, e614. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Zhou, D.; Daly, P.; Wang, X.; Wei, L. Characterization and functional importance of two glycoside hydrolase family 16 genes from the rice white tip nematode Aphelenchoides besseyi. Animals 2021, 11, 374. [Google Scholar] [CrossRef]
- Shi, J.; Peng, D.; Zhang, F.; Ruan, L.; Sun, M. The Caenorhabditis elegans CUB-like-domain containing protein RBT-1 functions as a receptor for Bacillus thuringiensis Cry6Aa toxin. PLoS Pathog. 2020, 16, e1008501. [Google Scholar] [CrossRef]
- Armada, E.; Probanza, A.; Roldán, A.; Azcón, R. Native plant growth promoting bacteria Bacillus thuringiensis and mixed or individual mycorrhizal species improved drought tolerance and oxidative metabolism in Lavandula dentata plants. J. Plant Physiol. 2016, 192, 1–12. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Yu, C.; Wang, Y.; Gu, Q.; Wu, H.; Gao, X. Cadmium tolerant microbial strains possess different mechanisms for cadmium biosorption and immobilization in rice seedlings. Chemosphere 2022, 303, 135206. [Google Scholar] [CrossRef]
- Ben Abdallah, D.; Frikha-Gargouri, O.; Tounsi, S. Rizhospheric competence, plant growth promotion and biocontrol efficacy of Bacillus amyloliquefaciens subsp. plantarum strain 32a. Biol. Control 2018, 124, 61–67. [Google Scholar] [CrossRef]
- Cherif-Silini, H.; Silini, A.; Yahiaoui, B.; Ouzari, I.; Boudabous, A. Phylogenetic and plant-growth-promoting characteristics of Bacillus isolated from the wheat rhizosphere. Ann. Microbiol. 2016, 66, 1087–1097. [Google Scholar] [CrossRef]
- Raddadi, N.; Cherif, A.; Ouzari, H.; Marzorati, M.; Brusetti, L.; Boudabous, A.; Daffonchio, D. Bacillus thuringiensis beyond insect biocontrol: Plant growth promotion and biosafety of polyvalent strains. Ann. Microbiol. 2007, 57, 481–494. [Google Scholar] [CrossRef]
- Liaquat, F.; Munis, M.F.H.; Arif, S.; Haroon, U.; Shi, J.; Saqib, S.; Zaman, W.; Che, S.; Liu, Q. PacBio Single-Molecule Long-Read Sequencing Reveals Genes Tolerating Manganese Stress in Schima superba Saplings. Front. Genet. 2021, 12, 635043. [Google Scholar] [CrossRef] [PubMed]
- Azizoglu, U. Bacillus thuringiensis as a biofertilizer and biostimulator: A mini-review of the little-known plant growth-promoting properties of Bt. Curr. Microbiol. 2019, 76, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Iatsenko, I.; Boichenko, I.; Sommer, R.J. Bacillus thuringiensis DB27 produces two novel protoxins, Cry21Fa1 and Cry21Ha1, which act synergistically against nematodes. Appl. Environ. Microbiol. 2014, 80, 3266–3275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, W.; Zhu, L.; Liu, Y.; Crickmore, N.; Peng, D.; Ruan, L.; Sun, M. Mining new crystal protein genes from Bacillus thuringiensis on the basis of mixed plasmid-enriched genome sequencing and a computational pipeline. Appl. Environ. Microbiol. 2012, 78, 4795–4801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crickmore, N.; Berry, C.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2021, 186, 107438. [Google Scholar] [CrossRef]
- Peng, Q.; Yu, Q.; Song, F. Expression of cry genes in Bacillus thuringiensis biotechnology. Appl. Microbiol. Biotechnol. 2019, 103, 1617–1626. [Google Scholar] [CrossRef]
- Vilas-Bôas, G.T.; Peruca, A.P.S.; Arantes, O.M.N. Biology and taxonomy of Bacillus cereus, Bacillus anthracis, and Bacillus thuringiensis. Can. J. Microbiol. 2007, 53, 673–687. [Google Scholar] [CrossRef]
- Vílchez, S. Making 3D-cry toxin mutants: Much more than a tool of understanding toxins mechanism of action. Toxins 2020, 12, 600. [Google Scholar] [CrossRef]
- Xu, L.-N.; Wang, Y.-Q.; Wang, Z.-Y.; Hu, B.-J.; Ling, Y.-H.; He, K.-L. Transcriptome differences between Cry1Ab resistant and susceptible strains of Asian corn borer. BMC Genom. 2015, 16, 173. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Lin, Q.; Qian, X.; Zheng, Y.; Yao, J.; Wu, H.; Li, M.; Jin, X.; Pan, X.; Zhang, L. Nematicidal activity of Cry1Ea11 from Bacillus thuringiensis BRC-XQ12 against the pine wood nematode (Bursaphelenchus xylophilus). Phytopathology 2018, 108, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Rai, V.; Acharya, S.; Dey, N. Implications of Nanobiosensors in Agriculture. J. Biomater. Nanobiotechnol. 2012, 3, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.; Rae, R.; Iatsenko, I.; Sommer, R.J. System wide analysis of the evolution of innate immunity in the nematode model species Caenorhabditis elegans and Pristionchus pacificus. PLoS ONE 2012, 7, e44255. [Google Scholar] [CrossRef]
- Liu, Z.; Budiharjo, A.; Wang, P.; Shi, H.; Fang, J.; Borriss, R.; Zhang, K.; Huang, X. The highly modified microcin peptide plantazolicin is associated with nematicidal activity of Bacillus amyloliquefaciens FZB42. Appl. Microbiol. Biotechnol. 2013, 97, 10081–10090. [Google Scholar] [CrossRef]
- Guo, S.; Liu, M.; Peng, D.; Ji, S.; Wang, P.; Yu, Z.; Sun, M. New strategy for isolating novel nematicidal crystal protein genes from Bacillus thuringiensis strain YBT-1518. Appl. Environ. Microbiol. 2008, 74, 6997–7001. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Luo, H.; Xiong, J.; Zhou, Q.; Xia, L.; Sun, M.; Li, L.; Yu, Z. B acillus thuringiensis C ry6 A exhibits nematicidal activity to C aenorhabditis elegans bre mutants and synergistic activity with C ry5 B to C. elegans. Lett. Appl. Microbiol. 2014, 58, 511–519. [Google Scholar] [CrossRef]
- Ji, F.; Zhu, Y.; Ju, S.; Zhang, R.; Yu, Z.; Sun, M. Promoters of crystal protein genes do not control crystal formation inside exosporium of Bacillus thuringiensis ssp. finitimus strain YBT-020. FEMS Microbiol. Lett. 2009, 300, 11–17. [Google Scholar] [CrossRef]
- Ammons, D.R.; Reyna, A.; Granados, J.C.; Ventura-Suárez, A.; Rojas-Avelizapa, L.I.; Short, J.D.; Rampersad, J.N. A novel Bacillus thuringiensis Cry-like protein from a rare filamentous strain promotes crystal localization within the exosporium. Appl. Environ. Microbiol. 2013, 79, 5774–5776. [Google Scholar] [CrossRef] [Green Version]
- Belair, G. Nematodes, these roundworms that harm plants…by their roots. Phytoprotection 2005, 86, 65–69. [Google Scholar]
- Eves-van den Akker, S.; Lilley, C.J.; Ault, J.R.; Ashcroft, A.E.; Jones, J.T.; Urwin, P.E. The feeding tube of cyst nematodes: Characterisation of protein exclusion. PLoS ONE 2014, 9, e87289. [Google Scholar]
- Kahn, T.W.; Duck, N.B.; McCarville, M.T.; Schouten, L.C.; Schweri, K.; Zaitseva, J.; Daum, J. A Bacillus thuringiensis Cry protein controls soybean cyst nematode in transgenic soybean plants. Nat. Commun. 2021, 12, 3380. [Google Scholar] [CrossRef]
- Ramalakshmi, A.; Sharmila, R.; Iniyakumar, M.; Gomathi, V. Nematicidal activity of native Bacillus thuringiensis against the root knot nematode, Meloidogyne incognita (Kofoid and White). Egypt. J. Biol. Pest Control 2020, 30, 90. [Google Scholar] [CrossRef]
- Chandler, S.; Van Hese, N.; Coutte, F.; Jacques, P.; Höfte, M.; De Vleesschauwer, D. Role of cyclic lipopeptides produced by Bacillus subtilis in mounting induced immunity in rice (Oryza sativa L.). Physiol. Mol. Plant Pathol. 2015, 91, 20–30. [Google Scholar] [CrossRef]
- Huang, H.; Ayaz, A.; Zheng, M.; Yang, X.; Zaman, W.; Zhao, H.; Lü, S. Arabidopsis KCS5 and KCS6 Play Redundant Roles in Wax Synthesis. Int. J. Mol. Sci. 2022, 23, 4450. [Google Scholar] [CrossRef]
- Mengiste, T. Plant immunity to necrotrophs. Annu. Rev. Phytopathol. 2012, 50, 267–294. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A. Mechanism of tobacco osmotin gene in plant responses to biotic and abiotic stress tolerance: A brief history. Biocell 2022, 46, 623–632. [Google Scholar] [CrossRef]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular cloning and functional analysis of GmLACS2-3 reveals its involvement in cutin and suberin biosynthesis along with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Weber, H.; Chételat, A.; Reymond, P.; Farmer, E.E. Selective and powerful stress gene expression in Arabidopsis in response to malondialdehyde. Plant J. 2004, 37, 877–888. [Google Scholar] [CrossRef]
- Salehi Jouzani, G.; Seifinejad, A.; Saeedizadeh, A.; Nazarian, A.; Yousefloo, M.; Soheilivand, S.; Mousivand, M.; Jahangiri, R.; Yazdani, M.; Amiri, R.M.; et al. Molecular detection of nematicidal crystalliferous Bacillus thuringiensis strains of Iran and evaluation of their toxicity on free-living and plant-parasitic nematodes. Can. J. Microbiol. 2008, 54, 812–822. [Google Scholar] [CrossRef]
- Wilson, M.K.; Abergel, R.J.; Raymond, K.N.; Arceneaux, J.E.L.; Byers, B.R. Siderophores of Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis. Biochem. Biophys. Res. Commun. 2006, 348, 320–325. [Google Scholar] [CrossRef]
- Manghwar, H.; Hussain, A.; Ali, Q.; Saleem, M.H.; Abualreesh, M.H.; Alatawi, A.; Ali, S.; Munis, M.F.H. Disease Severity, Resistance Analysis, and Expression Profiling of Pathogenesis-Related Protein Genes after the Inoculation of Fusarium equiseti in Wheat. Agronomy 2021, 11, 2124. [Google Scholar] [CrossRef]
- Kong, M.; Sheng, T.; Liang, J.; Ali, Q.; Gu, Q.; Wu, H.; Chen, J.; Liu, J.; Gao, X. Melatonin and Its Homologs Induce Immune Responses via Receptors trP47363-trP13076 in Nicotiana benthamiana. Front. Plant Sci. 2021, 12, 691835. [Google Scholar] [CrossRef] [PubMed]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Safdar, A.; Huang, Z.; Rajer, F.U.; Gao, X. Effect of volatile compounds produced by Ralstonia solanacearum on plant growth promoting and systemic resistance inducing potential of Bacillus volatiles. BMC Plant Biol. 2017, 17, 133. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Gu, Q.; Xie, Y.; Lou, Z.; Xue, P.; Fang, L.; Yu, C.; Jia, D.; Huang, G.; Zhu, B.; et al. Cold-adapted Bacilli isolated from the Qinghai—Tibetan Plateau are able to promote plant growth in extreme environments. Environ. Microbiol. 2019, 21, 3505–3526. [Google Scholar] [CrossRef]
- Fernandez-Luna, M.T.; Kumar, P.; Hall, D.G.; Mitchell, A.D.; Blackburn, M.B.; Bonning, B.C. Toxicity of Bacillus thuringiensis-derived pesticidal proteins cry1Ab and cry1Ba against asian citrus psyllid, Diaphorina citri (Hemiptera). Toxins 2019, 11, 173. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Shao, Y.; Wei, L.H.; Gao, C.Y.; Zhou, Y.J. The white-tip nematode, Aphelenchoides besseyi, exhibits an auxin-orientated behaviour affecting its migration and propagation. Nematology 2014, 16, 837–845. [Google Scholar] [CrossRef]
- Viglierchio, D.R.; Schmitt, R. V On the methodology of nematode extraction from field samples: Baermann funnel modifications. J. Nematol. 1983, 15, 438–444. [Google Scholar]
- Liang, Z.; Qiao, J.Q.; Li, P.P.; Zhang, L.L.; Qiao, Z.X.; Lin, L.; Yu, C.J.; Yang, Y.; Zubair, M.; Gu, Q.; et al. A novel Rap-Phr system in Bacillus velezensis NAU-B3 regulates surfactin production and sporulation via interaction with ComA. Appl. Microbiol. Biotechnol. 2020, 104, 10059–10074. [Google Scholar] [CrossRef]
- Wu, H.; Yang, Y.; Wang, S.; Qiao, J.; Xia, Y.; Wang, Y.; Wang, W.; Gao, S.; Liu, J.; Xue, P. Cloning, expression and characterization of a new aspartate aminotransferase from Bacillus subtilis B3. FEBS J. 2011, 278, 1345–1357. [Google Scholar] [CrossRef]
- Kourtis, N.; Nikoletopoulou, V.; Tavernarakis, N. Small heat-shock proteins protect from heat-stroke-associated neurodegeneration. Nature 2012, 490, 213–218. [Google Scholar] [CrossRef]
- Chen, H.; Gong, Y.; Han, R. Cadmium telluride quantum dots (CdTe-QDs) and enhanced ultraviolet-B (UV-B) radiation trigger antioxidant enzyme metabolism and programmed cell death in wheat seedlings. PLoS ONE 2014, 9, e110400. [Google Scholar] [CrossRef]
- Kong, M.; Liang, J.; Ali, Q.; Wen, W.; Wu, H.; Gao, X.; Gu, Q. 5-Methoxyindole, a Chemical Homolog of Melatonin, Adversely Affects the Phytopathogenic Fungus Fusarium graminearum. Int. J. Mol. Sci. 2021, 22, 10991. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Z.; Ali, Q.; Wang, Y.; Mu, G.; Kan, X.; Ren, Y.; Manghwar, H.; Gu, Q.; Wu, H.; Gao, X. Toxicity of Bacillus thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides besseyi. Int. J. Mol. Sci. 2022, 23, 8189. https://doi.org/10.3390/ijms23158189
Liang Z, Ali Q, Wang Y, Mu G, Kan X, Ren Y, Manghwar H, Gu Q, Wu H, Gao X. Toxicity of Bacillus thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides besseyi. International Journal of Molecular Sciences. 2022; 23(15):8189. https://doi.org/10.3390/ijms23158189
Chicago/Turabian StyleLiang, Zhao, Qurban Ali, Yujie Wang, Guangyuan Mu, Xuefei Kan, Yajun Ren, Hakim Manghwar, Qin Gu, Huijun Wu, and Xuewen Gao. 2022. "Toxicity of Bacillus thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides besseyi" International Journal of Molecular Sciences 23, no. 15: 8189. https://doi.org/10.3390/ijms23158189
APA StyleLiang, Z., Ali, Q., Wang, Y., Mu, G., Kan, X., Ren, Y., Manghwar, H., Gu, Q., Wu, H., & Gao, X. (2022). Toxicity of Bacillus thuringiensis Strains Derived from the Novel Crystal Protein Cry31Aa with High Nematicidal Activity against Rice Parasitic Nematode Aphelenchoides besseyi. International Journal of Molecular Sciences, 23(15), 8189. https://doi.org/10.3390/ijms23158189