
Dimethyl Fumarate Induces Apoptosis via Inhibition of NF-κB and Enhances the Effect of Paclitaxel and Adriamycin in Human TNBC Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phosphorylated NF-κB p65 Protein and NFKBIA Gene Expression Is Associated with Prognosis in Patients with TNBC
2.2. DMF Induced Apoptosis in MDA-MB-231 and BT549 Cells
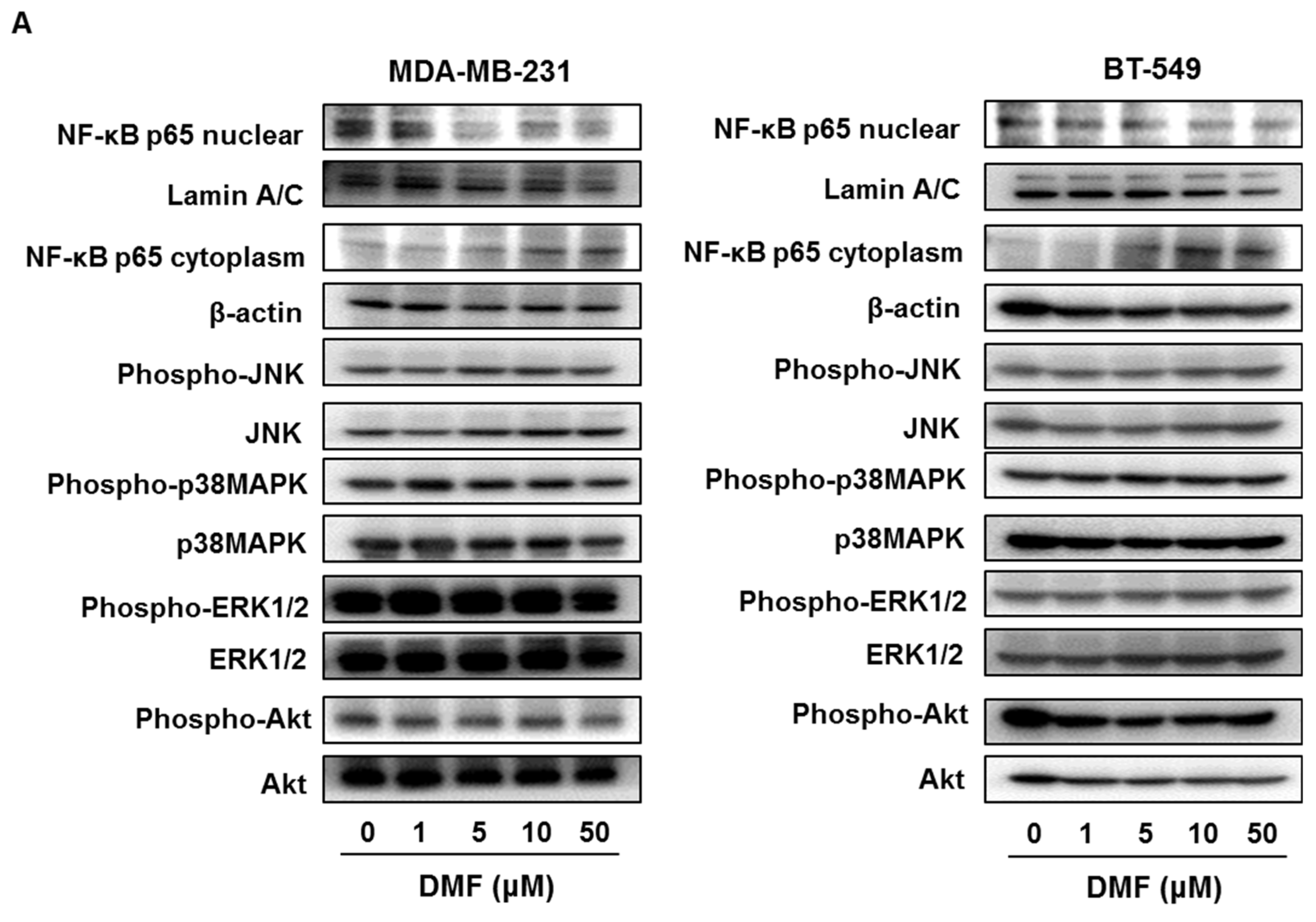
2.3. DMF Suppressed NF-κB p65 Nuclear Translocation and XIAP, Survivin, Bcl-xL, and Bcl-2 Expression in MDA-MB-231 and BT-549 Cells
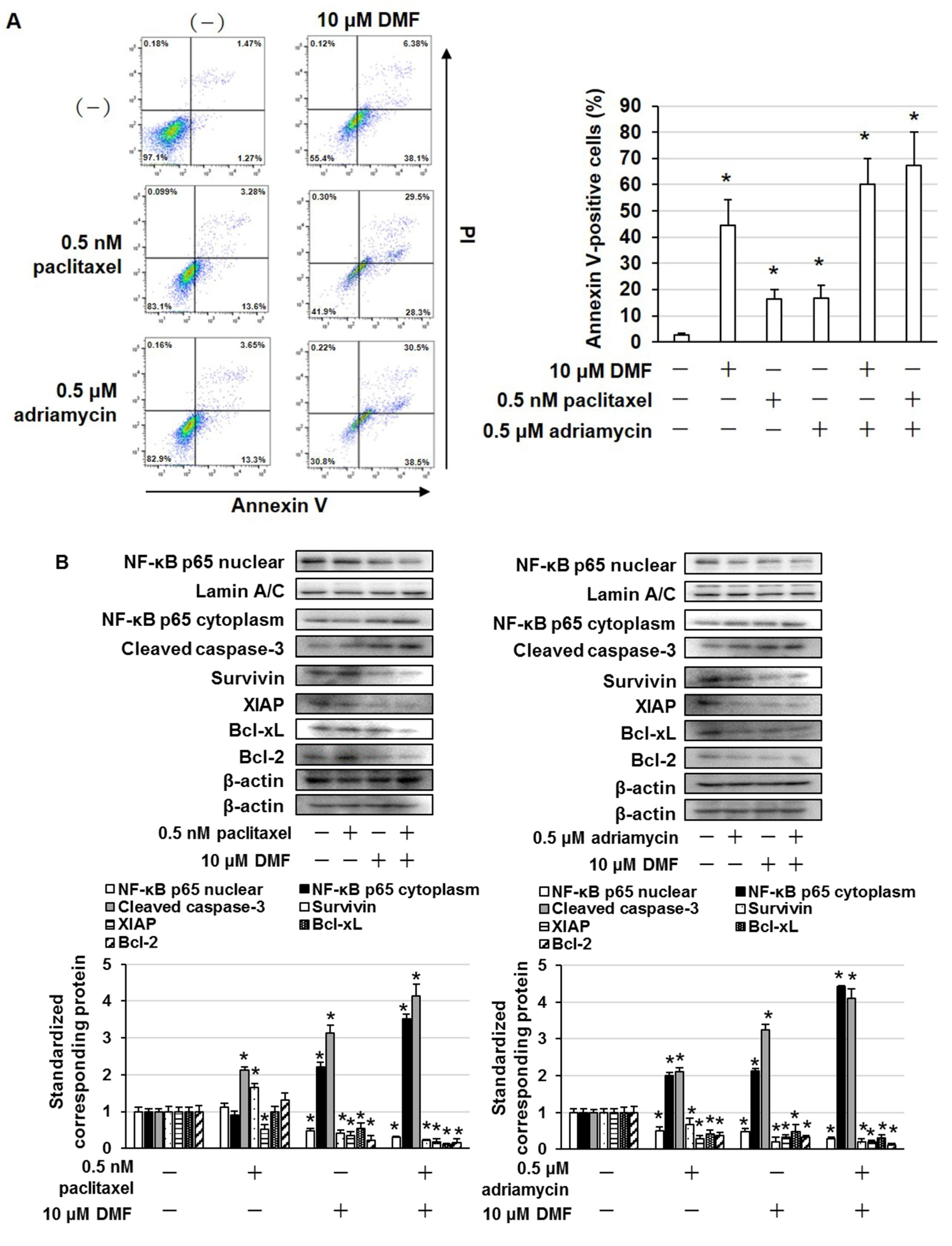
2.4. DMF Enhanced the Cytotoxic Effects of Paclitaxel and Adriamycin in MDA-MB-231 and BT-549 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. Cell Survival and Apoptosis Detection Assay
4.3. Immunoblotting
4.4. Gene Expression Omnibus Dataset
4.5. Kaplan–Meier Plotter Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, P.; Scatena, C.; Ghilli, M.; Bargagna, I.; Lorenzini, G.; Nicolini, A. Molecular Mechanisms, Biomarkers and Emerging Therapies for Chemotherapy Resistant TNBC. Int. J. Mol. Sci. 2022, 23, 1665. [Google Scholar] [CrossRef] [PubMed]
- Mireștean, C.C.; Volovăț, C.; Iancu, R.I.; Iancu, D.P.T. Radiomics in Triple Negative Breast Cancer: New Horizons in an Ag-gressive Subtype of the Disease. J. Clin. Med. 2022, 11, 616. [Google Scholar]
- Kanzaki, H.; Chatterjee, A.; Ariani, H.H.N.; Zhang, X.; Chung, S.; Deng, N.; Ramanujan, V.K.; Cui, X.; Greene, M.I.; Murali, R. Disabling the Nuclear Translocalization of RelA/NF-κB by a Small Molecule Inhibits Triple-Negative Breast Cancer Growth. Breast Cancer 2021, 13, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Ehmsen, S.; Ditzel, H.J. Signaling pathways essential for triple-negative breast cancer stem-like cells. Stem Cells 2020, 39, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Jung, H.H.; Ahn, S.; Bae, S.; Lee, S.K.; Kim, S.W.; Lee, J.E.; Nam, S.J.; Ahn, J.S.; Im, Y.H.; et al. The relationship between nuclear factor (NF)-κB family gene expression and prognosis in triple-negative breast cancer (TNBC) patients receiving adjuvant doxorubicin treatment. Sci. Rep. 2016, 6, 31804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Ma, C.; Zhang, Z.; Zhang, H.; Hu, H. NF-κB signaling in inflammation and cancer. MedComm 2021, 2, 618–653. [Google Scholar]
- Lategan, T.W.; Wang, L.; Sprague, T.N.; Rousseau, F.S. Pharmacokinetics and Bioavailability of Monomethyl Fumarate Fol-lowing a Single Oral Dose of Bafiertam™ (Monomethyl Fumarate) or Tecfidera® (Dimethyl Fumarate). CNS Drugs 2021, 35, 567–574. [Google Scholar] [CrossRef]
- Takeda, T.; Tsubaki, M.; Asano, R.; Itoh, T.; Imano, M.; Satou, T.; Nishida, S. Dimethyl fumarate suppresses metastasis and growth of melanoma cells by inhibiting the nuclear translocation of NF-κB. J. Dermatol. Sci. 2020, 99, 168–176. [Google Scholar] [CrossRef]
- Kaluzki, I.; Hailemariam-Jahn, T.; Doll, M.; Kaufmann, R.; Balermpas, P.; Zöller, N.; Kippenberger, S.; Meissner, M. Dime-thylfumarate Inhibits Colorectal Carcinoma Cell Proliferation: Evidence for Cell Cycle Arrest, Apoptosis and Autophagy. Cells 2019, 8, 1329. [Google Scholar] [CrossRef] [Green Version]
- Nicolay, J.P.; Müller-Decker, K.; Schroeder, A.; Brechmann, M.; Möbs, M.; Géraud, C.; Assaf, C.; Goerdt, S.; Krammer, P.H.; Gülow, K. Dimethyl fumarate restores apoptosis sensitivity and inhibits tumor growth and metastasis in CTCL by targeting NF-κB. Blood 2016, 128, 805–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubaki, M.; Ogawa, N.; Takeda, T.; Sakamoto, K.; Shimaoka, H.; Fujita, A.; Itoh, T.; Imano, M.; Satou, T.; Nishida, S. Dimethyl fumarate induces apoptosis of hematopoietic tumor cells via inhibition of NF-κB nuclear translocation and down-regulation of Bcl-xL and XIAP. Biomed. Pharmacother. 2014, 68, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, B.; Yu, J. Mitochondrial signaling in cell death via the Bcl-2 family. Cancer Biol. Ther. 2010, 9, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittendorf, E.A.; Zhang, H.; Barrios, C.H.; Saji, S.; Jung, K.H.; Hegg, R.; Koehler, A.; Sohn, J.; Iwata, H.; Telli, M.L.; et al. Neo-adjuvant atezolizumab in combination with sequential nab-paclitaxel and anthracycline-based chemotherapy versus placebo and chemotherapy in patients with early-stage triple-negative breast cancer (IMpassion031): A randomised, double-blind, phase 3 trial. Lancet 2020, 396, 1090–1100. [Google Scholar]
- Bredel, M.; Kim, H.; Thudi, N.K.; Scholtens, D.M.; Bonner, J.A.; Sikic, B.I. NFKBIA deletion in triple-negative breast cancer. J. Clin. Oncol. 2013, 31, 1012. [Google Scholar] [CrossRef]
- Momeny, M.; Zakidizaji, M.; Ghasemi, R.; Dehpour, A.R.; Rahimi-Balaei, M.; Abdolazimi, Y.; Ghavamzadeh, A.; Ali-Moghaddam, K.; Ghaffari, S.H. Arsenic trioxide induces apoptosis in NB-4, an acute promyelocytic leukemia cell line, through up-regulation of p73 via suppression of nuclear factor kappa B-mediated inhibition of p73 transcription and prevention of NF-kappaB-mediated induction of XIAP, cIAP2, BCL-XL and survivin. Med. Oncol. 2010, 27, 833–842. [Google Scholar] [PubMed]
- Su, J.; Zhao, P.; Kong, L.; Li, X.; Yan, J.; Zeng, Y.; Li, Y. Trichothecin induces cell death in NF-κB constitutively activated human cancer cells via inhibition of IKKβ phosphorylation. PLoS ONE 2013, 8, e71333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batool, R.; Aziz, E.; Tan, B.K.; Mahmood, T. Rumex dentatus Inhibits Cell Proliferation, Arrests Cell Cycle, and Induces Apoptosis in MDA-MB-231 Cells through Suppression of the NF-κB Pathway. Front. Pharmacol. 2017, 8, 731. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, Y.; Su, R.; Jia, Y.; Lai, X.; Su, H.; Fan, Y.; Wang, Y.; Xing, W.; Qin, J. Dimethyl Fumarate Combined with Vemurafenib Enhances Anti-Melanoma Efficacy via Inhibiting the Hippo/YAP, NRF2-ARE, and AKT/mTOR/ERK Pathways in A375 Melanoma Cells. Front. Oncol. 2022, 12, 794216. [Google Scholar] [CrossRef]
- Shafer, D.; Tombes, M.B.; Shrader, E.; Ryan, A.; Bandyopadhyay, D.; Dent, P.; Malkin, M. Phase I trial of dimethyl fumarate, temozolomide, and radiation therapy in glioblastoma. Neurooncol. Adv. 2020, 2, vdz052. [Google Scholar] [CrossRef]
- Nicolay, J.P.; Albrecht, J.D.; Assaf, C.; Dippel, E.; Stadler, R.; Wehkamp, U.; Wobser, M.; Guelow, K.; Goerdt, S.; Krammer, P.H. Dimethyl fumarate (DMF) therapy in CTCL: Results from a clinical phase II study. Eur. J. Cancer 2021, 156, S21–S22. [Google Scholar] [CrossRef]
- Kastrati, I.; Siklos, M.I.; Calderon-Gierszal, E.L.; El-Shennawy, L.; Georgieva, G.; Thayer, E.N.; Thatcher, G.R.; Frasor, J. Di-methyl Fumarate Inhibits the Nuclear Factor κB Pathway in Breast Cancer Cells by Covalent Modification of p65 Protein. J. Biol. Chem. 2016, 291, 3639–3647. [Google Scholar] [CrossRef] [Green Version]
- Tsubaki, M.; Takeda, T.; Noguchi, M.; Jinushi, M.; Seki, S.; Morii, Y.; Shimomura, K.; Imano, M.; Satou, T.; Nishida, S. Over-activation of Akt Contributes to MEK Inhibitor Primary and Acquired Resistance in Colorectal Cancer Cells. Cancers 2019, 11, 1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubaki, M.; Takeda, T.; Tomonari, Y.; Koumoto, Y.I.; Imano, M.; Satou, T.; Nishida, S. Overexpression of HIF-1α contributes to melphalan resistance in multiple myeloma cells by activation of ERK1/2, Akt, and NF-κB. Lab. Investig. 2019, 99, 72–84. [Google Scholar] [CrossRef]
- Tsubaki, M.; Takeda, T.; Tomonari, Y.; Mashimo, K.; Koumoto, Y.; Hoshida, S.; Itoh, T.; Imano, M.; Satou, T.; Sakaguchi, K.; et al. The MIP-1α autocrine loop contributes to decreased sensitivity to anticancer drugs. J. Cell. Physiol. 2017, 233, 4258–4271. [Google Scholar] [CrossRef] [PubMed]
- Morii, Y.; Tsubaki, M.; Takeda, T.; Otubo, R.; Seki, S.; Yamatomo, Y.; Imano, M.; Satou, T.; Shimomura, K.; Nishida, S. Perifosine enhances the potential antitumor effect of 5-fluorourasil and oxaliplatin in colon cancer cells harboring the PIK3CA mutation. Eur. J. Pharmacol. 2021, 898, 173957. [Google Scholar] [CrossRef]
- Gyorffy, B.; Lánczky, A.; Szállási, Z. Implementing an online tool for genome-wide validation of survival-associated bi-omarkers in ovarian-cancer using microarray data from 1287 patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ősz, Á.; Lánczky, A.; Győrffy, B. Survival analysis in breast cancer using proteomic data from four independent datasets. Sci. Rep. 2021, 11, 16787. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsurushima, K.; Tsubaki, M.; Takeda, T.; Matsuda, T.; Kimura, A.; Takefuji, H.; Okada, A.; Sakamoto, C.; Ishizaka, T.; Nishida, S. Dimethyl Fumarate Induces Apoptosis via Inhibition of NF-κB and Enhances the Effect of Paclitaxel and Adriamycin in Human TNBC Cells. Int. J. Mol. Sci. 2022, 23, 8681. https://doi.org/10.3390/ijms23158681
Tsurushima K, Tsubaki M, Takeda T, Matsuda T, Kimura A, Takefuji H, Okada A, Sakamoto C, Ishizaka T, Nishida S. Dimethyl Fumarate Induces Apoptosis via Inhibition of NF-κB and Enhances the Effect of Paclitaxel and Adriamycin in Human TNBC Cells. International Journal of Molecular Sciences. 2022; 23(15):8681. https://doi.org/10.3390/ijms23158681
Chicago/Turabian StyleTsurushima, Katsumasa, Masanobu Tsubaki, Tomoya Takeda, Takuya Matsuda, Akihiro Kimura, Honoka Takefuji, Akane Okada, Chiaki Sakamoto, Toshihiko Ishizaka, and Shozo Nishida. 2022. "Dimethyl Fumarate Induces Apoptosis via Inhibition of NF-κB and Enhances the Effect of Paclitaxel and Adriamycin in Human TNBC Cells" International Journal of Molecular Sciences 23, no. 15: 8681. https://doi.org/10.3390/ijms23158681
APA StyleTsurushima, K., Tsubaki, M., Takeda, T., Matsuda, T., Kimura, A., Takefuji, H., Okada, A., Sakamoto, C., Ishizaka, T., & Nishida, S. (2022). Dimethyl Fumarate Induces Apoptosis via Inhibition of NF-κB and Enhances the Effect of Paclitaxel and Adriamycin in Human TNBC Cells. International Journal of Molecular Sciences, 23(15), 8681. https://doi.org/10.3390/ijms23158681