
Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Nutrient Management Approaches to Alleviate the Abiotic Stresses
2.1. Primary Nutrients
2.1.1. Nitrogen
2.1.2. Phosphorous
2.1.3. Potassium
2.2. Secondary Nutrients
2.2.1. Calcium
2.2.2. Magnesium
2.2.3. Sulphur
2.3. Micronutrients
2.3.1. Boron
2.3.2. Zinc
2.3.3. Iron
2.3.4. Copper
2.4. Beneficial Nutrients
2.4.1. Cobalt
2.4.2. Selenium
2.4.3. Silicon
3. Combined Effects of Plant Nutrients
4. Conclusions and Future Needs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malhi, Y.; Franklin, J.; Seddon, N.; Solan, M.; Turner, M.G.; Field, C.B.; Knowlton, N. Climate change and ecosystems: Threats, opportunities and solutions. Philos. Trans. R. Soc. B 2020, 375, 20190104. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil. Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Liang, T.B.; Wang, Z.L.; Wang, R.J.; Liu, L.L.; Shi, C.Y. Effects of potassium humate on ginger root growth and its active oxygen metabolism. J. Appl. Ecol. 2007, 18, 813–817. [Google Scholar]
- Seleiman, M.F.; Kheir, A.S. Maize productivity, heavy metals uptake and their availability in contaminated clay and sandy alkaline soils as affected by inorganic and organic amendments. Chemosphere 2018, 204, 514–522. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group, I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- UNFCC. The Glasgow Climate Pact–Key Outcomes from COP26. 2022. Available online: https://unfccc.int/process-and-meetings/the-paris-agreement/the-glasgow-climate-pact-key-outcomes-fromcop26#:~:text=The%20package%20of%20decisions%20consists,the%20necessary%20finance%20for%20both (accessed on 18 February 2022).
- Hajiboland, R. Effect of Micronutrient Deficiencies on Plants Stress Responses. In Abiotic Stress Responses in Plants; Springer: New York, NY, USA, 2012; pp. 283–329. [Google Scholar]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Aslam, M.T. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, W.; Yu, H.; Yang, X. Exogenous abscisic acid alleviates low temperature-induced oxidative damage in seedlings of Cucumis sativus L. Trans. Chin. Soc. Agric. Eng. 2012, 28, 221–228. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y.; Saifullah Ahmad, M. Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agric. Scand. B Soil. Plant Sci. 2011, 61, 291–304. [Google Scholar] [CrossRef]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Reduced reliance on the trace element selenium during evolution of mammals. Genome Biol. 2008, 9, R62. [Google Scholar] [CrossRef] [Green Version]
- Peck, A.W.; McDonald, G.K. Adequate zinc nutrition alleviates the adverse effects of heat stress in bread wheat. Plant Soil. 2010, 337, 355–374. [Google Scholar] [CrossRef]
- Nievola, C.C.; Carvalho, C.P.; Carvalho, V.; Rodrigues, E. Rapid responses of plants to temperature changes. Temperature 2017, 4, 371–405. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Chakraborty, B.; Chakraborty, U. Plant growth promoting rhizobacteria protect wheat plants against temperature stress through antioxidant signalling and reducing chloroplast and membrane injury. J. Plant Growth Regul. 2018, 37, 1396–1412. [Google Scholar] [CrossRef]
- Demirel, U.; Morris, W.L.; Ducreux, L.J.; Yavuz, C.; Asim, A.; Tindas, I.; Hancock, R.D. Physiological, biochemical, and transcriptional responses to single and combined abiotic stress in stress-tolerant and stress-sensitive potato genotypes. Front. Plant Sci. 2020, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Esim, N.; Atici, O. Nitric oxide improves chilling tolerance of maize by affecting apoplastic antioxidative enzymes in leaves. Plant Growth Regul. 2014, 72, 29–38. [Google Scholar] [CrossRef]
- Megha, S.; Basu, U.; Kav, N.N. Regulation of low temperature stress in plants by microRNAs. Plant Cell Environ. 2018, 41, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Khare, T.; Sharma, M.; Wani, S.H. ROS-Induced Signaling and Gene Expression in Crops under Salinity Stress. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 159–184. [Google Scholar]
- Jalil, S.U.; Ansari, M.I. Physiological Role of Gamma-Aminobutyric Acid in Salt Stress Tolerance. In Salt and Drought Stress Tolerance in Plants; Hasanuzzaman, M., Tanveer, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 337–350. [Google Scholar]
- Iqbal, M.S.; Singh, A.K.; Ansari, M.I. Effect of Drought Stress on Crop Production. In New Frontiers in Stress Management for Durable Agriculture; Rakshit, A., Singh, H.B., Singh, A.K., Singh, U.S., Fraceto, L., Eds.; Springer: Singapore, 2020; pp. 35–47. [Google Scholar]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A. Waterlogging stress in plants: A Review. Afr. J. Agric. Res. 2012, 7, 1976–1981. [Google Scholar]
- Noshi, M.; Hatanaka, R.; Tanabe, N.; Terai, Y.; Maruta, T.; Shigeoka, S. Redox regulation of ascorbate and glutathione by a chloroplastic dehydroascorbate reductase is required for high-light stress tolerance in Arabidopsis. Biosci. Biotechnol. 2016, 80, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Zhu, J.K. Plant abiotic stress response and nutrient use efficiency. Sci. China Life 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling, and molecular interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Morton, M.J.L.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrão, S.; Tester, M. Salt stress under the scalpel—Dissecting the genetics of salt tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Tahir, M.A.; Aziz, T.; Rahmatullah. Silicon-induced growth and yield enhancement in two wheat genotypes differing in salinity tolerance. Commun. Soil. Sci. Plant Anal. 2011, 42, 395–407. [Google Scholar] [CrossRef]
- Tran, T.T.; Kano-Nakata, M.; Takeda, M.; Menge, D.; Mitsuya, S.; Inukai, Y.; Yamauchi, A. Nitrogen application enhanced the expression of developmental plasticity of root systems triggered by mild drought stress in rice. Plant Soil. 2014, 378, 139–152. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Cui, Y.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T. Nitrogen Nutrition Improves the Potential of Wheat (Triticum aestivum L.) to Alleviate the Effects of Drought Stress during Vegetative Growth Periods. Front. Plant Sci. 2016, 7, 981. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Y.; Wang, L.L.; Liu, Y.L.; Zhang, Q.; Wei, Q.P.; Zhang, W.H. Nitric oxide enhances salt tolerance in maize seedlings through increasing activities of proton-pump and Na1/H1 antiport in the tonoplast. Planta 2006, 224, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Albert, B.; Le Cahérec, F.; Niogret, M.F.; Avice, J.C.; Faes, P.; Leport, L.; Bouchereau, A. Nitrogen availability impacts oilseed rape (Brassica napus L.) plant water status and proline production efficiency under water-limited conditions. Planta 2012, 236, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostamza, M.; Chaichi, M.R.; Jahansooz, M.R.; Rahimian Mashhadi, H.; Sharifi, H.R. Effects of water stress and nitrogen fertilizer on multi-cut pearl millet forage yield, nitrogen, and water use efficiency. Commun Soil. Sci. Plant Anal. 2011, 42, 2427–2440. [Google Scholar] [CrossRef]
- Ahmed, M.; Khan, S.; Irfan, M.; Aslam, M.A.; Shabbir, G.; Ahmad, S.; Fahad, S.; Basir, A.; Adnan, M. Effect of Phosphorus on Root Signaling of Wheat under Different Water Regimes. In Global Wheat Production; IntechOpen: London, UK, 2018; pp. 1–29. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Pallaghy, C.K.; Singh, D. Phosphorus nutrition and tolerance of cotton to water stress: I. Seed cotton yield and leaf morphology. Field Crop. Res. 2006, 96, 191–198. [Google Scholar] [CrossRef]
- Garg, B.K.; Burman, U.; Kathju, S. The influence of phosphorus nutrition on the physiological response of moth bean genotypes to drought. J. Plant Nutr. Soil. Sci. 2004, 167, 503–508. [Google Scholar] [CrossRef]
- Zain, N.A.M.; Ismail, M.R.; Puteh, A.; Mahmood, M.; Islam, M.R. Drought tolerance and ion accumulation of rice following application of additional potassium fertilizer. Commun Soil. Sci. Plant Anal. 2014, 45, 2502–2514. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R. Potassium up-regulates antioxidant metabolism and alleviates growth inhibition under water and osmotic stress in wheat (Triticum aestivum L). Protoplasma 2017, 254, 1471–1486. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.; Tomar, N.S.; Shrivastava, M. Potassium induces positive changes in nitrogen metabolism and antioxidant system of oat (Avena sativa L. cultivar Kent). J. Plant Interact. 2015, 10, 211–223. [Google Scholar] [CrossRef]
- Barranco, I.; Tvarijonaviciute, A.; Perez-Patiño, C.; Vicente-Carrillo, A.; Parrilla, I.; Ceron, J.J.; Martinez, E.A.; Rodriguez-Martinez, H.; Roca, J. Effect of potassium fertilizer on the growth, physiological parameters, and water status of Brassica juncea cultivars under different irrigation regimes. PLoS ONE 2021, 16, e0257023. [Google Scholar] [CrossRef]
- Zahoor, R.; Dong, H.; Abid, M.; Zhao, W.; Wang, Y.; Zhou, Z. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Song, C.-J.; Ma, K.-M.; Qu, L.-Y.; Liu, Y.; Xu, X.-L.; Fu, B.-J.; Zhong, J.-F. Interactive effects of water, nitrogen and phosphorus on the growth, biomass partitioning and water-use efficiency of Bauhinia faberi seedlings. J. Arid Environ. 2010, 74, 1003–1012. [Google Scholar] [CrossRef]
- Ata-Ul-Karim, S.T.; Liu, X.; Lu, Z.; Yuan, Z.; Zhu, Y.; Cao, W. In-season estimation of rice grain yield using critical nitrogen dilution curve. Field Crop. Res. 2016, 195, 1–8. [Google Scholar] [CrossRef]
- Hassan, M.J.; Wang, F.; Ali, S.; Zhang, G. Toxic effects of cadmium on rice as affected by nitrogen fertilizer form. Plant Soil. 2005, 277, 359–365. [Google Scholar] [CrossRef]
- Jalloh, M.A.; Chen, J.; Zhen, F.; Zhang, G. Effect of different N fertilizer forms on antioxidant capacity and grain yield of rice growing under Cd stress. J. Hazard Mater. 2009, 162, 1081–1085. [Google Scholar] [CrossRef]
- Li, S.; Zhou, L.; Addo-Danso, S.D.; Ding, G.; Sun, M.; Wu, S.; Lin, S. Nitrogen supply enhances the physiological resistance of Chinese fr plantlets under polyethylene glycol (PEG)-induced drought stress. Sci. Rep. 2020, 10, 7509. [Google Scholar] [CrossRef]
- Zhu, Z.; Gerendas, J.; Bendixen, R.; Schinner, K.; Tabrizi, H.; Sattelmacher, B.; Hansen, U.P. Different tolerance to light stress in N03--and NH4+-grown Phaseolus vulgaris L. Plant Biol. 2000, 2, 558–570. [Google Scholar] [CrossRef]
- Bendixen, R.; Gerendas, J.; Schirmer, K.; Sattelmacher, B.; Hansen, U.P. Difference in zeaxanthin formation in nitrate-and ammonium-grown Phaseolus vulgaris. Physiol. Plant 2001, 111, 255–261. [Google Scholar] [CrossRef]
- Kato, M.C.; Hikosaka, K.; Hirotsu, N.; Makin, A.; Hirose, T. The excess light energy that is neither utilized in photosynthesis nor dissipated by photoprotective mechanisms determines the rate of photoinactivation in photosystem II. Plant Cell Physiol. 2003, 44, 318–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Binghua, L.; Liang, C.; Mingjun, L.; Dong, L.; Yangjun, Z.; Fengwang, M. Interactive effects of water and nitrogen supply on growth, biomass partitioning, and water-use efficiency of young apple trees. Afr. J. Agric. Res. 2012, 7, 978–985. [Google Scholar] [CrossRef]
- Saneoka, H.; Moghaieb, R.E.A.; Premachandra, G.S.; Fujita, K. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relations in Agrostis palustris Huds. Environ. Exp. Bot. 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Saud, S.; Fahad, S.; Yajun, C.; Ihsan, M.Z.; Hammad, H.M.; Nasim, W.; Amanullah, J.; Arif, M.; Alharby, H. Effects of Nitrogen Supply on Water Stress and Recovery Mechanisms in Kentucky Bluegrass Plants. Front. Plant Sci. 2017, 8, 983. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Guo, J.Y.; Wang, G.X.; Yang, L.D.; Yang, Y. Effects of drought and nitrogen addition on photosynthetic characteristics and resource allocation of Abies fabri seedlings in eastern Tibetan plateau. New For. 2012, 43, 505–518. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, S.X.; Zhang, H.; Liang, Z.S. Nitrogen rates and drought stress effects on production, lipid peroxidation and antioxidative enzyme activities in two maize (Zea mays L.) genotypes. J. Agron. Crop. Sci. 2007, 193, 387–397. [Google Scholar] [CrossRef]
- Tarvainen, L.; Näsholm, T. Can adjustments in foliar nitrogen-use efficiency reduce drought stress impacts on boreal trees? Tree Physiol. 2017, 37, 415–417. [Google Scholar] [CrossRef] [Green Version]
- Arun, T.; Upadhyaya, S.D.; Upadhyay, A.; Preeti Sagar, N. Responses of moisture stress on growth, yield and quality of isabgol (Plantago ovata Forsk). J. Agric. Technol. 2012, 8, 563–570. [Google Scholar]
- Song, J.; Wang, Y.; Pan, Y.; Pang, J.; Zhang, X.; Fan, J.; Zhang, Y. The influence of nitrogen availability on anatomical and physiological responses of Populus alba × P. glandulosa to drought stress. BMC Plant Biol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Real, M.M.; Baille, A. Changes in leaf photosynthetic parameters with leaf position and nitrogen content within a rose plant canopy (Rosa hybrida). Plant Cell Environ. 2000, 23, 351–363. [Google Scholar] [CrossRef]
- Li, H.; Li, M.; Luo, J.; Cao, X.Q.L.; Gai, Y.; Jiang, X.N.; Liu, T.X.; Bai, H.; Janz, D.; Polle, A.; et al. N-fertilization has different effects on the growth, carbon and nitrogen physiology, and wood properties of slow- and fast-growing Populus species. J. Exp. Bot. 2012, 63, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Euring, D.; Bai, H.; Janz, D.; Polle, A. Nitrogen-driven stem elongation in poplar is linked with wood modification and gene clusters for stress, photosynthesis and cell wall formation. BMC Plant Biol. 2014, 14, 391. [Google Scholar] [CrossRef] [Green Version]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Silveira, J.A.G.; Costa, R.C.L.; Oliveira, J.T.A. Drought-induced effects and recovery of nitrate assimilation and nodule activity in cowpea plants inoculated with Bradyrhizobium spp. under moderate nitrate level. Braz. J. Microbiol. 2001, 32, 187–194. [Google Scholar] [CrossRef]
- Luo, J.; Li, H.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [Green Version]
- Correia, M.J.; Filomena, F.; Azedo-Silva, J.; Dias, C.; David, M.M.; Barrote, I.; Osorio, M.L.; Osorio, J. Effects of water deficit on the activity of nitrate reductase and contents of sugars, nitrate and free amino acids in the leaves and roots of sunflower and white lupin plants growing under two nutrient supply regimes. Physiol. Plant 2005, 124, 61–70. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C.X.; Su, L.; Li, Y.M.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Vries, F.T.; Brown, C.; Stevens, C.J. Grassland species root response to drought: Consequences for soil carbon and nitrogen availability. Plant Soil. 2016, 409, 297–312. [Google Scholar] [CrossRef]
- Banerjee, P.; Visha Kumari, V.; Nath, R.; Bandyopadhyay, P. Seed priming and foliar nutrition studies on relay grass pea after winter rice in lower Gangetic plain. J. Crop. Weed 2019, 15, 72–78. [Google Scholar] [CrossRef]
- Jin, J.; Lauricella, D.; Armstrong, R.; Sale, P.; Tang, C. Phosphorus application and elevated CO2 enhance drought tolerance in field pea grown in a phosphorus-deficient vertisol. Ann. Bot. 2015, 116, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Sun, X.; Song, D.; Chen, W.; Zhang, A.; et al. Phosphorous Application Improves Drought Tolerance of Phoebe zhennan. Front. Plant Sci. 2017, 8, 1561. [Google Scholar] [CrossRef] [Green Version]
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrup-Andersen, P.; Merino, G.; Hemre, G.-I.; Williams, M. Feeding 2015, 9 billion by 2050—Putting fish back on the menu. Food Sec. 2015, 7, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Song, L.; Zhang, Y.; Zheng, Z.; Liu, D. Arabidopsis PHL2 and PHR1 act redundantly as the key components of the central regulatory system controlling transcriptional responses to phosphate starvation. Plant Physiol. 2016, 170, 499–514. [Google Scholar] [CrossRef] [Green Version]
- Bechtaoui, N.; Rabiu, M.K.; Raklami, A.; Oufdou, K.; Hafidi, M.; Jemo, M. Phosphate-dependent regulation of growth and stresses management in plants. Front. Plant Sci. 2021, 12, 679916. [Google Scholar] [CrossRef]
- Faustino, L.I.; Bulfe, N.M.L.; Pinazo, M.A.; Monteoliva, S.E.; Graciano, C. Dry weight partitioning and hydraulic traits in young Pinus taeda trees fertilized with nitrogen and phosphorus in a subtropical area. Tree Physiol. 2013, 33, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Hansel, F.D.; Amado, T.J.C.; Ruiz Diaz, D.A.; Rosso, L.H.M.; Nicoloso, F.T.; Schorr, M. Phosphorus fertilizer placement and tillage affect soybean root growth and drought tolerance. Agron. J. 2017, 109, 2936–2944. [Google Scholar] [CrossRef] [Green Version]
- Gorny, A.G.; Garczynski, S. Genotypic and nutrition dependent variation in water use efficiency and photosynthetic activity of leaves in winter wheat (Triticum aestivum L.). J. Appl. Genet. 2002, 43, 145–160. [Google Scholar] [PubMed]
- Sardans, J.; Roda, F.; Penuelas, J. Effects of water and a nutrient pulse supply on Rosmarinus of ficinalis growth, nutrient content and flowering in the field. Environ. Exp. Bot. 2005, 53, 1–11. [Google Scholar] [CrossRef]
- Dey, G.; Banerjee, P.; Sharma, R.K.; Maity, J.P.; Etesami, H.; Shaw, A.K.; Huang, Y.H.; Huang, H.B.; Chen, C.Y. Management of phosphorus in salinity-stressed agriculture for sustainable crop production by salt-tolerant phosphate-solubilizing bacteria—A Review. Agronomy 2021, 11, 1552. [Google Scholar] [CrossRef]
- Jones, C.A.; Jacobsen, J.S.; Wraithl, J.M. Response of malt barley to phosphorus fertilization under drought conditions. J. Plant Nutr. 2005, 28, 1605–1617. [Google Scholar] [CrossRef]
- Campbell, C.D.; Sage, R.F. Interactions between the effects of atmospheric CO2 content and P nutrition on photosynthesis in white lupin (Lupinus albus L.). Plant Cell Environ. 2006, 29, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Badgujar, G.; Reddy, V.R.; Fleisher, D.H.; Bunce, J.A. Carbon dioxide diffusion across stomata and mesophyll and photo-biochemical processes as affected by growth CO2 and phosphorus nutrition in cotton. J. Plant Physiol. 2013, 170, 801–813. [Google Scholar] [CrossRef]
- Ajouri, A.H.; Asgedom Becker, M. Seed priming enhances germination and seedling growth of barley under conditions of P and Zn deficiency. J. Plant Nutr. Soil. Sci. 2004, 167, 630–636. [Google Scholar] [CrossRef]
- Sato, A.M.; Catuchi, T.A.; Ribeiro, R.V.; Souza, G.M. The use of network analysis to uncover homeostatic responses of a drought-tolerant sugarcane cultivar under severe water deficit and phosphorus supply. Acta Physiol. Plant 2010, 32, 1145–1151. [Google Scholar] [CrossRef]
- López-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Zhang, L.; Wu, X.; Chen, W.; Song, D.; et al. Phosphorous fertilization alleviates drought effects on Alnus cremastogyne by regulating its antioxidant and osmotic potential. Sci. Rep. 2018, 8, 5644. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A Review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up-regulating antioxidant metabolism and osmolyte accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Yang, J.; Fan, W.; Zheng, S. Mechanisms and regulation of Aluminium induced secretion of organic acid anions from plant roots. J. Zhejiang Univ. Sci. B 2019, 20, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Chen, W.W.; Wang, Y.Y.; Huang, Z.R.; Ye, X.; Chen, L.S.; Yang, L.T. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef] [PubMed]
- Burman, U.; Garg, B.K.; Kathju, S. Effect of phosphorus application on cluster bean under different intensities of water stress. J. Plant Nutr. 2009, 32, 668–680. [Google Scholar] [CrossRef]
- Sardans, J.; Penuelas, J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [Green Version]
- Ge, T.D.; Sun, N.B.; Bai, L.P.; Tong, C.L.; Sui, F.G. Effects of drought stress on phosphorus and potassium uptake dynamics in summer maize (Zea mays) throughout the growth cycle. Act. Physiol. Plant. 2012, 34, 2179–2186. [Google Scholar] [CrossRef]
- Razaq, M.; Zhang, P.; Shen, H.; Salahuddin, S. Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Nasri, N.; Maatallah, S.; Kaddour, R.; Lachâal, M. Effect of salinity on Arabidopsis thaliana seed germination and acid phosphatase activity. Arch. Biol. Sci. 2016, 68, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.C.; Liao, H. Organic acid anions: An effective defensive weapon for plants against aluminum toxicity and phosphorus deficiency in acidic soils. J. Genet. Genom. 2016, 43, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, M.H.M.B.; Hasanuzzaman, M.; Mahmud, J.; Hossain, M.S.; Alam, M.U.; Fujita, M. Explicating physiological and biochemical responses of wheat cultivars under acidity stress: Insight into the antioxidant defence and glyoxalase systems. Physiol. Mol. Biol. Plant 2019, 25, 865–879. [Google Scholar] [CrossRef] [PubMed]
- Goud, V.V.; Konde, N.M.; Mohod, P.V.; Kharche, V.K. Response of chickpea to potassium fertilization on yield, quality, soil fertility and economic in vertisols. Legum. Res. 2012, 37, 311–315. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Huang, X. Effect of potassium supply on plant potassium distribution and growth and leaf photosynthetic capacity of Pyrus pyrifolia. J. Nanjing Agric. Univ. 2017, 40, 60–67. [Google Scholar]
- Jiang, M.Y.; Zhang, J.H. Involvement of plasma membrane NADPH oxidase in abscisic acid- and water stress-induced antioxidant defence in leaves of maize seedlings. Planta 2002, 215, 1022–1030. [Google Scholar] [CrossRef]
- Danial, H.F.; Ewees, M.S.; Moussa, S.A. Significance of influence potassium on the tolerance to induce moisture stress and biological activity of some legume crops grown on a sandy soil Egypt. Egypt J. Soil. Sci. 2010, 43, 180–204. [Google Scholar]
- Jan, A.U.; Hadi, F.; Nawaz, M.A.; Rahman, K. Potassium and zinc increase tolerance to salt stress in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2017, 116, 139–149. [Google Scholar] [CrossRef]
- Banerjee, P.; Ghosh, A.; Visha Kumari, V.; Nath, R. Effect of canopy temperature on physiological processes of grass pea as influenced by seed priming and foliar fertilization. J. Agrometeorol. 2021, 23, 340–343. [Google Scholar] [CrossRef]
- Fooladivanda, Z.; Zadehdelouei, M.H.; Zarifinia, N. Effects of water stress and potassium on quantity traits of two varieties of mungbean (Vigna radiata L.). Cercet. Agron. Mold. 2014, 47, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Soleimanzadeh, H.; Habibi, D.; Ardakani, M.; Paknejad, F.; Rejali, F. Effect of potassium levels on antioxidant enzymes and malondialdehyde content under drought stress in sunflower (Helianthus annuus L.). Am. J. Agric. Biol. Sci. 2010, 5, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant responses and tolerance to abiotic oxidative stress: Antioxidant defence is a key factor. In Crop Stress and Its Management: Perspectives and Strategies; Bandi, V., Shanker, A.K., Shanker, C., Mandapaka, Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 261–316. [Google Scholar]
- Cha-um, S.; Siringam, K.; Juntawong, N.; Kirdmanee, C. Water relations, pigment stabilization, photosynthetic abilities and growth improvement in salt stressed rice plants treated with exogenous potassium nitrate application. Int. J. Plant Prod. 2010, 4, 187–198. [Google Scholar]
- Subbaramamma, P.; Sangamitra, M.; Manjusha, D. Mitigation of drought stress in production of pulses. Int. J. Multidiscip. Adv. Res. Trends 2017, 4, 41–62. [Google Scholar]
- Majeed, S.; Akram, M.; Latif, M.; Ijaz, M.; Hussain, M. Mitigation of drought stress by foliar application of salicylic acid and potassium in mungbean (Vigna radiata L.). Legum. Res. 2016, 39, 208–214. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil. 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Hossen, M.S.; Masud, A.A.C.; Moumita Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Rao, D.S.N.; Naidu, T.C.M.; Rani, Y.A. Effect of foliar nutrition on antioxidant enzymes, photosynthetic rate, dry matter production and yield of mung bean under receding soil moisture condition. Int. J. Pure Appl. Biosci. 2015, 3, 115–123. [Google Scholar]
- Yadav, G.S.; Devi, A.G.; Das, A.; Kandpal, B.; Babu, S.; Das, R.C.; Nath, M. Foliar application of urea and potassium chloride minimizes terminal moisture stress in lentil (Lens culinaris L.) crop. Legume Res. 2019, 44, 627–633. [Google Scholar] [CrossRef]
- El–Mageed, T.A.A.; El-Sherif, A.M.; Ali, M.M.; El-Wahed, M.H.A. Combined effect of deficit irrigation and potassium fertilizer on physiological response, plant water status and yield of soybean in calcareous soil. Arch. Agron. Soil. Sci. 2016, 63, 827–840. [Google Scholar] [CrossRef]
- Banerjee, P.; Venugopalan, V.K.; Nath, R.; Althobaiti, Y.S.; Gaber, A.; Al-Yasi, H.; Hossain, A. Physiology, Growth and Productivity of Spring–Summer Black Gram (Vigna mungo L. Hepper) as Influenced by Heat and Moisture Stresses in Different Dates of Sowing and Nutrient Management Conditions. Agronomy 2021, 11, 2329. [Google Scholar] [CrossRef]
- Halford, N.G. New insights on the effects of heat stress on crops. J. Exp. Bot. 2009, 60, 4215–4216. [Google Scholar] [CrossRef] [Green Version]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of temperature stress by nutrient management in crop plants: A review. J. Soil. Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Ahmad, M.S.A.; Ashraf, M.; Al-Qurainy, F.; Ashraf, M.Y. Alleviation of waterlogging stress in upland cotton (Gossypium hirsutum L.) by exogenous application of potassium in soil and as a foliar spray. Crop. Past. Sci. 2011, 26, 25–38. [Google Scholar] [CrossRef]
- Dwivedi, S.K.; Kumar, S.; Bhakta, N.; Singh, S.K.; Rao, K.K.; Mishra, J.S.; Singh, A.K. Improvement of submergence tolerance in rice through efficient application of potassium under submergence-prone rainfed ecology of Indo-Gangetic Plain. Funct. Plant Biol. 2017, 4, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Luo, W.; Lin, W.; Ma, L.; Kabir, M.H. Model of cation transportation mediated by high-affinity potassium transporters (HKTs) in higher plants. Biol. Proced. Online 2015, 17, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Vanacker, H.; Gornez, L.D.; Harbinson, J. Regulation of photosynthesis and antioxidant metabolism in maize leaves at optimal and chilling temperatures: Review. Plant Physiol. Biochem. 2002, 40, 659–668. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Raper, T.B. Potassium and stress alleviation: Physiological functions and management of cotton. J. Plant Nutr. Soil. Sci. 2013, 176, 331–343. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.R.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Hossen, M.Z.; Hossain, M.A. Physiological mechanisms of exogenous calcium on alleviating salinity-induced stress in rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2019, 25, 611–624. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, X.; Wang, W.; Zhang, S.; Xu, B. Calcium regulates the cell-to-cell water flow pathway in maize roots during variable water conditions. Plant Physiol. Biochem. 2012, 58, 212–219. [Google Scholar] [CrossRef]
- Naeem, M.; Naeem, M.S.; Ahmad, R.; Ihsan, M.Z.; Ashraf, M.Y.; Hussain, Y.; Fahad, S. Foliar calcium spray confers drought stress tolerance in maize via modulation of plant growth, water relations, proline content and hydrogen peroxide activity. Arch. Agron. Soil. Sci. 2018, 64, 116–131. [Google Scholar] [CrossRef]
- Guo, T.R.; Chen, Y.; Zhang, Y.H.; Jin, Y.F. Alleviation of Al toxicity in Barley by addition of calcium. Agric. Sci. China 2006, 5, 828–833. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Réthoré, E.; Pluchon, S.; Ali, N.; Billiot, B.; Yvin, J.C. Calcium Application Enhances Drought Stress Tolerance in Sugar Beet and Promotes Plant Biomass and Beetroot Sucrose Concentration. Int. J. Mol. Sci. 2019, 20, 3777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Meng, Q.W.; Brestic, M.; Olsovska, K.; Yang, X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J. Plant Physiol. 2011, 168, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Xu, G. Low magnesium with high potassium supply changes sugar partitioning and root growth pattern prior to visible magnesium deficiency in leaves of Rice (Oryza sativa L.). Am. J. Plant Sci. 2011, 2, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Mengutay, M.; Ceylan, Y.; Kutman, U.B.; Cakmak, I. Adequate magnesium nutrition mitigates adverse effects of heat stress on maize and wheat. Plant Soil. 2013, 368, 57–72. [Google Scholar] [CrossRef]
- Shabala, S.; Hari, Y. Effects of magnesium availability on the activity of plasma membrane ion transporters and light-induced responses from broad bean leaf mesophyll. Planta 2005, 221, 56–65. [Google Scholar] [CrossRef]
- Thalooth, A.T.; Tawfik Mohamed, H.M. A comparative study on the effect of foliar application of zinc, potassium and magnesium on growth, yield and some chemical constituents of mungbean plants grown under water stress conditions. World J. Agric. Sci. 2006, 2, 37–46. [Google Scholar]
- Dixit, G.; Singh, A.P.; Kumar, A.; Dwivedi, S.; Deeba, F.; Kumar, S.; Pandey, V. Sulfur alleviates arsenic toxicity by reducing its accumulation and modulating proteome, amino acids and thiol metabolism in rice leaves. Sci. Rep. 2015, 5, 16205. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.Z.; Qin, M.L.; Lin, X.Y.; Zhu, Z.W.; Chen, M.X. Sulfur supply reduces cadmium uptake and translocation in rice grains (Oryza sativa L.) by enhancing iron plaque formation, cadmium chelation and vacuolar sequestration. Environ. Pollut. 2018, 238, 76–84. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.H.; Wu, F.H.; He, E.M.; Liu, X.; Shangguan, Z.P.; Zheng, H.L. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawood, M.; Cao, F.; Jahangir, M.M.; Zhang, G.; Wu, F. Alleviation of aluminum toxicity by hydrogen sulfide is related to elevated ATPase, and suppressed aluminum uptake and oxidative stress in barley. J. Hazard. Mater. 2012, 209, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.R.; Zaman, R.; Avice, J.C.; Ourry, A.; Kim, T.H. Sulfur use efficiency is a significant determinant of drought stress tolerance in relation to photosynthetic activity in Brassica napus cultivars. Front Plant Sci. 2016, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Anjum, N.A.; Umar, S.; Ahmad, A.; Iqbal, M.; Khan, N.A. Sulphur protects mustard (Brassica campestris L.) from cadmium toxicity by improving leaf ascorbate and glutathione Sulphur protects mustard from cadmium toxicity. Plant Growth Regu. 2008, 54, 271–279. [Google Scholar] [CrossRef]
- Fatma, M.; Masood, A.; Per, T.S.; Rasheed, F.; Khan, N.A. Interplay between nitric oxide and sulfur assimilation in salt tolerance in plants. Crop. J. 2016, 4, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Mostofa, M.G.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Exogenous calcium alleviates cadmium-induced oxidative stress in rice (Oryza sativa L.) seedlings by regulating the antioxidant defence and glyoxalase systems. Braz. J. Bot. 2015, 39, 393–407. [Google Scholar] [CrossRef]
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.P. Alleviation of cadmium toxicity in Brassica juncea L. (Czern. & Coss.) by calcium application involves various physiological and biochemical strategies. PLoS ONE 2015, 10, e0114571. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Mostofa, M.G.; Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium mitigates arsenic toxicity in rice seedlings by reducing arsenic uptake and modulating the antioxidant defence and glyoxalase systems and stress markers. BioMed Res. Int. 2015, 2015, 340812. [Google Scholar] [CrossRef] [Green Version]
- Parvin, K.; Nahar, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.; Fujita, M. Calcium-Mediated Growth Regulation and Abiotic Stress Tolerance in Plants. In Plant Abiotic Stress to Lerance; Springer: Cham, Switzerland, 2019; pp. 291–331. [Google Scholar]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14, 401–417. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.I.; Park, H.J.; Park, J.H.; Kim, S.; Im, M.Y.; Seo, H.H.; Kim, Y.W.; Hwang, I.; Kim, S.Y. Arabidopsis calcium dependent protein kinase AtCPK32 interacts with ABF4, a transcriptional regulator of abscisic acid-responsive gene expression, and modulates its activity. Plant Physiol. 2005, 139, 1750–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.C.; Zhu, S.Y.; Gao, G.F.; Wang, X.J.; Zhao, R.; Zou, K.Q.; Wang, X.F.; Zhang, X.Y.; Wu, F.Q.; Peng, C.C.; et al. Expression of a grape calcium-dependent protein kinase ACPK1 in Arabidopsis thaliana promotes plant growth and confers abscisic acid-hypersensitivity in germination, post-germination growth, and stomatal movement. Plant Mol. Biol. 2007, 64, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yoo, B.C.; Lee, J.Y.; Pan, W.; Harmon, A.C. Calcium-regulated phosphorylation of soybean serine acetyl transferase in response to oxidative stress. J. Biol. Chem. 2006, 281, 27405–27415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, F. Uptake of Mineral Elements during Abiotic Stress. In Abiotic Stress Responses in Plants; Springer: New York, NY, USA, 2012; pp. 267–281. [Google Scholar]
- Tuna, A.L.; Kaya, C.; Ashraf, M.; Altunlu, H.; Yokas, I.; Yagmur, B. The effects of calcium sulphate on growth, membrane stability and nutrient uptake of tomato plants grown under salt stress. Environ. Exp. Bot. 2007, 59, 173–178. [Google Scholar] [CrossRef]
- Bonilla, I.; El-Hamdaoui, A.; Bolanos, L. Boron and calcium increase Pisum sativum seed germination and seedling development under salt stress. Plant Soil. 2004, 267, 97–107. [Google Scholar] [CrossRef]
- Zehra, A.; Gul, B.; Ansari, R.; Khan, M.A. Role of calcium in alleviating effect of salinity on germination of Phragmites karka seeds. S. Afr. J. Bot. 2012, 78, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Turkmen, O.; Dursun, A.; Turan, M.; Erdinc, C. Calcium and humic acid affect seed germination, growth and nutrient content of tomato (Lycopersicon esculentum L.) seedlings under saline soil conditions. Acta Agric. Scand. B 2004, 54, 168–174. [Google Scholar] [CrossRef]
- Liu, L.-Y.; Wang, M.-Y. Effect of CaCl2 on germination of wheat seeds under salt stress. J. Henan Agric. Sci. 2010, 1, 5–7. [Google Scholar]
- Yao, S.; Chen, S.; Zhao, J.; Xu, D.; Lan, H.; Zhang, F. Effect of three salts on germination and seedling survival of dimorphic seeds of Chenopodium album. Botany 2010, 88, 821–828. [Google Scholar] [CrossRef]
- Patel, N.T.; Vaghela, P.M.; Patel, A.D.; Pandey, A.M. Implications of calcium nutrition on the response of Caesalpinia crista (Fabaceae) to soil salinity. Acta Ecol. Sin. 2011, 31, 24–30. [Google Scholar] [CrossRef]
- Mei, J.; Wang, W.; Peng, S.; Nie, L. Seed pelleting with calcium peroxide improves crop establishment of direct-seeded rice under waterlogging conditions. Sci. Rep. 2017, 7, 4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water deficit stress-induced anatomical changes in higher plants. CR Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Song, W.; Liu, Z.; Zhang, H.; Guo, H.; Shao, H.; Ni, F. The dynamic changing of Ca2+ cellular localization in maize leaflets under drought stress. Comptes Rendus Biol. 2009, 332, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Li, X.; Zhang, L. The effect of calcium chloride on growth, photosynthesis, and antioxidant responses of Zoysia japonica under drought conditions. PLoS ONE 2013, 8, e68214. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, G.; Zhang, X.; Mercier, R.; Leng, Q.; Lawton, M. Co-expression of calcium-dependent protein kinase with the inward rectified guard cell K + channel KAT1 alters current parameters in Xenopus laevis oocytes. Plant Cell Physiol. 2000, 41, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Palta, J.P. Stress interactions at the cellular and membrane levels. Hort. Sci. 2000, 25, 1377–1381. [Google Scholar] [CrossRef]
- Shaikh, F.; Gul, B.; Li, W.Q.; Jing, L.X.; Khan, M.A. Effect of calcium and light on the germination of Urochondra setulosa under different salts. J. Zhejiang Univ. Sci.B 2007, 8, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Nayyar, H.; Kaushal, S.K. Chilling induced oxidative stress in germinating wheat grains as affected by water stress and calcium. Biol. Plant 2002, 45, 601–604. [Google Scholar] [CrossRef]
- Sreedhara, A.; Cowan, J.A. Structural and catalytic roles for divalent magnesium in nucleic acid biochemistry. Biometals 2002, 15, 211–223. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–189. [Google Scholar]
- Hermans, C.N. Verbruggen Physiological characterization of Mg deficiency in Arabidopsis thaliana. J. Exp. Bot. 2005, 56, 2153–2161. [Google Scholar] [CrossRef] [Green Version]
- Hermans, C.; Conn, S.J.; Chen, J.; Xiao, Q.; Verbruggen, N. An update on magnesium homeostasis mechanisms in plants. Metallomics 2013, 5, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Kirkby, E.A. Role of magnesium in carbon partitioning and alleviating photooxidative damage. Physiol. Plant 2008, 133, 692–704. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil. 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Gerendás, J.; Führs, H. The significance of magnesium for crop quality. Plant Soil. 2013, 368, 101–128. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Chen, S.; Hussain, N.; Cong, Y.; Liang, Z.; Chen, K. Magnesium stress signaling in plant: Just a beginning. Plant Signal. Behav. 2015, 10, e992287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Hussain, N.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A. Experimental assessment of salinity tolerance of Ceriops tagal seedlings and saplings from the Indus delta. Pak. Aquat. Bot 2001, 70, 259–268. [Google Scholar]
- Ferreira, R.G.; Tavora, F.J.A.F.; Hernandez, F.F.F. Dry matter partitioning and mineral composition of roots, stems and leaves of guava grown under salt stress conditions. Pesqui. Agropecu. Bras. 2001, 36, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Barhoumi, Z.; Djebali, W.; Smaoui, A.; Chaïbi, W.; Abdelly, C. Contribution of NaCl excretion to salt resistance of Aeluropus littoralis (Willd) Parl. J. Plant Physiol. 2007, 164, 842–850. [Google Scholar] [CrossRef]
- Mahouachi, J. Changes in nutrient concentrations and leaf gas exchange parameters in banana plantlets under gradual soil moisture depletion. Sci. Hort. 2009, 120, 460–466. [Google Scholar] [CrossRef]
- Cao, M.J.; Wang, Z.; Zhao, Q.; Mao, J.L.; Speiser, A.; Wirtz, M.; Xiang, C.B. Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana. Plant J. 2014, 77, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Khan, M.I.R.; Asgher, M.; Fatma, M.; Masood, A.; Syeed, S. Salinity tolerance in plants: Revisiting the role of sulfur metabolites. J. Plant Biochem. Physiol. 2014, 2, 120. [Google Scholar]
- Anjum, N.A.; Gill, R.; Kaushik, M.; Hasanuzzaman, M.; Pereira, E.; Ahmad, I.; Tuteja, N.; Gill, S.S. ATP-sulfurylase, sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 2015, 6, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talukdar, D. Functional interplay between glutathione and hydrogen sulfide in regulation of thiol cascade during arsenate tolerance of common bean (Phaseolus vulgaris L.) genotypes. 3 Biotech. 2015, 5, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, J.; Liu, J.; Liu, T.; Xue, S. Hydrogen sulfide (H2S) signaling in plant development and stress responses. Abiotech 2021, 2, 32–63. [Google Scholar] [CrossRef]
- Min, Y.; Qin, B.-P.; Ma, X.-L.; Wang, P.; Li, M.-L.; Chen, L.L.; Chen, L.-T.; Sun, A.; Wang, Z.; Yin, Y. Foliar application of sodium hydrosulfide (NaHS), a hydrogen sulfide (H2S) donor, can protect seedlings against heat stress in wheat (Triticum aestivum L.). J. Integr. Agric. 2016, 15, 2745–2758. [Google Scholar]
- Zhang, J.; Zhao, Q.Z.; Duan, G.L.; Huang, Y.C. Influence of sulphur on arsenic accumulation and metabolism in rice seedlings. Environ. Exp. Bot. 2011, 72, 34–40. [Google Scholar] [CrossRef]
- Sheng, H.; Zeng, J.; Liu, Y.; Wang, X.; Wang, Y.; Kang, H.; Fan, X.; Sha, L.; Zhang, H.; Zhou, Y. Sulfur mediated alleviation of Mn toxicity in polish wheat relates to regulating Mn allocation and improving antioxidant system. Front. Plant Sci. 2016, 7, 1382. [Google Scholar] [CrossRef] [Green Version]
- Ardıc, M.; Sekmen, A.H.; Tokur, S.; Ozdemir, F.; Turkan, I. Antioxidant responses of chickpea plants subjected to boron toxicity. Plant Biol. 2009, 11, 328–338. [Google Scholar] [CrossRef]
- Michael, P.I.; Krishnaswamy, M. Oxidative stress and antioxidants in cowpea plants subjected to boron and high irradiance stresses. J. Plant Nutr. 2012, 35, 2180–2197. [Google Scholar] [CrossRef]
- Keleş, Y.; Ergün, N.; Öncel, I. Antioxidant enzyme activity affected by high boron concentration in sunflower and tomato seedlings. Commun. Soil. Sci. Plant Anal. 2011, 42, 173–183. [Google Scholar] [CrossRef]
- Zhu, C.Q.; Cao, X.C.; Zhu, L.F.; Hu, W.J.; Hu, A.Y.; Abliz, B.; Zhang, J.H. Boron reduces cell wall aluminum content in rice (Oryza sativa) roots by decreasing H2O2 accumulation. Plant Physiol. Biochem. 2019, 13, 80–90. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; Abdeldaym, E.A.; Abdelaziz, S.M.; El-Sawy, M.B.; Mottaleb, S.A. Synergetic effects of zinc, boron, silicon, and zeolite nanoparticles on confer tolerance in potato plants subjected to salinity. Agronomy 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, R.L.; Jin, J.Y. Effects of zinc and soil moisture on photosynthetic rate and chlorophyll fluorescence parameters of maize. Biol. Plant 2009, 53, 191–194. [Google Scholar] [CrossRef]
- Vazin, F. Effect of zinc sulfate on quantitative and qualitative characteristics of corn (Zea mays) in drought stress. Cercet. Agron. Mold. 2012, 45, 15–24. [Google Scholar] [CrossRef]
- Kheirizadeh Arough, Y.; Seyed Sharifi, R.; Seyed Sharifi, R. Bio fertilizers and zinc effects on some physiological parameters of triticale under water-limitation condition. J. Plant Interact. 2016, 11, 167–177. [Google Scholar] [CrossRef]
- Pavia, I.; Roque, J.; Rocha, L.; Ferreira, H.; Castro, C.; Carvalho, A.; Correia, C. Zinc priming and foliar application enhances photoprotection mechanisms in drought-stressed wheat plants during anthesis. Plant Physiol. Biochem. 2019, 140, 27–42. [Google Scholar] [CrossRef]
- Ullah, A.; Romdhane, L.; Rehman, A.; Farooq, M. Adequate zinc nutrition improves the tolerance against drought and heat stresses in chickpea. Plant Physiol. Biochem. 2019, 143, 11–18. [Google Scholar] [CrossRef]
- Mahmood, A.; Kanwal, H.; Kausar, A.; Ilyas, A.; Akhter, N.; Ilyas, M.; Khalid, H. Seed priming with zinc modulate growth, pigments and yield of chickpea (Cicer arietinum L.) under water deficit conditions. Appl. Ecol. Environ. Res. 2019, 17, 147–160. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Desoky, E.; Elrys, A.S. Influencing of water stress and micronutrients on physio-chemical attributes, yield and anatomical features of Common Bean plants (Phaseolus vulgaris L.). Egypt. J. Agron. 2017, 39, 251–265. [Google Scholar] [CrossRef] [Green Version]
- Sakya, A.T.; Sulistyaningsih, E.; Indradewa, D.; Purwanto, B.H. Stomata character and chlorophyll content of tomato in response to Zn application under drought condition. IOP Conf. Ser. Earth Environ. Sci. 2018, 142, 012033. [Google Scholar] [CrossRef] [Green Version]
- Zafar, S.; Nasri, M.; Moghadam, H.R.T.; Zahedi, H. Effect of zinc and sulfur foliar applications on physiological characteristics of sunflower (Helianthus annuus L.) under water deficit stress. Int. J. Biosci. 2014, 5, 87–96. [Google Scholar]
- Guha, T.; Ravikumar, K.V.G.; Mukherjee, A.; Mukherjee, A.; Kundu, R. Nanopriming with zero valent iron (nZVI) enhances germination and growth in aromatic rice cultivar (Oryza sativa cv. Gobindabhog L.). Plant Physiol. Biochem. 2018, 127, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Al-Amri, N.; Tombuloglu, H.; Slimani, Y.; Akhtar, S.; Barghouthi, M.; Almessiere, M.; Ozcelik, S. Size effect of iron (III) oxide nanomaterials on the growth, and their uptake and translocation in common wheat (Triticum aestivum L.). Ecotoxicol. Environ. Saf. 2020, 194, 110377. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, H.; Amani-Ghadim, A.R.; Matin, A.A.; Ghorbanpour, M. FeO nanoparticles improve physiological and antioxidative attributes of sunflower (Helianthus annuus) plants grown in soil spiked with hexavalent chromium. 3 Biotech 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Alidoust, D.; Isoda, A. Effect of γFe2O3 nanoparticles on photosynthetic characteristic of soybean (Glycine max (L.) Merr.): Foliar spray versus soil amendment. Acta Physiol. Plant. 2013, 35, 3365–3375. [Google Scholar] [CrossRef]
- Pérez-Labrada, F.; López-Vargas, E.R.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Responses of tomato plants under saline stress to foliar application of copper nanoparticles. Plants 2019, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.N.; Rasheed, R.; Ashraf, M.Y.; Ashraf, M.A.; Hussain, I. Exogenously applied zinc and copper mitigate salinity effect in maize (Zea mays L.) by improving key physiological and biochemical attributes. Environ. Sci. Pollut. Res. 2018, 25, 23883–23896. [Google Scholar] [CrossRef]
- O’Neill, M.A.; Ishii, T.; Albershim, P.; Darvill, A.G. Rhamnogalacturonan II: Structure and function of a borate cross-linked cell wall pectic polysaccharite. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Nayak, A.K.; Tripathi, R.; Katara, J.L.; Bihari, P.; Lal, B.; Gautam, P. Boron application improves yield of rice cultivars under high temperature stress during vegetative and reproductive stages. Int. J. Biometeorol. 2018, 62, 1375–1387. [Google Scholar] [CrossRef] [PubMed]
- Venugopalan, V.K.; Nath, R.; Sengupta, K.; Nalia, A.; Banerjee, S.; Sarath Chandran, M.A.; İbrahimova, U.; Dessoky, E.S.; Attia, A.O.; Hassan, M.; et al. The response of lentil (Lens culinaris medik.) to soil moisture and heat stress under different dates of sowing and foliar application of micronutrients. Front. Plant Sci. 2021, 10, 679469. [Google Scholar] [CrossRef] [PubMed]
- Molassiotis, A.; Sotiropoulos, T.; Tanou, G.; Diamantidis, G.; Therios, I. Boron-induced oxidative damage and antioxidant and nucleolytic responses in shoot tips culture of the apple rootstock EM 2006, 9 (Malus domestica Borkh). Environ. Exp. Bot. 2006, 56, 54–62. [Google Scholar] [CrossRef]
- Ilyas, M.; Ayub, G.; Imran Ali Awan, A.; Ahmad, M. Calcium and Boron effect on production and quality of autumn potato crop under chilling temperature. Commun. Soil. Sci. Plant Anal. 2021, 52, 375–388. [Google Scholar] [CrossRef]
- Sayed, S.A. Impacts of boron application on maize plants growing under flooded and unflooded conditions. Biol. Plant 1998, 41, 101–109. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón-Grao, S.; Martínez-Nicolás, J.J.; Alfosea-Simón, M.; Liu, C.; Chatzissavvidis, C.; Cámara-Zapata, J.M. Multiple stresses occurring with boron toxicity and deficiency in plants. J. Hazard. Mater. 2020, 397, 122713. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.; Rasheed, S.; Kobayashi, T.; Seki, M.; Nishizawa, N.K. Regulating subcellular metal homeostasis: The key to crop improvement. Front. Plant Sci. 2016, 2004, 1192. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K. Cold stress tolerance mechanisms in plants. A Review. Agron. Sustain. Dev. 2010, 30, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Chen, P.; Yan, Y.; Bao, C.; Li, X.; Wang, L.; Shen, X.; Li, H.; Liu, X.; Niu, C.; et al. An atypical R2R3 MYB transcription factor increases cold hardiness by CBF-dependent and CBF-independent pathways in apple. New Phytol. 2018, 218, 201–218. [Google Scholar] [CrossRef]
- Nadeem, F.; Farooq, M. Application of micronutrients in rice-wheat cropping system of South Asia. Rice Sci. 2019, 26, 356–371. [Google Scholar] [CrossRef]
- Liu, C.; Wu, Y.; Wang, X. bZIP transcription factor OsbZIP52/RISBZ5: A potential negative regulator of cold and drought stress response in rice. Planta 2012, 235, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Jiang, M.; Zhang, H.; Li, R. Zinc oxide nanoparticles alleviate chilling stress in rice (Oryza sativa L.) by regulating antioxidative system and chilling response transcription factors. Molecules 2021, 26, 2196. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.; Rashid, A.; Arif, M.; Yunas, M. Alleviating Micronutrient Deficiencies in Alkaline Soils of the North-West Frontier Province of Pakistan: On-Farm Seed Priming with Zinc in Wheat and Chickpea. In Micronutrients in South and South East Asia, International Centre for Integrated Mountain Development; Andersen, P., Tuladhar, J.K., Karki, K.B., Maskey, S.L., Eds.; International Centre for Integrated Mountain Development (ICIMOD): Kathmundu, Nepal, 2005; pp. 143–151. [Google Scholar]
- Umair Hassan, M.; Aamer, M.; Umer Chattha, M.; Haiying, T.; Shahzad, B.; Barbanti, L.; Guoqin, H. The critical role of zinc in plants facing the drought stress. Agriculture 2020, 10, 396. [Google Scholar] [CrossRef]
- Daneshbakhsh, B.; Khoshgoftarmanesh, A.H.; Shariatmadari, H.; Cakmak, I. Effect of zinc nutrition on salinity-induced oxidative damages in wheat genotypes differing in zinc deficiency tolerance. Acta Physiol. Plant 2013, 35, 881–889. [Google Scholar] [CrossRef]
- Sohrabi, Y.; Heidari, G.; Esmailpoor, B. Effect of salinity on growth and yield of Desi and Kabuli chickpea cultivars. Pak. J. Biol. Sci. 2008, 11, 664–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, A.; Farooq, M.; Rehman, A.; Hussain, M.; Siddique, K.H. Zinc nutrition in chickpea (Cicer arietinum): A review. Crop. Pasture Sci. 2020, 71, 199–218. [Google Scholar] [CrossRef]
- Banti, V.; Giuntoli, B.; Gonzali, S.; Loreti, E.; Magneschi, L.; Novi, G.; Paparelli, E.; Parlanti, S.; Pucciariello, C.; Santaniello, A.; et al. Low oxygen response mechanisms in green organisms. Int. J. Mol. Sci. 2013, 14, 4734–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porwal, P.; Sonkar, S.; Singh, A.K. Plant Stress Enzymes Nanobiotechnology. In Nanobiotechnology; Springer: Cham, Switzerland, 2021; pp. 327–348. [Google Scholar]
- Kumar, P.; Tewari, R.K.; Sharma, P.N. Sodium nitroprusside-mediated alleviation of iron deficiency and modulation of antioxidant responses in maize plants. AoB Plants 2010, 2010, plq002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venugopalan, V.K.; Roy, A.; Vijayan, R.; Banerjee, P.; Verma, V.C.; Nalia, A.; Pramanik, M.; Mukherjee, B.; Ghosh, A.; Reja, M.; et al. Drought and heat stress in cool-season food legumes in sub-tropical regions: Consequences, adaptation, and mitigation strategies. Plants 2021, 10, 1038. [Google Scholar]
- Balk, J.; Pilon, M. Ancient and essential: The assembly of iron–sulfur clusters in plants. Trends Plant Sci. 2011, 16, 218–226. [Google Scholar] [CrossRef]
- Khan, M.A.; Ansari, R.; Ali, H.; Gul, B.; Nielsen, B.L. Panicum turgidum, a potentially sustainable cattle feed alternative to maize for saline areas. Agric. Ecosyst. Environ. 2009, 129, 542–546. [Google Scholar] [CrossRef]
- Ghasemi, S.; Khoshgoftarmanesh, A.H.; Afyuni, M.; Hadadzadeh, H. Iron (II)–amino acid chelates alleviate salt-stress induced oxidative damages on tomato grown in nutrient solution culture. Sci. Hortic. 2014, 165, 91–98. [Google Scholar] [CrossRef]
- Pourgholam, M.O.; Nemati, N.A.; Oveysi, M.E. Effect of zinc and iron under the influence of drought on yield and yield components of rapeseed (Brassica napus). Ann. Biol. Res. 2013, 4, 186–189. [Google Scholar]
- Kumari, V.V.; Banerjee, P.; Nath, R.; Sengupta, K.; Chandran, M.A.S.; Kumar, R. Effect of foliar spray on phenology and yield of Lentil sown on different dates. J. Crop. Weed 2019, 15, 54–58. [Google Scholar] [CrossRef]
- Kumari, V.V.; Nath, R.; Sengupta, K.; Banerjee, S.; Dutta, D.; Karmakar, S. Effect of date of sowing and foliar spray of micronutrients on growth, phenology, yield and seed quality of Lentil (Lens culinaris) in New Alluvial Zone of West Bengal. Indian J. Agric. Sci. 2021, 91, 573–576. [Google Scholar]
- Rotaru, V. The effect of phosphorus and iron on plant growth and nutrient status of two soybean (Glycine max L.) cultivars under suboptimal water regime of soil. Lucr. Stiintifice Supl. Agron. 2011, 54, 11–16. [Google Scholar]
- Baghizadeh, A.; Shahbazi, M. Effect of Zn and Fe foliar application on yield, yield components and some physiological traits of cumin (Cuminum cyminum) in dry farming. Int. J. Agron. Plant Prod. 2013, 4, 3231–3237. [Google Scholar]
- Afshar, R.M.; Hadi, H.; Pirzad, A. Effect of nano-iron on the yield and yield component of cowpea (Vigna unguiculata) under end season water deficit. Int. J. Agric. Sci. 2013, 3, 27. [Google Scholar]
- Hasanpour, H.; Maali-Amir, R.; Zeinali, H. Effect of TiO2 nanoparticles on metabolic limitations to photosynthesis under cold in chickpea. Russ. J. Plant Physiol. 2015, 62, 779–787. [Google Scholar] [CrossRef]
- Atar, N.; Eren, T.; Yola, M.L.; Karimi-Maleh, H.; Demirdögen, B. Magnetic iron oxide and iron oxide@ gold nanoparticle anchored nitrogen and sulfur-functionalized reduced graphene oxide electrocatalyst for methanol oxidation. RSC Adv. 2015, 5, 26402–26409. [Google Scholar] [CrossRef]
- Dufey, I.; Gheysens, S.; Ingabire, A.; Lutts, S.; Bertin, P. Silicon application in cultivated rices (Oryza sativa L and Oryza glaberrima Steud) alleviates iron toxicity symptoms through the reduction in iron concentration in the leaf tissue. J. Agron. Crop. Sci. 2014, 200, 132–142. [Google Scholar] [CrossRef]
- Smethurst, C.F.; Garnett, T.; Shabala, S. Nutritional and chlorophyll fluorescence responses of lucerne (Medicago sativa) to waterlogging and subsequent recovery. Plant Soil. 2005, 270, 31–45. [Google Scholar] [CrossRef]
- Syuhada, N.; Jahan, M.S.; Khandaker, M.M.; Nashriyah, M.; Khairi, M.; Nozulaidi, M.; Razali, M.B. Application of copper increased corn yield through enhancing physiological functions. Aust. J. Basic. Appl. Sci. 2014, 8, 282–286. [Google Scholar]
- Yamasaki, H.; Pilon, M.; Shikanai, T. How do plants respond to copper deficiency? Plant Signal. Behav. 2008, 3, 231–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilon, M.; Abdel-Ghany, S.E.; Cohu, C.M.; Gogolin, K.A.; Ye, H. Copper cofactor delivery in plant cells. Curr. Opin. Plant Biol. 2006, 9, 256–263. [Google Scholar] [CrossRef]
- Kamat, J.P.; Boloor, K.K.; Devasagayam, T.P. Chlorophyllin as an effective antioxidant against membrane damage in vitro and ex vivo. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2000, 1487, 113–127. [Google Scholar] [CrossRef]
- Zhang, X.; Goatley, M.; Conner, J.; Wilkins, M.; Teshler, I.; Liu, J.; Fefer, M.; Ckurshumova, W. Copper chlorophyllin impacts on growth and drought stress tolerance of tomato plants. Hort. Sci. 2019, 54, 2195–2201. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Ckurshumova, W.; Fefer, M.; Liu, J.; Uddin, W.; Rosa, C. A plant based modified biostimulant (copper chlorophyllin), mediates defence response in Arabidopsis thaliana under salinity stress. Plants 2021, 10, 625. [Google Scholar] [CrossRef]
- Hejazi, M.M.; Shariatmadari, H.; Khoshgoftarmanesh, A.H.; Dehghani, F. Copper effects on growth, lipid peroxidation, and total phenolic content of rosemary leaves under salinity stress. J. Agric. Sci. Technol. 2019, 54, 2195–2201. [Google Scholar]
- Jacobson, A.; Doxey, S.; Potter, M.; Adams, J.; Britt, D.; McManus, P.; McLean, J.; Anderson, A. Interactions between a plant probiotic and nanoparticles on plant responses related to drought tolerance. Ind. Biotechnol. 2018, 14, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.; Waheed, Z.; Naseem, A. Nanotechnology and Abiotic Stresses. In Nanoagronomy; Javad, S., Ed.; Springer: Cham, Switzerland, 2020; pp. 37–52. [Google Scholar]
- Sarma, B.; Devi, P.; Gogoi, N.; Devi, Y.M. Effects of cobalt induced stress on Triticum aestivum L. crop. Asian J. Agric. Biol. 2014, 2, 137–147. [Google Scholar]
- Gad, N.; El–Metwally, I.M. Chemical and physiological response of maize to salinity using cobalt supplement. Int. J. Chem. Tech. Res. 2015, 8, 45–52. [Google Scholar]
- Jayakumar, K.; Vijayarengan, P.; Chang-Xing, Z.; Jaleel, C.A. Soil applied cobalt alters the nodulation, leg-haemoglobin content and antioxidant status of Glycine max (L.) Merr. Colloids Surf. B Biointerfaces 2008, 67, 272–275. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced up-regulation of the antioxidant defence and methylglyoxal detoxification system reduces salinity-induced damage in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Marxen, A.; Klotzbücher, T.; Jahn, R. Interaction between silicon cycling and straw decomposition in a silicon deficient rice production system. Plant Soil. 2016, 398, 153–163. [Google Scholar] [CrossRef]
- Gong, H.; Zhu, X.; Chen, K.; Wang, S.; Zhang, C. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 2005, 169, 313–321. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon improves water use efficiency in maize plants. J. Plant Nutr. 2004, 27, 1457–1470. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.; Tran, L.S.P. Impacts of priming with silicon on the growth and tolerance of maize plants to alkaline stress. Front. Plant Sci. 2016, 7, 243. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxova´, M.; Lux, A. Application of silicon enhanced drought tolerance in Sorghum bicolour. Physiol. Plant 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Iram, A.; Awan, T.H.; Tanveer, A.; Akbar, N.; Saleem, M.F.; Safdar, M.E. Optimization of cobalt and nitrogen for improving seed yield, protein content and nitrogen use efficiency in mungbean. J. Environ. Agric. 2017, 2, 173–179. [Google Scholar]
- Lwalaba, J.W.; Louisa, L.T.; Zvobgoa, G.; Richmonda, M.E.A.; Fua, L.; Naza, S.; Mwambaa, M.; Mundendeb, R.P.M.; Zhang, G. Physiological and molecular mechanisms of cobalt and copper interaction in causing phyto-toxicity to two barley genotypes difering in Co tolerance. Ecotoxicol. Environ. Saf. 2020, 187, 109866. [Google Scholar] [CrossRef] [PubMed]
- Awomi, T.A.; Singh, A.K.; Kumar, M.; Bordoloi, L.J. Effect of phosphorus, molybdenum and cobalt nutrition on yield and quality of mungbean (Vigna radiata L.) in acidic soil of Northeast India. Indian J. Hill Farm. 2012, 25, 22–26. [Google Scholar]
- Minz, A.; Sinha, A.K.; Kumar, R.; Kumar, B.; Deep, K.P.; Kumar, S.B. A Review on Importance of Cobalt in Crop Growth and Production. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2978–2984. [Google Scholar]
- DalCorso, G.; Manara, A.; Piasentin, S.; Furini, A. Nutrient metal elements in plants. Metallomics 2014, 6, 1770–1788. [Google Scholar] [CrossRef] [PubMed]
- Arif, N.; Yadav, V.; Singh, S.; Singh, S.; Ahmad, P.; Mishra, R.K.; Sharma, S.; Tripathi, D.K.; Dubey, N.K.; Chauhan, D.K. Influence of High and Low Levels of Plant-Beneficial Heavy Metal Ions on Plant Growth and Development. Front. Environ. Sci. 2016, 4, 69. [Google Scholar] [CrossRef]
- Srivastava, S.; Shukla, A.K. Differential response of blackgram towards heavy metal stress. Environ. Pollut. Prot. 2016, 1, 89–96. [Google Scholar]
- Banerjee, P.B.; Hattacharya, P. Investigating cobalt in soil-plant-animal-human system: Dynamics, Impact and Management. J. Soil. Sci. Plant Nutr. 2021, 21, 2339–2354. [Google Scholar] [CrossRef]
- Jaleel, A.; Jayakumar, K.; Chang-Xing, Z.; Azooz, M.M. Antioxidant potentials protect Vigna radiata (L.) Wilczek plants from soil cobalt stress and improve growth and pigment composition. Plant Omics 2009, 2, 120–126. [Google Scholar]
- El-Baz, E.E.T.; Lo’ay, A.A.; Ibrahium, E.G.; El-Deeb, M.R.I. Effect of cobalt and some vitamins as foliar application treatments on productivity and quality of williams banana cultivar. J. Plant Prod. 2016, 7, 777–786. [Google Scholar] [CrossRef]
- Banerjee, P.; Mukherjee, B.; Venugopalan, V.K.; Nath, R.; Chandran, M.A.S.; Dessoky, E.S.; Ismail, I.A.; El-Hallous, E.I.; Hossain, A. Thermal response of spring–summer-grown blackgram (Vigna mungo L. Hepper) in Indian subtropics. Atmosphere 2021, 12, 1489. [Google Scholar] [CrossRef]
- Gad, N.; Kandil, H. Influence of cobalt on phosphorus uptake, growth and yield of tomato. Agric. Biol. J. N. Am. 2010, 1, 1069–1075. [Google Scholar] [CrossRef]
- Kalavrouziotis, I.K.; Koukoulakis, P.H.; Manouris, G.; Papadopoulos, A.H. Interactions between cadmium, lead, cobalt, and nickel in broccoli, irrigated with treated municipal wastewater. Eur. Water 2009, 25, 13–23. [Google Scholar]
- Chmielowska-Bąk, J.; Lefèvre, I.; Lutts, S.; Kulik, A.; Deckert, J. Effect of cobalt chloride on soybean seedlings subjected to cadmium stress. Acta Soc. Bot. Pol. 2014, 83, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Mozafariyan, M.; Kamelmanesh, M.M.; Hawrylak-Nowak, B. Ameliorative effect of selenium on tomato plants grown under salinity stress. Arch. Agron. Soil. Sci. 2016, 62, 1368–1380. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Plant Oxidative Stress: Biology, Physiology and Mitigation. Plants 2022, 11, 1185. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium in higher plants: Physiological role, antioxidant metabolizm and abiotic stress tolerance. J. Plant Sci. 2010, 5, 354–375. [Google Scholar] [CrossRef] [Green Version]
- Sieprawska, A.; Kornaś, A.; Filek, M. Involvement of selenium in protective mechanisms of plants under environmental stress conditions—Review. Acta Biol. Crac. Ser. Bot. 2015, 57, 9–20. [Google Scholar] [CrossRef]
- Singhal, R.K.; Fahad, S.; Kumar, P.; Choyal, P.; Javed, T.; Jinger, D.; Prabha, S.; Debanjana, S.; Prathibha, M.D.; Bandana, B.; et al. Beneficial elements: New Players in improving nutrient use efficiency and abiotic stress tolerance. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, S.; Kaur, S.; Nayyar, H. Selenium in agriculture: A nutrient or contaminant for crops? Arch. Agron. Soil. Sci. 2014, 60, 1593–1624. [Google Scholar] [CrossRef]
- Hartikainen, H.; Xue, T.; Piironen, V. Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil. 2000, 225, 193–200. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Shanker, A.K.; Sheeba, J.A.; Devi, D.D.; Bangarusamy, U. Selenium-An antioxidative protectant in soybean during senescence. Plant Soil. 2005, 272, 77–86. [Google Scholar] [CrossRef]
- Del Pino, A.M.; Regni, L.; D’Amato, R.; Di Michele, A.; Proietti, P.; Palmerini, C.A. Persistence of the effects of Se-fertilization in olive trees over time, monitored with the cytosolic Ca2+ and with the germination of pollen. Plants 2021, 10, 2290. [Google Scholar] [CrossRef] [PubMed]
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R.; Janeeshma, E.; Puthur, J.T.; Aliniaeifard, S.; Chauhan, D.K.; et al. Metal/Metalloid-Based Nanomaterials for Plant Abiotic Stress Tolerance: An Overview of the Mechanisms. Plants 2022, 11, 316. [Google Scholar] [CrossRef]
- Shekari, F.; Abbasi, A.; Mustafavi, S.H. Effect of silicon and selenium on enzymatic changes and productivity of dill in saline condition. J. Saudi. Soc. Agric. Sci. 2017, 27, R713–R715. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Rahman, K.; Sathi, K.S.; Alam, M.M.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Supplemental selenium and boron mitigate salt-induced oxidative damages in Glycine max L. Plants 2021, 10, 2224. [Google Scholar] [CrossRef]
- Valadabadi, S.A.; Shiranirad, A.H.; Farahani, H.A. Ecophysiological influences of zeolite and selenium on water deficit stress tolerance in different rapeseed cultivars. J. Ecol. Nat. Environ. 2010, 2, 154–159. [Google Scholar]
- Habibi, G. Selenium ameliorates salinity stress in Petroselinum crispum by modulation of photosynthesis and by reducing shoot Na accumulation. Russ. J. Plant Physiol. 2015, 64, 368. [Google Scholar] [CrossRef]
- Germ, M.; Stibilj, V.; Kreft, I. Metabolic importance of selenium for plants. Eur. J. Plant Sci. Biotech. 2007, 1, 91–97. [Google Scholar]
- Hawrylak-Nowak, B.; Matraszek, R.; Szymanska, M. Selenium modifies the effect of short-term chilling stress on cucumber plants. Biol. Trace. Elem. Res. 2010, 138, 307–315. [Google Scholar] [CrossRef]
- Hu, K.; Zhang, L.; Wang, J.; You, Y. Influence of selenium on growth, lipid peroxidation and antioxidative enzyme activity in melon (Cucumis melo L.) seedlings under salt stress. Acta Soc. Bot. Pol. 2013, 82, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.D.; Mapeli, A.M.; Antunes, W.C.; Barros, R.S. A dual role of selenium in the growth control of seedlings of Stylosanthes humilis. Agric. Sci. 2011, 2, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and growth-promoting effect of selenium on senescing lettuce. Plant Soil. 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Nair, R.; Giraldo, J.P.; Prasad, P.V.V. Cerium oxide nanoparticles decrease drought-induced oxidative damage in sorghum leading to higher photosynthesis and grain yield. ACS Omega 2018, 3, 14406–14416. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, M.; Abolghasemi, R.; da Silva, J.A.T. Low and high temperature stress affect the growth characteristics of tomato in hydroponic culture with Se and nano-Se amendment. Sci. Hortic. 2014, 178, 231–240. [Google Scholar] [CrossRef]
- Yin, H.; Qi, Z.; Li, M.; Jalal, G.; Chu, X.; Zhou, J. Selenium forms and methods of application differentially modulate plant growth, photosynthesis, stress tolerance, selenium content and speciation in Oryza sativa L. Ecotoxicol. Environ. Saf. 2019, 169, 911–917. [Google Scholar] [CrossRef]
- Nawaz, F.; Ahmad, R.; Ashraf, M.Y.; Waraich, E.A.; Khan, S.Z. Effect of selenium foliar spray on physiological and biochemical processes and chemical constituents of wheat under drought stress. Ecotoxicol. Environ. Saf. 2015, 113, 191–200. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-Mageed, T.A.; Abdelkhalik, A.; Hemida, K.A.; Abdurrahman, H.A.; Howladar, S.M.; Leilah, A.A.A.; Rady, M.O.A. Selenium Modulates Antioxidant Activity, Osmoprotectants, and Photosynthetic Efficiency of Onion under Saline Soil Conditions. Agronomy 2021, 11, 855. [Google Scholar] [CrossRef]
- Kápolna, E.; Laursen, K.H.; Husted, S.; Larsen, E.H. Bio-fortification and isotopic labelling of Se metabolites in onions and carrots following foliar application of Se and 77 Se. Food Chem. 2012, 133, 650–657. [Google Scholar] [CrossRef]
- Jó'zwiak, W.; Politycka, B. Effect of selenium on alleviating oxidative stress caused by a water deficit in cucumber roots. Plants 2019, 8, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.A.; Akbar, A.; Parveen, A.; Rasheed, R.; Hussain, I. Plant physiology and biochemistry phenological application of selenium differentially improves growth, oxidative defense and ion homeostasis in maize under salinity stress. Plant Physiol. Biochem. 2018, 123, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Roychoudhury, A. Role of Beneficial Trace Elements in Salt Stress Tolerance of Plants. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer Nature Ltd.: Singapore, 2018; pp. 377–390. [Google Scholar] [CrossRef]
- Desoky, E.M.; Abdel-Rahman, M.; Mohamed, F.; Abo, E.M.; Esayed, M.; Safaa, M.A.A.I.A.; Mohamed, F.A.; Mohamed, F.R.; Seham, A.I. Physiological and Biochemical Mechanisms of Exogenously Applied Selenium for Alleviating Destructive Impacts Induced by Salinity Stress in Bread Wheat. Agronomy 2021, 11, 926. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Beneficial effects of exogenous selenium in cucumber seedlings subjected to salt stress. Biol. Trace. Elem. Res. 2009, 132, 259–269. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Réthoré, E.; Yvin, J.C.; Hosseini, S.A. The Regulatory Role of Silicon in Mitigating Plant Nutritional Stresses. Plants 2020, 9, 1779. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.A.; Bhat, B.A.; Yousuf, H.; Islam, S.T.; Raza, A.; Rizvi, M.A.; Charagh, S.; Albaqami, M.; Sofi, P.A.; Zargar, S.M. Multidimensional Role of Silicon to Activate Resilient Plant Growth and to Mitigate Abiotic Stress. Front. Plant Sci. 2022, 13, 819658. [Google Scholar] [CrossRef] [PubMed]
- Balakhnina, T.; Borkowska, A. Effects of silicon on plant resistance to environmental stresses: A Review. Int. Agrophys 2013, 27, 225–232. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, K.A.A.; Mazrou, Y.S.A.; Hafez, Y.M. Silicon foliar application mitigates salt stress in sweet pepper plants by enhancing water status, photosynthesis, antioxidant enzyme activity and fruit yield. Plants 2020, 9, 733. [Google Scholar] [CrossRef]
- Ahmed, M.; Fayyaz Ul, H.; Qadeer, U.; Aslam, M.A. Silicon application and drought tolerance mechanism of sorghum. Afr. J. Agric. Res. 2011, 6, 594–607. [Google Scholar]
- Gagoonani, S.; Enteshari, S.; Delavar, K.; Behyar, M. Interactive effects of silicon and aluminum on the malondialdehyde (MDA), proline, protein and phenolic compounds in Borago officinalis L. J. Med. Plant Res. 2011, 5, 5818–5827. [Google Scholar]
- Ma, J.; Cai, H.; He, C.; Zhang, W.; Wang, L. A hemicellulose-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells. New Phytol. 2015, 206, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil. Sci. Plant Nutr. 2015, 50, 11–18. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, P.; Chen, D.; Yin, L.; Li, H.; Deng, X. Silicon enhanced salt tolerance by improving the root water uptake and decreasing the ion toxicity in cucumber. Front. Plant Sci. 2015, 6, 759. [Google Scholar] [CrossRef] [Green Version]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.V.; Sanglard, L.M.V.P.; Reis, J.V.; Detmann, E.; Rodrigues, F.Á.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon nutrition increases grain yield, which, in turn, exerts a feed-forward stimulation of photosynthetic rates via enhanced mesophyll conductance and alters primary metabolism in rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Detmann, K.; Araújo, W.; Martins, S.; Fernie, A.R.; DaMatta, F. Metabolic alterations triggered by silicon nutrition: Is there a signaling role for silicon? Plant Signal. Behav. 2013, 8, e22523. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.X.; Xu, X.B.; Hu, Y.H.; Han, W.H.; Yin, J.L.; Li, H.L.; Gong, H.J. Silicon improves salt tolerance by increasing root water uptake in Cucumis sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, K.; Hattori, T.; An, P.; Tsuji, W.; Eneji, A.E.; Kobayashi, S.; Kawamura, Y.; Tanaka, K.; Inanaga, S. Effect of silicon application on sorghum root responses to water stress. J. Plant Nutr. 2010, 34, 71–82. [Google Scholar] [CrossRef]
- Eshi, Y.; Ezhang, Y.; Ehan, W.; Efeng, R.; Ehu, Y.; Eguo, J.; Egong, H. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef]
- Al-Aghabary, K.; Zhu, Z.; Shi, Q.H. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Ahmed, M.; Qadeer, U.; Ahmed, Z.I.; Fayyaz-Ul, H. Improvement of wheat (Triticum aestivum) drought tolerance by seed priming with silicon. Arch. Acker Pflanzenbau Bodenkd. 2016, 62, 299–315. [Google Scholar] [CrossRef]
- Banerjee, P.; Venugopalan, V.K.; Nath, R.; Chakraborty, P.K.; Gaber, A.; Alsanie, W.F.; Raafat, B.M.; Hossain, A. Seed priming and foliar application of nutrients influence the productivity of relay grass pea (Lathyrus sativus L.) through accelerating the photosynthetically active radiation (PAR) use efficiency. Agronomy 2022, 12, 1125. [Google Scholar] [CrossRef]
- Pathak, J.; Ahmed, H.; Kumari, N.; Pandey, A.; Rajneesh Sinha, R. Role of Calcium and Potassium in Amelioration of Environmental Stress in Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 535–562. [Google Scholar] [CrossRef]
- Venugopalan, V.K.; Nath, R.; Sengupta, K.; Pal, A.K.; Banerjee, S.; Banerjee, P.; Chandran, M.A.S.; Roy, S.; Sharma, L.; Hossain, A.; et al. Foliar spray of micronutrients alleviates heat and moisture stress in lentil (Lens culinaris Medik) grown under rainfed field conditions. Front. Plant Sci. 2022, 13, 47743. [Google Scholar] [CrossRef]
- Dias, A.S.; Lido, F.C.; Ramalho, J.C. Heat stress in Triticum: Kinetics of Fe and Mn accumulation. Braz. J. Plant Physiol. 2009, 21, 153–164. [Google Scholar] [CrossRef]
- Akeel, A.; Jahan, A. Role of cobalt in plants: Its stress and alleviation. In Contaminants in Agriculture; Naeem, M., Ansari, A., Gill, S., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]






{kind=link}
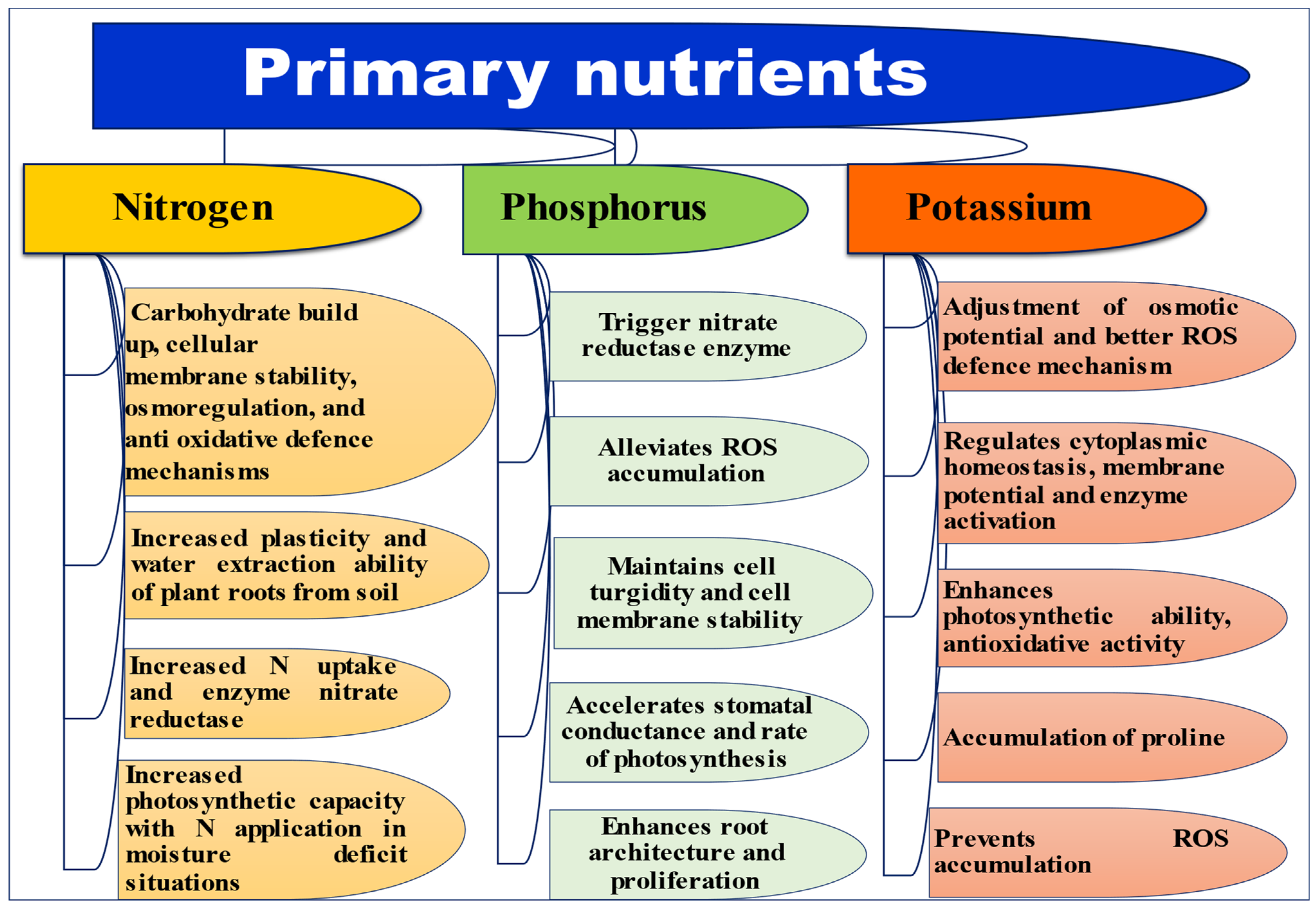{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abiotic Stress | Effects on Plant | References |
|---|---|---|
| Heat | Causes water scarcity, osmotic and oxidative stress that enhances ROS production, protein misfolding and denaturation. Foliar senescence and leaf discoloration, reduced CO2 fixation and disturbed ion transport. | [14,15,16] |
| Chilling | Leads to osmotic and oxidative stress, nutritional imbalance. Accumulation of ROS, inhibition of enzyme activities, and reduced photosynthetic activity. | [17,18] |
| Salinity | Responsible for water scarcity and ionic imbalance. Osmotic and oxidative stress that enhanced ROS production, restricted uptake and translocation of water and mineral nutrients, decreased stomata opening and reduced photosynthesis. | [19,20] |
| Drought (Water deficit) | Causes osmotic and oxidative stress. Increased ROS production and ion leakage. Decrease in absorption and translocation of mineral nutrients. Protein denaturation, loss of enzyme activities | [21] |
| Flooding/water-logging | Leads to oxidative stress and increased ROS production. Reduced gaseous exchange and photosynthetic activity due to lower chlorophyll content. | [22,23] |
| Light/radiation | Oxidative stress, increased ROS production and oxidative damage, reduced photosynthetic activity, and chlorophyll degradation. | [24] |
| Macronutrient | Tested Crop | Mechanisms Related to Stress Alleviation | References |
|---|---|---|---|
| N | Rice | Strengthening root system, improved xylem transport, | [30] |
| Wheat | upregulated photosynthesis, chloroplast fluorescence, enzymatic functions, relative leaf water content, nutrients uptake, lower lipid peroxidation, improved antioxidant defence mechanism in terms of SOD and POX | [31] | |
| Maize | Higher leaf expansion, reduced Na uptake, delayed cell senescence, stomatal regulation | [32] | |
| Rapeseed | Improved plant water status, greater proline accumulation | [33] | |
| Forage pearl millet | Escalated water use efficiency, profuse branching | [34] | |
| P | Wheat | Better root and shoot extension, carbohydrate transport, increased nutrients and water use efficiencies | [35] |
| Cotton | Improved relative leaf water status | [36] | |
| Moth bean | Enhanced nutrients uptake, root hydraulic conductance, modified root development, leaf moisture content | [37] | |
| K | Rice | Better shoot development, greater synthesis of osmolytes | [38] |
| Wheat | Improved antioxidant enzymatic functions | [39] | |
| Oat | Augmented nitrogen metabolism and antioxidant defence system | [40] | |
| Indian mustard | Higher leaf area expansion, membrane stability, leaf water status, modified antioxidants activity regarding CAT, POX, APX, SOD | [41] | |
| Cotton | Enhanced photosynthetic mechanism, improved carbohydrate metabolism | [42] |
| Secondary Nutrient | Tested Crop | Mechanisms Related to Stress Alleviation | References |
|---|---|---|---|
| Ca | Rice | Improved germination characters, shoot and root development, enhanced leaf chlorophyll and proline contents, oppression of ROS by stimulation of CAT and POX | [127] |
| Maize | Water regulation at cellular level and root development | [128] | |
| Improved growth, osmotic relation and proline content, reduced H2O2 activity | [129] | ||
| Barley | Alleviation of Al toxicity, ROS suppression and antioxidative enzymes | [130] | |
| Sugarbeet | Extensive leaf coverage, chlorophyll content, carbohydrate accumulation, reduced oxidative stress by escalating glutathione and free polyamine putrescine pools while reducing amino acid gamma-aminobutyric acid levels | [131] | |
| Tobacco | Accelerated photosynthetic activity, stomatal conductance, improved thermostability of different oxygen-evolving complex and less accumulation of ROS | [132] | |
| Mg | Rice | Efficient sugar partitioning, better root proliferation | [133] |
| Wheat, | Restricted ROS production, reduced peroxidative damage in leaf chloroplasts, improved antioxidative defence enzymes | [134] | |
| Maize | |||
| Broad bean | Increased activity of plasma membrane ion transporters, improved light-induced responses of leaf mesophyll | [135] | |
| Mungbean | Greater synthesis of photosynthetic pigments, higher proline accumulation | [136] | |
| S | Rice | Alleviation of As toxicity through improved amino acids, proteome and thiol metabolisms | [137] |
| Alleviation of Cd toxicity by means of increasing Fe plaque formation, Cd chelation and vacuolar sequestration | [138] | ||
| Barley | Reduction of salt accumulation, regulation of NO signalling and ion homeostasis | [139] | |
| Alleviation of Al toxicity through stimulation of ATPase activity and reducing oxidative stress | [140] | ||
| Oilseed rape | Improved photosynthesis, nutrient uptake | [141] | |
| Mustard | Enhancing leaf ascorbate and glutathione levels, amelioration of Cd toxicity, | [142] | |
| Improved photosynthesis, salt tolerance through greater glutathione production | [143] |
| Micronutrients | Tested Crop | Mechanism to Mitigate Abiotic Stress | References |
|---|---|---|---|
| B | Chickpea | Increased antioxidative enzymes, such as SOD, CAT and APX | [190] |
| Cowpea | Increase in SOD activity, photosynthesis, leaf chlorophyll content | [191] | |
| Sunflower | Increase in SOD activity, increases photosynthetic activity | [192] | |
| Tomato | Increase in SOD activity | [192] | |
| Rice | Increase in the enzymatic activity of APX, POD and CAT. | [193] | |
| Potato | Increase in leaf proline, protein, carbohydrates and antioxidant enzymes such as polyphenol oxidase and peroxidase in tubers | [194] | |
| Zn | Maize | Higher plant biomass, stomatal conductance and quantum yield of photosystem II. Improved grain yield, RWC and chlorophyll content under drought stress. | [195,196] |
| Wheat | Higher chlorophyll content and activities of SOD, POD and CAT at grain filling stage, quantum yield of PS-II, chlorophyll content, stomatal conductance, | [197,198] | |
| Chickpea | CO2 assimilation rate, proline content and activities of SOD, APX. Increases chlorophyll and carotenoid contents, seedling vigor and seed yield. Reduces MDA contents | [199,200] | |
| Common bean | Higher shoot biomass, chlorophyll and carotenoid contents, leaf NPK content Reduces MDA contents | [201] | |
| Tomato | Improved stomatal aperture and chlorophyll content | [202] | |
| Sunflower | Higher chlorophyll, proline contents and SOD activities | [203] | |
| Fe | Rice | Higher water content, higher activity of hydrolytic enzymes amylase and protease, increased activity of SOD, CAT and glutathione peroxidase, increased cell membrane integrity, cell viability, chlorophyll and iron content, increased activity of NADPH dehydrogenase | [204] |
| Wheat | Increased root length, biomass growth and chlorophyll content, increased activity of peroxidases and SOD, decreased level of MDA | [205] | |
| Sunflower | Increased activity of antioxidant enzymes, superoxide dismutase, peroxidase, catalase and ascorbate peroxidase | [206] | |
| Soybean | Enhanced net photosynthetic rate, stomatal conductance, intercellular CO2 concentration, transpiration rate, increased shoot weight | [207] | |
| Cu | Tomato | Increased antioxidant activity, phenols, vitamin C, glutathione, and improved Na+/K+ ratio | [208] |
| Maize | Increased photosynthesis, water relation, osmotic adjustment, decreased membrane damage and lipid peroxidation, increase in RWC | [209] |
| Beneficial Nutrients | Tested Crop | Mechanisms Related to Stress Alleviation | References |
|---|---|---|---|
| Co | Wheat | Increased chlorophyll content, chlorophyll stability index and proline content | [254] |
| Maize | Enhanced physiological efficiency | [255] | |
| Black gram | Improved chlorophyll and carotenoid contents, proline and nitrate reductase contents, better cell membrane stability | [116] | |
| Soybean | Enhanced antioxidant activities | [256] | |
| Se | Maize | Accelerated net photosynthetic rate, greater integrity in chloroplast ultrastructure | [257] |
| Rapeseed | Rejuvenation of entire antioxidant system in terms of APX, MDHAR, DHAR, GR, GST, GPX, CAT, and glyoxalase I and II, higher ROS scavenging, reduced membrane peroxidation and subsequent lower production of MDA | [258] | |
| Si | Rice | Increased nutrients uptake, improved plant growth, yield, and quality | [259] |
| Wheat | Better regulation of antioxidant enzymes | [260] | |
| Maize | increased water use efficiency, reduced leaf transpiration | [261] | |
| Enhanced leaf relative water content, concentrations of photosynthetic pigments, soluble sugars, soluble proteins, free amino acids, improved K/Na ratio by reducing, Na uptake | [262] | ||
| Sorghum | Higher water uptake, greater rate of assimilates transport | [263] |
| Interacting Nutrients | Synergistic Effect | Antagonistic Effect | References |
|---|---|---|---|
| N, P, K, B, Co | Increased growth rates, accelerated enzymatic activity, proline content and cell membrane stability | - | [116] |
| N, P, K, Mo | Improved growth, physiological efficiency, nutrients uptake and yield ameliorate heat and moisture stress | - | [332] |
| K and Ca | Improved physiological, biochemical and molecular mechanisms ameliorating drought, salinity and cold stress | - | [333] |
| Zn, B and Si | Increased plant height, shoot dry weight, number of stems per plant, leaf relative water content, leaf photosynthetic rate, leaf stomatal conductance, chlorophyll content, and tuber yield | [194] | |
| B, Fe, Zn | Improved chlorophyll biosynthesis, photosynthetic rate, gaseous exchange regulation and osmoregulation to mitigate the abiotic stress in late sown lentil.Scavenge ROS, enhance antioxidant enzyme activity in chloroplast, maintain membrane integrity and decrease lipid peroxidation | - | [334] |
| Fe and Mn | Improved photosynthetic activity in plants ameliorating heat stress | - | [335] |
| Co with N/P/K/S/Zn/B/Mo/Ni/Sn | Greater uptake and utilisation of reserved and applied Co alleviating salinity, heat and moisture | - | [336] |
| Co with Ca/Mn/Fe/Cu/Cr/Cd | - | Immobilisation of available soil Co and thereby reduction in uptake of Co preventing heavy metal stress | [271] |
| B, Se | The combined application was more effective enhancing the activity of MDHAR and GR under salt stress, combined spray enhanced the enzymatic activities (APX, MDHAR, DHAR, GR, CAT, GPX, GST, POD) under salt stress | [291] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. https://doi.org/10.3390/ijms23158519
Kumari VV, Banerjee P, Verma VC, Sukumaran S, Chandran MAS, Gopinath KA, Venkatesh G, Yadav SK, Singh VK, Awasthi NK. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. International Journal of Molecular Sciences. 2022; 23(15):8519. https://doi.org/10.3390/ijms23158519
Chicago/Turabian StyleKumari, Venugopalan Visha, Purabi Banerjee, Vivek Chandra Verma, Suvana Sukumaran, Malamal Alickal Sarath Chandran, Kodigal A. Gopinath, Govindarajan Venkatesh, Sushil Kumar Yadav, Vinod Kumar Singh, and Neeraj Kumar Awasthi. 2022. "Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops" International Journal of Molecular Sciences 23, no. 15: 8519. https://doi.org/10.3390/ijms23158519
APA StyleKumari, V. V., Banerjee, P., Verma, V. C., Sukumaran, S., Chandran, M. A. S., Gopinath, K. A., Venkatesh, G., Yadav, S. K., Singh, V. K., & Awasthi, N. K. (2022). Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. International Journal of Molecular Sciences, 23(15), 8519. https://doi.org/10.3390/ijms23158519