
Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. The Experimental Data—On the Effects of Mutations in Plant Promoters on Gene Expression—That We Could Find in the PubMed Database in Order to Investigate Them in This Work
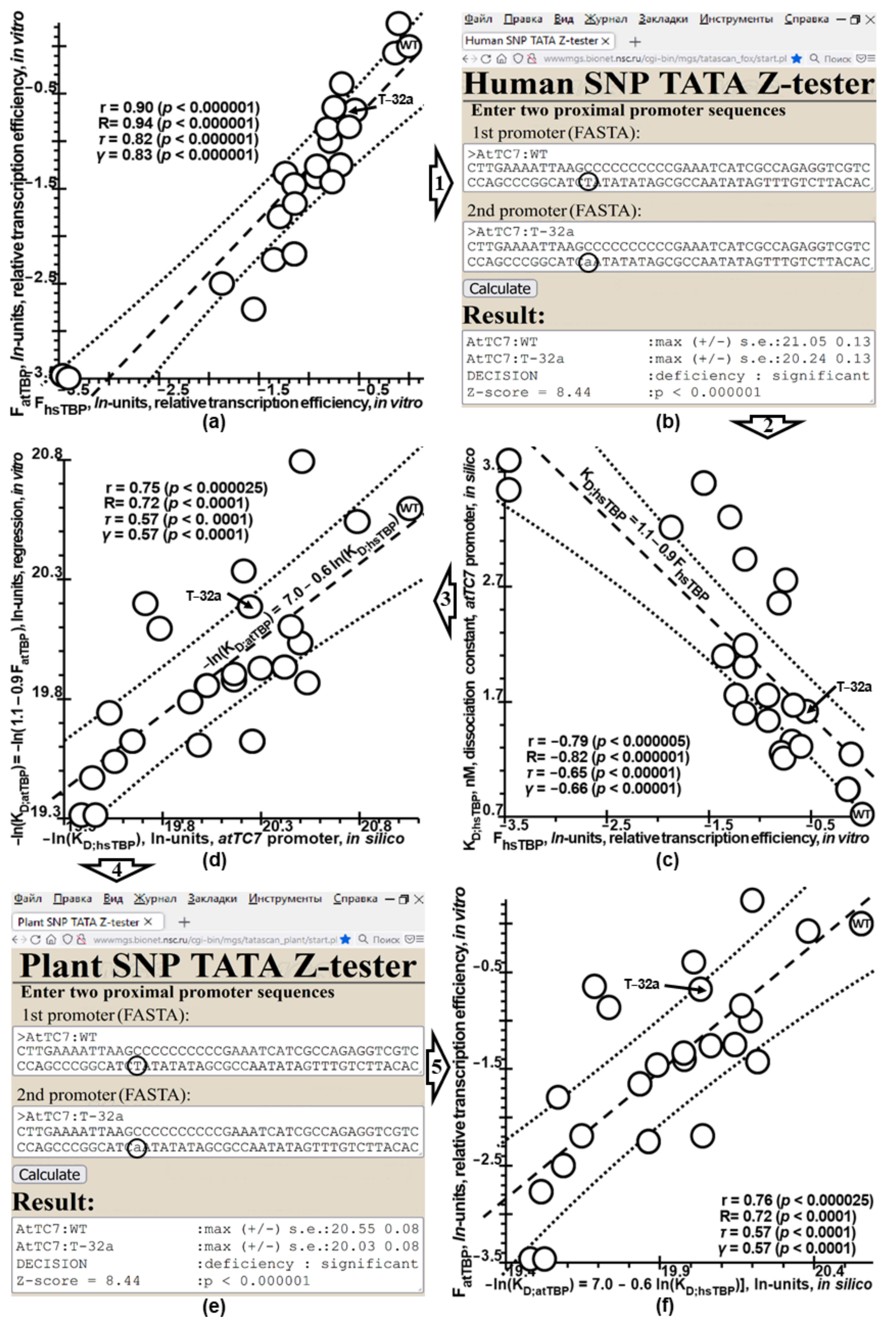
2.2. The Ill-Posed Inverse Problem under Study and Its Solution via Tikhonov’s Regularization
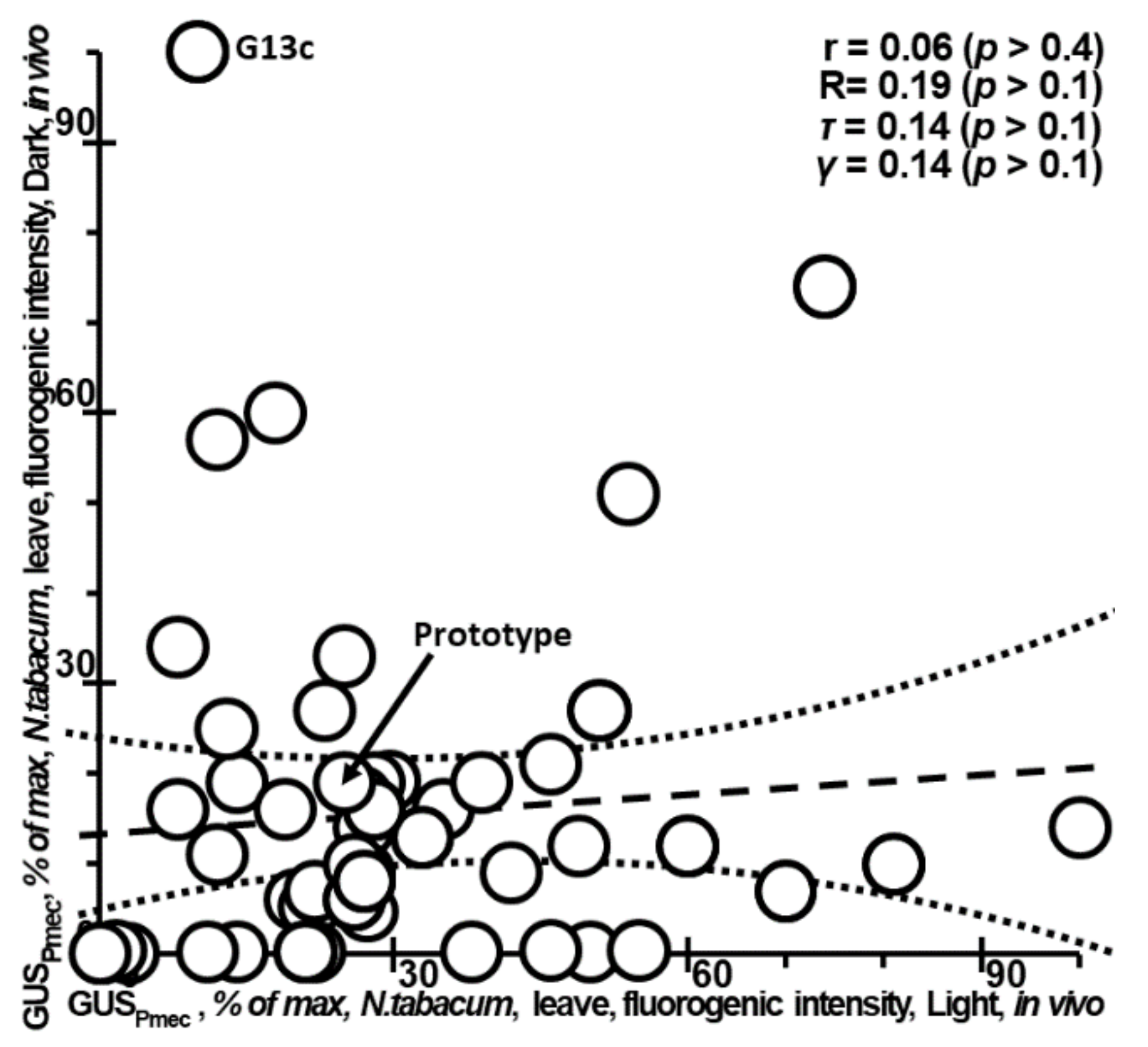
2.3. Determining Application Limits of Plant_SNP_TATA_Z-Tester by Means of Experimental Data on Tobacco Development in the Dark or under Light, Indicating That the Inverse Problem under Study Is Ill-Posed
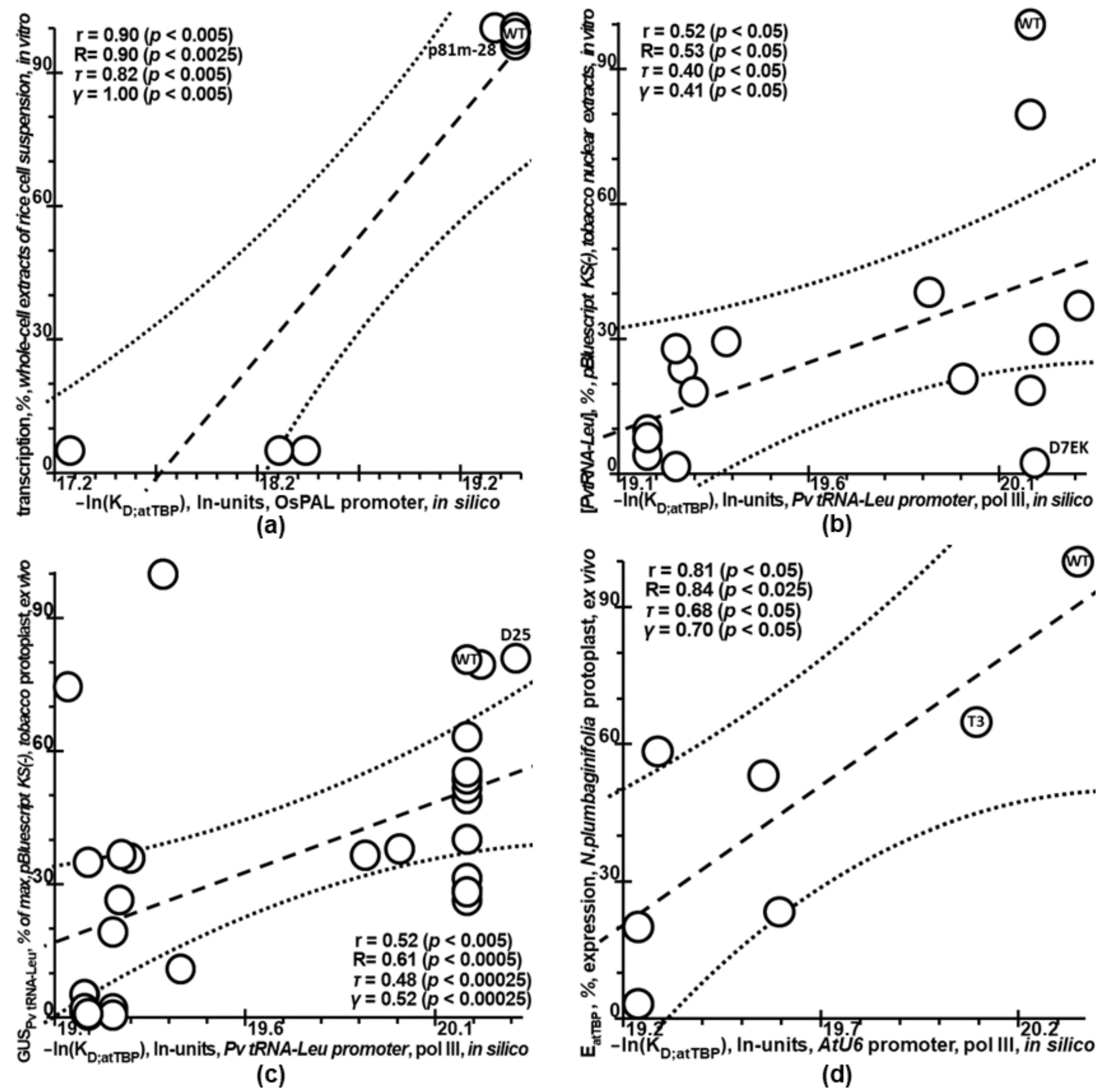
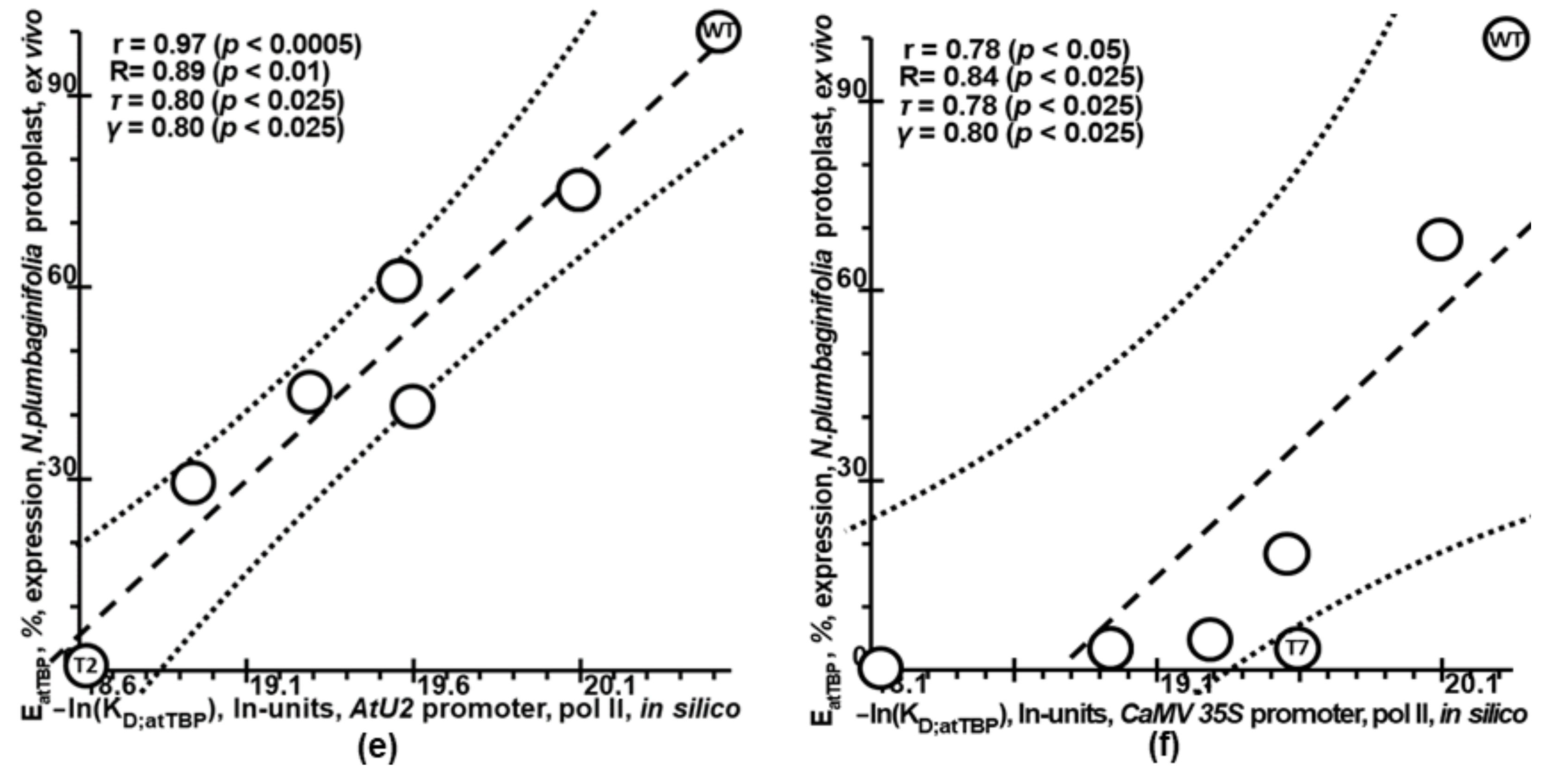
2.4. Verification of Plant_SNP_TATA_Z-Tester Using Independent Experimental Data on Mutations within Natural Proximal Promoters of Plant Genes
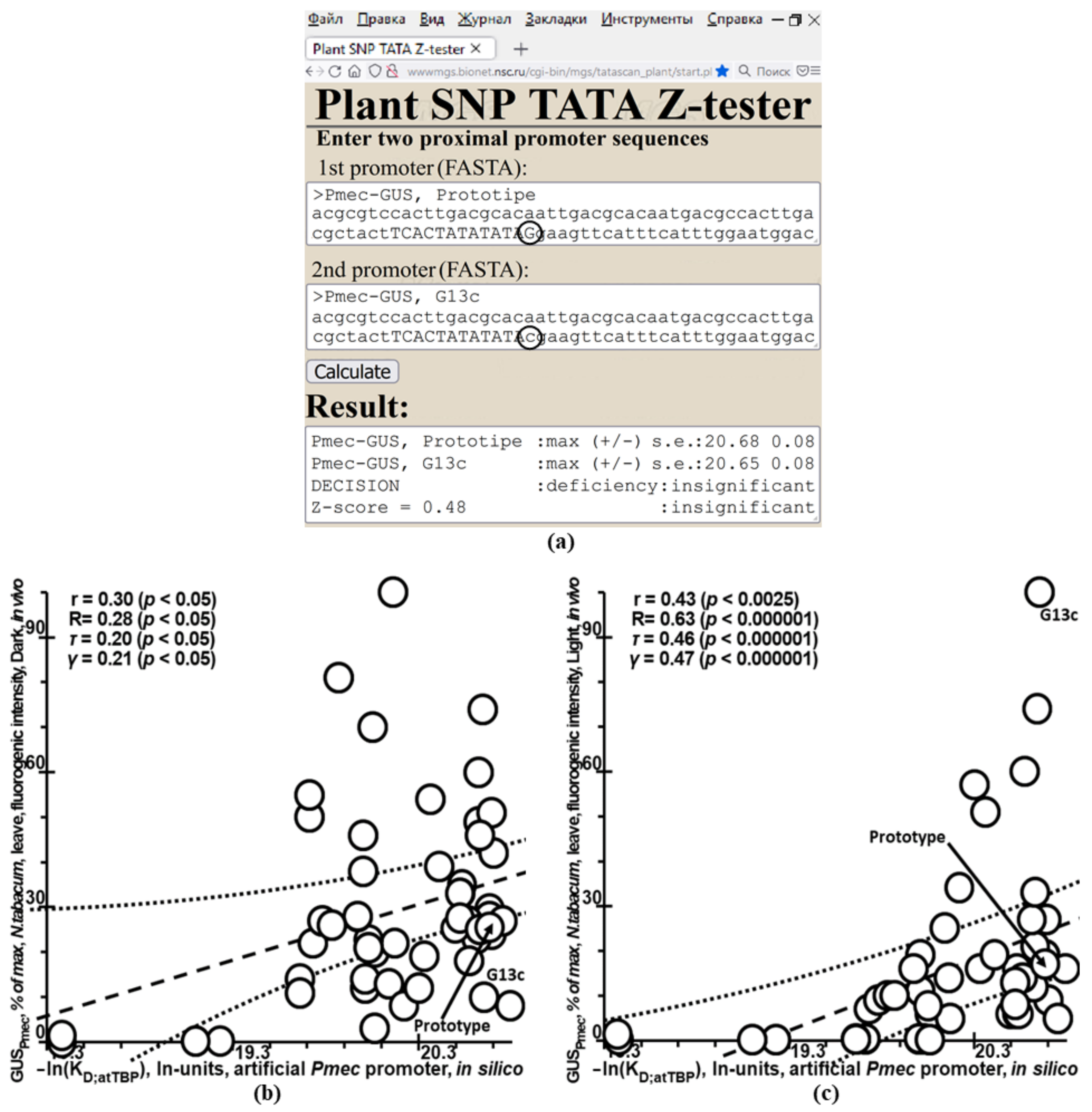
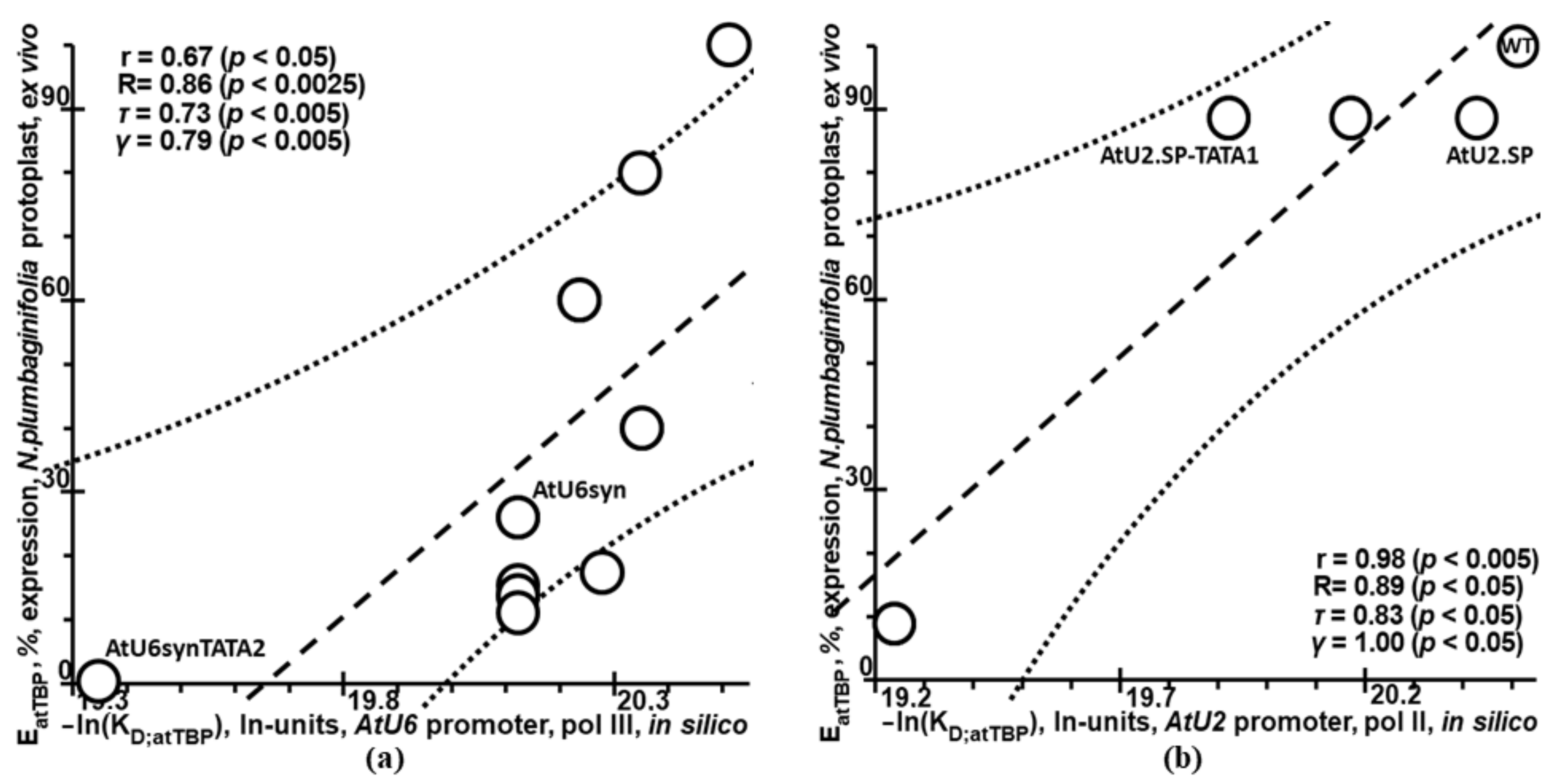
2.5. Validation of Plant_SNP_TATA_Z-Tester by Means of Experimental Data on Mutations in the Synthetic Proximal Promoters Designed on the Basis of Natural Ones
3. Discussion
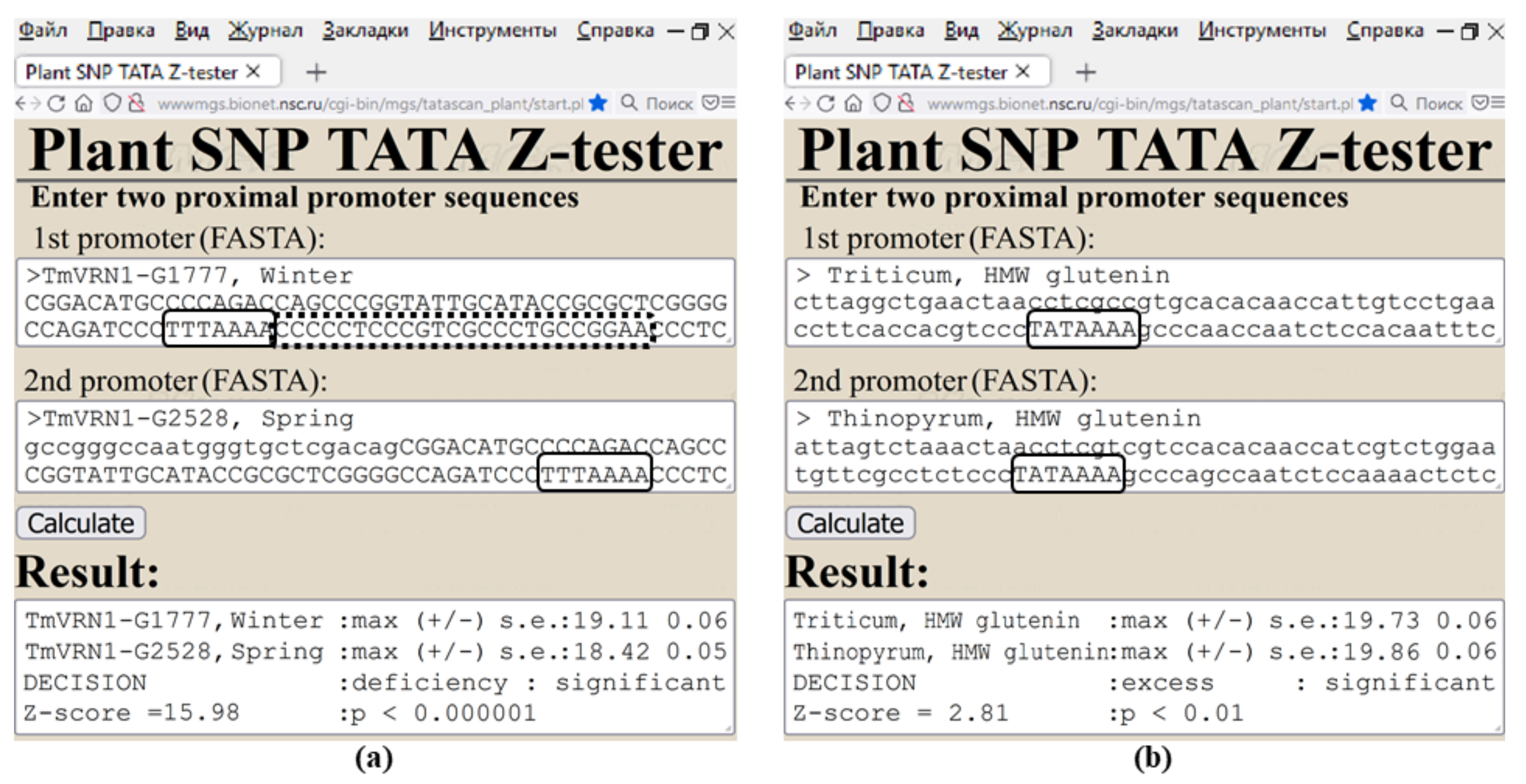
4. Materials and Methods
4.1. Experimental Data under Study
4.2. In Silico Analysis of DNA Sequences
4.3. The Knowledge Base (on Effects of Mutations in Plant Promoters on Gene Expression) Created in this Work
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CaMV | Cauliflower mosaic virus |
| HMW | High-molecular-weight |
| ln units | Natural logarithm units |
| QTL | Quantitative trait loci |
| PAM | Protospacer-adjacent motif |
| SNP | Single-nucleotide polymorphism |
| TAIR | The Arabidopsis Information Resource |
| TBP | TATA-binding protein |
| TSS | Transcription start site |
References
- Buzdin, A.V.; Patrushev, M.V.; Sverdlov, E.D. Will plant genome editing play a decisive role in “quantum-leap” improvements in crop yield to feed an increasing global human population? Plants 2021, 10, 1667. [Google Scholar] [CrossRef]
- Kim, K.D.; Kang, Y.; Kim, C. Application of genomic Big Data in plant breeding: Past, present, and future. Plants 2020, 9, 1454. [Google Scholar] [CrossRef]
- Jores, T.; Tonnies, J.; Wrightsman, T.; Buckler, E.S.; Cuperus, J.T.; Fields, S.; Queitsch, C. Synthetic promoter designs enabled by a comprehensive analysis of plant core promoters. Nat. Plants 2021, 7, 842–855. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Yadav, O.P.; Kaliamoorthy, S.; Gupta, S.K.; Serba, D.D.; Choudhary, S.; Govindaraj, M.; Kholova, J.; Murugesan, T.; Satyavathi, C.T.; et al. Breeding drought-tolerant pearl millet using conventional and genomic approaches: Achievements and prospects. Front. Plant Sci. 2022, 13, 781524. [Google Scholar] [CrossRef]
- Bernardo, R. Bandwagons I, too, have known. Theor. Appl. Genet. 2016, 129, 2323–2332. [Google Scholar] [CrossRef]
- Alix, K.; Gerard, P.R.; Schwarzacher, T.; Heslop-Harrison, J.S.P. Polyploidy and interspecific hybridization: Partners for adaptation, speciation and evolution in plants. Ann. Bot. 2017, 120, 183–194. [Google Scholar] [CrossRef]
- Manimekalai, R.; Suresh, G.; Govinda Kurup, H.; Athiappan, S.; Kandalam, M. Role of NGS and SNP genotyping methods in sugarcane improvement programs. Crit. Rev. Biotechnol. 2020, 40, 865–880. [Google Scholar] [CrossRef]
- Cui, Z.; Tian, R.; Huang, Z.; Jin, Z.; Li, L.; Liu, J.; Huang, Z.; Xie, H.; Liu, D.; Mo, H.; et al. FrCas9 is a CRISPR/Cas9 system with high editing efficiency and fidelity. Nat. Commun. 2022, 13, 1425. [Google Scholar] [CrossRef]
- Yang, M.Q.; Laflamme, K.; Gotea, V.; Joiner, C.H.; Seidel, N.E.; Wong, C.; Petrykowska, H.M.; Lichtenberg, J.; Lee, S.; Welch, L.; et al. Genome-wide detection of a TFIID localization element from an initial human disease mutation. Nucleic Acids Res. 2011, 39, 2175–2187. [Google Scholar] [CrossRef] [Green Version]
- Rhee, H.S.; Pugh, B.F. Genome-wide structure and organization of eukaryotic pre-initiation complexes. Nature 2012, 483, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Choukrallah, M.A.; Kobi, D.; Martianov, I.; Pijnappel, W.W.; Mischerikow, N.; Ye, T.; Heck, A.J.; Timmers, H.T.; Davidson, I. Interconversion between active and inactive TATA-binding protein transcription complexes in the mouse genome. Nucleic Acids Res. 2012, 40, 1446–1459. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y.; Ichida, H.; Matsui, M.; Obokata, J.; Sakurai, T.; Satou, M.; Seki, M.; Shinozaki, K.; Abe, T. Identification of plant promoter constituents by analysis of local distribution of short sequences. BMC Genom. 2007, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Muller, F.; Lakatos, L.; Dantonel, J.; Strähle, U.; Tora, L. TBP is not universally required for zygotic RNA polymerase II transcription in zebrafish. Curr. Biol. 2001, 11, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Martianov, I.; Viville, S.; Davidson, I. RNA polymerase II transcription in murine cells lacking the TATA binding protein. Science 2002, 298, 1036–1039. [Google Scholar] [CrossRef]
- Mogno, I.; Vallania, F.; Mitra, R.D.; Cohen, B.A. TATA is a modular component of synthetic promoters. Genome Res. 2010, 20, 1391–1397. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, M.; Mironova, V.; Gunbin, K.; Savinkova, L. Hogness Box. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, CA, USA, 2013; Volume 3, pp. 491–494. [Google Scholar] [CrossRef]
- Savinkova, L.; Ponomarenko, M.; Ponomarenko, P.; Drachkova, I.; Lysova, M.; Arshinova, T.; Kolchanov, N. TATA box polymorphisms in human gene promoters and associated hereditary pathologies. Biochemistry 2009, 74, 117–129. [Google Scholar] [CrossRef]
- Coleman, R.A.; Pugh, B.F. Evidence for functional binding and stable sliding of the TATA binding protein on nonspecific DNA. J. Biol. Chem. 1995, 270, 13850–13859. [Google Scholar] [CrossRef] [Green Version]
- Porto, M.S.; Pinheiro, M.P.; Batista, V.G.; dos Santos, R.C.; Filho, P.d.A.; de Lima, L.M. Plant promoters: An approach of structure and function. Mol. Biotechnol. 2014, 56, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Joshi, C.P. An inspection of the domain between putative TATA box and translation start site in 79 plant genes. Nucleic Acids Res. 1987, 15, 6643–6653. [Google Scholar] [CrossRef] [Green Version]
- Ohno, S.; Yomo, T. Various regulatory sequences are deprived of their uniqueness by the universal rule of TA/CG deficiency and TG/CT excess. Proc. Natl. Acad. Sci. USA 1990, 87, 1218–1222. [Google Scholar] [CrossRef] [Green Version]
- Korkuc, P.; Schippers, J.H.; Walther, D. Characterization and identification of cis-regulatory elements in Arabidopsis based on single-nucleotide polymorphism information. Plant Physiol. 2014, 164, 181–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplancke, B.; Alpern, D.; Gardeux, V. The genetics of transcription factor DNA binding variation. Cell 2016, 166, 538–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolachevskaya, O.O.; Myakushina, Y.A.; Getman, I.A.; Lomin, S.N.; Deyneko, I.V.; Deigraf, S.V.; Romanov, G.A. Hormonal regulation and crosstalk of auxin/cytokinin signaling pathways in potatoes in vitro and in relation to vegetation or tuberization stages. Int. J. Mol. Sci. 2021, 22, 8207. [Google Scholar] [CrossRef] [PubMed]
- Ageev, A.; Lee, C.-R.; Ting, C.-T.; Schafleitner, R.; Bishop-von Wettberg, E.; Nuzhdin, S.V.; Samsonova, M.; Kozlov, K. Modeling of flowering time in Vigna radiata with approximate bayesian computation. Agronomy 2021, 11, 2317. [Google Scholar] [CrossRef]
- Rakhmetullina, A.; Pyrkova, A.; Aisina, D.; Ivashchenko, A. In silico prediction of human genes as potential targets for rice miRNAs. Comput. Biol. Chem. 2020, 87, 107305. [Google Scholar] [CrossRef]
- Ponomarenko, M.; Rasskazov, D.; Arkova, O.; Ponomarenko, P.; Suslov, V.; Savinkova, L.; Kolchanov, N. How to use SNP_TATA_Comparator to find a significant change in gene expression caused by the regulatory SNP of this gene’s promoter via a change in affinity of the TATA-binding protein for this promoter. Biomed. Res. Int. 2015, 2015, 359835. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, M.; Kleshchev, M.; Ponomarenko, P.; Chadaeva, I.; Sharypova, E.; Rasskazov, D.; Kolmykov, S.; Drachkova, I.; Vasiliev, G.; Gutorova, N.; et al. Disruptive natural selection by male reproductive potential prevents underexpression of protein-coding genes on the human Y chromosome as a self-domestication syndrome. BMC Genet. 2020, 21, 89. [Google Scholar] [CrossRef]
- Ponomarenko, P.; Savinkova, L.; Drachkova, I.; Lysova, M.; Arshinova, T.; Ponomarenko, M.; Kolchanov, N. A step-by-step model of TBP/TATA box binding allows predicting human hereditary diseases by single nucleotide polymorphism. Dokl. Biochem. Biophys. 2008, 419, 88–92. [Google Scholar] [CrossRef]
- Berg, O.G.; von Hippel, P.H. Selection of DNA binding sites by regulatory proteins. Statistical-mechanical theory and application to operators and promoters. J. Mol. Biol. 1987, 193, 723–750. [Google Scholar] [CrossRef]
- Bucher, P. Weight matrix descriptions of four eukaryotic RNA polymerase II promoter elements derived from 502 unrelated promoter sequences. J. Mol. Biol. 1990, 212, 563–578. [Google Scholar] [CrossRef]
- Flatters, D.; Lavery, R. Sequence-dependent dynamics of TATA-Box binding sites. Biophys. J. 1998, 75, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo, R.; Whittington, J.; Parkhurst, L.; Parkhurst, L. The TBP core domain in solution variably bends TATA sequences via a three-step binding mechanism. Biochemistry 2009, 48, 1801–1809. [Google Scholar] [CrossRef]
- Vasiliev, G.; Chadaeva, I.; Rasskazov, D.; Ponomarenko, P.; Sharypova, E.; Drachkova, I.; Bogomolov, A.; Savinkova, L.; Ponomarenko, M.; Kolchanov, N.; et al. A bioinformatics model of human diseases on the basis of differentially expressed genes (of domestic versus wild animals) that are orthologs of human genes associated with reproductive-potential changes. Int. J. Mol. Sci. 2021, 22, 2346. [Google Scholar] [CrossRef]
- Arkova, O.; Ponomarenko, M.; Rasskazov, D.; Drachkova, I.; Arshinova, T.; Ponomarenko, P.; Savinkova, L.; Kolchanov, N. Obesity-related known and candidate SNP markers can significantly change affinity of TATA-binding protein for human gene promoters. BMC Genom. 2015, 16, S5. [Google Scholar] [CrossRef] [Green Version]
- Oshchepkov, D.; Chadaeva, I.; Kozhemyakina, R.; Zolotareva, K.; Khandaev, B.; Sharypova, E.; Ponomarenko, P.; Bogomolov, A.; Klimova, N.V.; Shikhevich, S.; et al. Stress reactivity, susceptibility to hypertension, and differential expression of genes in hypertensive compared to normotensive patients. Int. J. Mol. Sci. 2022, 23, 2835. [Google Scholar] [CrossRef]
- Ponomarenko, M.; Sharypova, E.; Drachkova, I.; Chadaeva, I.; Arkova, O.; Podkolodnaya, O.; Ponomarenko, P.; Kolchanov, N.; Savinkova, L. Unannotated single nucleotide polymorphisms in the TATA box of erythropoiesis genes show in vitro positive involvements in cognitive and mental disorders. BMC Med. Genet. 2020, 21, 165. [Google Scholar] [CrossRef]
- Ponomarenko, M.; Rasskazov, D.; Chadaeva, I.; Sharypova, E.; Drachkova, I.; Oshchepkov, D.; Ponomarenko, P.; Savinkova, L.; Oshchepkova, E.; Nazarenko, M.; et al. Candidate SNP markers of atherogenesis significantly shifting the affinity of TATA-binding protein for human gene promoters show stabilizing natural selection as a sum of neutral drift accelerating atherogenesis and directional natural selection slowing it. Int. J. Mol. Sci. 2020, 21, 1045. [Google Scholar] [CrossRef] [Green Version]
- Arkova, O.V.; Drachkova, I.A.; Arshinova, T.V.; Rasskazov, D.A.; Suslov, V.V.; Ponomarenko, P.M.; Ponomarenko, M.P.; Kolchanov, N.A.; Savinkova, L.K. Prediction and verification of the influence of the rs367781716 SNP on the interaction of the TATA-binding protein with the promoter of the human ABCA9 gene. Russ. J. Genet. Appl. Res. 2016, 6, 785–791. [Google Scholar] [CrossRef]
- Ponomarenko, P.; Chadaeva, I.; Rasskazov, D.A.; Sharypova, E.; Kashina, E.V.; Drachkova, I.; Zhechev, D.; Ponomarenko, M.P.; Savinkova, L.K.; Kolchanov, N. Candidate SNP markers of familial and sporadic Alzheimer’s diseases are predicted by a significant change in the affinity of TATA-binding protein for human gene promoters. Front. Aging Neurosci. 2017, 9, 231. [Google Scholar] [CrossRef]
- Sharypova, E.B.; Drachkova, I.A.; Kashina, E.V.; Rasskazov, D.A.; Ponomarenko, P.M.; Ponomarenko, M.P.; Kolchanov, N.A.; Savinkova, L.K. An experimental study of the effect of rare polymorphisms of human HBB, HBD and F9 promoter TATA boxes on the kinetics of interaction with the TATA-binding protein. Vavilov. J. Genet. Breed. 2018, 22, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Drachkova, I.; Savinkova, L.; Arshinova, T.; Ponomarenko, M.; Peltek, S.; Kolchanov, N. The mechanism by which TATA-box polymorphisms associated with human hereditary diseases influence interactions with the TATA-binding protein. Hum. Mutat. 2014, 35, 601–608. [Google Scholar] [CrossRef]
- Varzari, A.; Tudor, E.; Bodrug, N.; Corloteanu, A.; Axentii, E.; Deyneko, I.V. Age-specific association of CCL5 gene polymorphism with pulmonary tuberculosis: A case-control study. Genet. Test. Mol. Biomark. 2018, 22, 281–287. [Google Scholar] [CrossRef]
- Lu, Z. PubMed and beyond: A survey of web tools for searching biomedical literature. Database 2011, 2011, baq036. [Google Scholar] [CrossRef] [Green Version]
- Mukumoto, F.; Hirose, S.; Imaseki, H.; Yamazaki, K. DNA sequence requirement of a TATA element-binding protein from Arabidopsis for transcription in vitro. Plant Mol. Biol. 1993, 23, 95–1003. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Dabi, T.; Beeche, A.; Yamamoto, R.; Lawton, M.A.; Lamb, C. Cloning and properties of a rice gene encoding phenylalanine ammonia-lyase. Plant. Mol. Biol. 1995, 29, 535–550. [Google Scholar] [CrossRef]
- Zhu, Q.; Dabi, T.; Lamb, C. TATA box and initiator functions in the accurate transcription of a plant minimal promoter in vitro. Plant Cell. 1995, 7, 1681–1689. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, Y.; Sugita, M.; Choisne, N.; Small, I.; Sugiura, M. The TATA motif, the CAA motif and the poly(T) transcription termination motif are all important for transcription re-initiation on plant tRNA genes. Plant J. 2000, 22, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Choisne, N.; Martin-Canadell, A.; Small, I. Transactivation of a target gene using a suppressor tRNA in transgenic tobacco plants. Plant J. 1997, 11, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Heard, D.J.; Kiss, T.; Filipowicz, W. Both Arabidopsis TATA binding protein (TBP) isoforms are functionally identical in RNA polymerase II and III transcription in plant cells: Evidence for gene-specific changes in DNA binding specificity of TBP. EMBO J. 1993, 12, 3519–3528. [Google Scholar] [CrossRef] [PubMed]
- Waibel, F.; Filipowicz, W. U6 snRNA genes of Arabidopsis are transcribed by RNA polymerase III but contain the same two upstream promoter elements as RNA polymerase II-transcribed U-snRNA genes. Nucleic Acids Res. 1990, 18, 3451–3458. [Google Scholar] [CrossRef]
- Vankan, P.; Filipowicz, W. A U-snRNA gene-specific upstream element and a -30 ‘TATA box’ are required for transcription of the U2 snRNA gene of Arabidopsis thaliana. EMBO J. 1989, 8, 3875–3882. [Google Scholar] [CrossRef]
- Kiran, K.; Ansari, S.A.; Srivastava, R.; Lodhi, N.; Chaturvedi, C.P.; Sawant, S.V.; Tuli, R. The TATA-box sequence in the basal promoter contributes to determining light-dependent gene expression in plants. Plant Physiol. 2006, 142, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, K.; Imamoto, F. Selective and accurate initiation of transcription at the T-DNA promoter in a soluble chromatin extract from wheat germ. Mol. Gen. Genet. 1987, 209, 445–452. [Google Scholar] [CrossRef]
- Kim, J.S.; Yoon, S.J.; Park, Y.J.; Kim, S.Y.; Ryu, C.M. Crossing the kingdom border: Human diseases caused by plant pathogens. Environ. Microbiol. 2020, 22, 2485–2495. [Google Scholar] [CrossRef]
- Balmer, L.; Seth-Smith, H.M.B.; Egli, A.; Casanova, C.; Kronenberg, A.; Schrenzel, J.; Marschall, J.; Sommerstein, R. Agrobacterium species bacteraemia, Switzerland, 2008 to 2019: A molecular epidemiological study. Antimicrob. Resist. Infect. Control 2022, 11, 47. [Google Scholar] [CrossRef]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The Arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Sawant, S.; Singh, P.; Madanala, R.; Tuli, R. Designing of an artificial expression cassette for the high-level expression of transgenes in plants. Theor. Appl. Genet. 2001, 102, 635–644. [Google Scholar] [CrossRef]
- Iwataki, N.; Hoya, A.; Yamazaki, K. Restoration of TATA-dependent transcription in a heat-inactivated extract of tobacco nuclei by recombinant TATA-binding protein (TBP) from tobacco. Plant Mol. Biol. 1997, 34, 69–79. [Google Scholar] [CrossRef]
- Tikhonov, A.N.; Arsenin, V.Y. Solutions of Ill-Posed Problems; Halsted Press: Winston, WA, USA; New York, NY, USA, 1977; p. 258. [Google Scholar]
- Hahn, S.; Buratowski, S.; Sharp, P.; Guarente, L. Yeast TATA-binding protein TFIID binds to TATA elements with both consensus and nonconsensus DNA sequences. Proc. Natl. Acad. Sci. USA 1989, 86, 5718–5722. [Google Scholar] [CrossRef] [Green Version]
- Karas, H.; Knuppel, R.; Schulz, W.; Sklenar, H.; Wingender, E. Combining structural analysis of DNA with search routines for the detection of transcription regulatory elements. Comput. Applic. Biosci. 1996, 12, 441–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponomarenko, M.P.; Ponomarenko, J.V.; Frolov, A.S.; Podkolodny, N.L.; Savinkova, L.K.; Kolchanov, N.A.; Overton, G.C. Identification of sequence-dependent features correlating to activity of DNA sites interacting with proteins. Bioinformatics 1999, 15, 687–703. [Google Scholar] [CrossRef] [PubMed]
- Waardenberg, A.; Basset, S.; Bouveret, R.; Harvey, R. CompGO: An R package for comparing and visualizing Gene Ontology enrichment differences between DNA binding experiments. BMC Bioinform. 2015, 16, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, I.N. dbSNP in the detail and copy number complexities. Hum. Mutat. 2010, 31, 2–4. [Google Scholar] [CrossRef]
- Sharypova, E.B.; Drachkova, I.A.; Chadaeva, I.V.; Ponomarenko, M.P.; Savinkova, L.K. An experimental study of the effects of SNPs in the TATA boxes of the GRIN1, ASCL3 and NOS1 genes on interactions with the TATA-binding protein. Vavilovskii Zhurnal Genet. Selekt. 2022, 26, 227–233. [Google Scholar] [CrossRef]
- Godde, J.S.; Nakatani, Y.; Wolffe, A.P. The amino-terminal tails of the core histones and the translational position of the TATA box determine TBP/TFIIA association with nucleosomal DNA. Nucleic Acids Res. 1995, 23, 4557–4564. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, P.M.; Suslov, V.V.; Savinkova, L.K.; Ponomarenko, M.P.; Kolchanov, N.A. A precise equilibrium equation for four steps of binding between TBP and TATA-box allows for the prediction of phenotypical expression upon mutation. Biophysics 2010, 255, 358–369. [Google Scholar] [CrossRef]
- Sandukhadze, B.I.; Mamedov, R.Z.; Krakhmalyova, M.S.; Bugrova, V.V. Scientific breeding of winter bread wheat in the Non-Chernozem zone of Russia: The history, methods and results. Vavilovskii Zhurnal Genet. Selektsii 2021, 25, 367–373. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset # | Promoter | Transcribed Gene | Conditions | TBP | Pol | N | Ref. |
|---|---|---|---|---|---|---|---|
| 1 | TC7 | RNA transcript template of G-free sequence | In vitro: standard transcription reaction | Human | II | 24 | [45] |
| 2 | A chimeric in vitro system in which human TATA-binding protein (hsTBP) was replaced by purified TBP-1 of thale cress (atTBP) | Thale cress (Arabidopsis thaliana) | 24 | ||||
| 3 | OsPAL | OsPAL | In vitro: whole-cell extracts of rice cell suspension cultures | Rice (Oryza sativa) | II | 8 | [46,47] |
| 4 | Pv tRNA-Leu | Pv tRNA-Leu | In vitro: tobacco cell nuclear extract | Tobacco (Nicotiana plumbaginifolia) | III | 16 | [48] |
| 5 | Pv tRNA-Leu | gusA | Ex vivo: transient expression in tobacco protoplasts | III | 30 | [49] | |
| 6 | AtU6-26 snRNA | AtU6-26 snRNA | III | 7 | [50] | ||
| 7 | AtU2.2 snRNA | AtU2.2 snRNA | II | 7 | |||
| 8 | CaMV 35S | CaMV 35S | II | 7 | |||
| 9 | synthetic promoters based on AtU6snRNA | At U6-26 snRNA | III | 10 | [51] | ||
| 10 | synthetic promoters based on AtU2snRNA | At U2.2 snRNA | II | 5 | [52] | ||
| 11 | Pmec | gusA | In vivo: dark-grown tobacco leaves | II | 52 | [53] | |
| 12 | In vivo: light-grown tobacco leaves | II | 52 | ||||
| TOTAL | 10 promoters | 7 reporter genes | 6 experimental systems | 4 TBPs | 2 Pols | 242 | 9 Refs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasskazov, D.; Chadaeva, I.; Sharypova, E.; Zolotareva, K.; Khandaev, B.; Ponomarenko, P.; Podkolodnyy, N.; Tverdokhleb, N.; Vishnevsky, O.; Bogomolov, A.; et al. Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression. Int. J. Mol. Sci. 2022, 23, 8684. https://doi.org/10.3390/ijms23158684
Rasskazov D, Chadaeva I, Sharypova E, Zolotareva K, Khandaev B, Ponomarenko P, Podkolodnyy N, Tverdokhleb N, Vishnevsky O, Bogomolov A, et al. Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression. International Journal of Molecular Sciences. 2022; 23(15):8684. https://doi.org/10.3390/ijms23158684
Chicago/Turabian StyleRasskazov, Dmitry, Irina Chadaeva, Ekaterina Sharypova, Karina Zolotareva, Bato Khandaev, Petr Ponomarenko, Nikolay Podkolodnyy, Natalya Tverdokhleb, Oleg Vishnevsky, Anton Bogomolov, and et al. 2022. "Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression" International Journal of Molecular Sciences 23, no. 15: 8684. https://doi.org/10.3390/ijms23158684
APA StyleRasskazov, D., Chadaeva, I., Sharypova, E., Zolotareva, K., Khandaev, B., Ponomarenko, P., Podkolodnyy, N., Tverdokhleb, N., Vishnevsky, O., Bogomolov, A., Podkolodnaya, O., Savinkova, L., Zemlyanskaya, E., Golubyatnikov, V., Kolchanov, N., & Ponomarenko, M. (2022). Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression. International Journal of Molecular Sciences, 23(15), 8684. https://doi.org/10.3390/ijms23158684